基于Cyt b序列的贵州境内两江流域月鳢群体遗传结构
2022-09-30吴俣学张显波闵文武周其椿李世凯
吴俣学,张显波*,闵文武,周其椿,李世凯
(1.贵州省水产研究所,贵州 贵阳 550025;2.贵州省特种水产工程技术中心,贵州 贵阳 550025)
0 引言
【研究意义】贵州位于我国西南部,地处云贵高原东侧的大斜坡地带,处于长江和珠江两大流域上游交错地段,河流数量较多,具有充沛独特的水资源,加之丰富的地热、余热、冷泉水等资源以及特殊的喀斯特地形,孕育了种类繁多的水生生物,鱼类区系组成复杂,具有丰富的生物多样性。月鳢(Channaasiation)属鲈形目(Perciformes)鳢科(Channidae)鳢属(Channa),有七星鱼、山花鱼、点星鱼、星光鱼等俗称,属名贵淡水鱼类,具有极高的经济价值和药用价值,有保健鱼之称,为贵州原始土著鱼类之一。开展贵州月鳢自然种群的遗传背景研究,对贵州土著鱼类资源保护和开发具有重要的现实意义。【前人研究进展】研究人员利用ITS序列和Cyt b序列分析比较了月鳢、乌鳢和斑鳢之间的进化关系[1-2],也有相关研究从形态学方面分析鳢属鱼类系统发育关系[3],主要是针对同属不同种间的分析比较。Cytb基因是线粒体DNA上的编码区之一,进化速率适中、易扩增,比较适合阐明种内到种间的进化关系[4-6],广泛地应用于各类物种的系统发育研究[7-9]。【研究切入点】目前尚无关于贵州月鳢群体遗传学研究的相关报道,因此,以贵州境内长江流域和珠江流域的4个月鳢群体为研究对象,测定和分析其Cytb基因序列,对月鳢群体的遗传结构进行研究。【拟解决的关键问题】弄清贵州境内长江流域和珠江流域月鳢自然群体间的遗传关系,探究贵州两江流域月鳢自然种群的遗传背景,为贵州土著鱼类月鳢的种质资源保护和开发利用奠定理论基础。
1 材料与方法
1.1 材料
以贵州境内长江流域和珠江流域分布的月鳢群体为主要研究对象,共收集到月鳢130尾,均为自然群体,具体采样情况见表1。

表1 月鳢样本采集情况Table 1 Sample information of C. argus populations
1.2 基因组DNA的提取与检测
剪取月鳢样本的尾鳍下叶组织,无水乙醇浸泡,—20℃保存备用。参照改良的苯酚SDS法提取基因组DNA[10]。经1%琼脂糖凝胶电泳对提取的DNA进行完整性检测,浓度及纯度由NANODROP 2000分光光度仪测定,采用TE Buffer(pH=8.0)稀释至60 ng/μL,—20℃储存备用。
1.3 线粒体Cyt b序列片段的扩增和测序
利用Primer blast设计该片段的引物,得到上游引物Cytb-CDS-F1:atggccaatctacgaaaaacac,下游引物Cytb-CDS-R1:ttaacctccgacgtccggtt,引物由生工生物工程(上海)有限公司合成。PCR反应在PTC-200型PCR仪上进行,反应体系:上下游引物各2 μL,模板DNA 4 μL,MIX 20 μL,加无菌去离子水至40 μL;反应程序:95℃预变性3 min;94℃变性45 s,55℃复性45 s,72℃延伸1 min,并进行35个循环;72℃延伸10 min。扩增产物由生工生物工程(上海)有限公司测序。
1.4 数据分析
采用Clustal X软件将测序结果进行对位排序,并辅以人工校对。利用MEGA 5.0软件计算各种群间的遗传距离,运用基于遗传距离的邻接法(Neighbor-joining method,NJ)和基于信息位点的最大简约法(Maximum parsimonious criterion,MP)构建系统进化树,同时采用自举检验估计系统树中节点的置信度。用DNAsp5.0软件对各群体的单倍型多样性(Haplotype diversity,Hd)进行分析,估计不同种群的遗传多样性,并进行Tajima’s D检验统计。利用NCBI中 GenBank提供的生物信息资源共同进行数据分析。
2 结果与分析
2.1 4个月鳢群体的Cyt b序列特征
对贵州境内长江流域不同区域的2个群体和珠江流域不同区域的2个群体共130尾月鳢的Cytb基因的946 bp序列分析显示,A+T含量为53.7%,G+C含量为46.4%,其碱基组成差异不大。
长江流域乌江群体和清水江群体的碱基组成特征表现为保守位点940个,占核苷酸总数的99.37%,变异位点6个,其中,简约信息位点4个,单态突变位点2个;转换位点6个,无颠换位点。珠江流域猫营河群体和都柳江群体的碱基组成特征表现为保守位点939个,占核苷酸总数的99.26%,变异位点7个,其中,简约信息位点7个,无单态突变位点;转换位点6个,颠换位点1个。
2.2 4个月鳢群体的单倍型及核苷酸多样性
2.2.1 群体的单倍型多样性 从表2看出,长江流域和珠江流域4个月鳢群体130个样本共定义6种单倍型,其中,以单倍型Hap 4的个体分布最多,4个群体无共享单倍型,清水江群体独为1个单倍型。
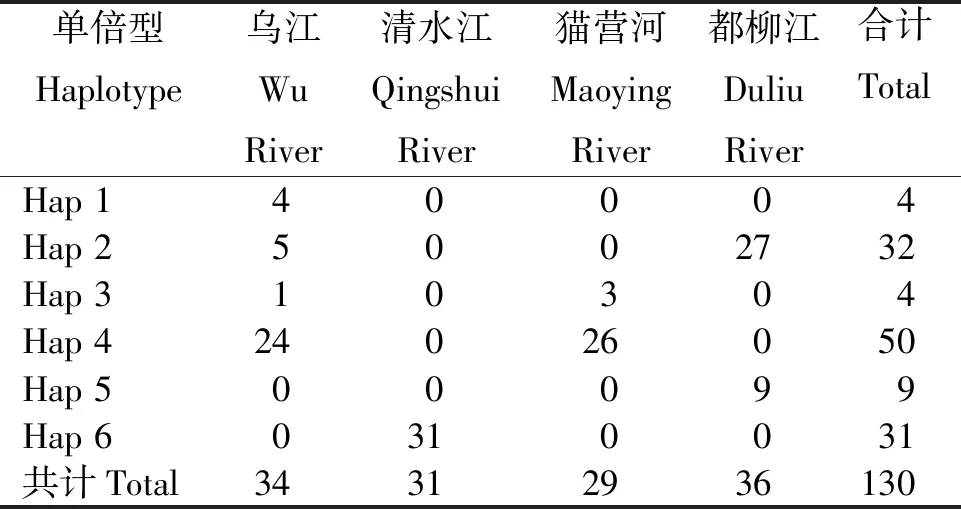
表2 4个月鳢群体Cyt b基因的单倍型分布Table 2 Distribution of the Cyt b haplotypes in C. argus population with four-month-age 尾
2.2.2 核苷酸多样性 从表3看出,长江流域乌江群体34个样本定义了4种单倍型,核苷酸多样性指数(π)为0.001 65,单倍型多样性指数(Hd)为0.480;长江流域清水江群体31个样本仅定义1种单倍型。珠江流域猫营河群体29个样本定义2种单倍型,π为0.000 61,Hd为0.192;珠江流域都柳江群体36个样本定义2种单倍型,π为0.002 04,Hd为0.386。

表3 4个月鳢群体的遗传多样性指数Table 3 Genetic diversity indexes of C. argus population with four-month-age
4个月鳢群体总体的π为0.001 98,Hd为0.734。长江流域月鳢群体出现5种单倍型,珠江流域月鳢群体出现4种单倍型。其中,乌江群体的遗传多样性最丰富,有4种单倍型;清水江群体遗传多样性水平最低,其π和Hd均为0,且只有1种单倍型。
2.3 4个月鳢群体的遗传距离与遗传分化指数
2.3.1 遗传距离 根据DNAsp的计算结果,4个月鳢群体间平均遗传距离(D)为0.002 29。从表4看出,猫营河-都柳江群体的D值最大,为0.003 08;乌江-猫营河群体的D值最小,为0.001 31;乌江-清水江群体、乌江-猫营河群体、清水江-都柳江D值低于平均水平,其他群体间的遗传距离值均大于平均遗传距离;但各群体间D值均小于0.01,表明群体间遗传差异较小,推测各群体间可能存在频繁的基因交流[11]。

表4 基于序列分析月鳢群体间的遗传分化Table 4 Genetic differentiation among C. argus populations based on sequence analysis
2.3.2 遗传分化 参照群体遗传分化标准(遗传分化指数Fst>0.25)[12],长江流域群体与珠江流域群体间基因存在一定的遗传分化。根据一般判断标准(Fst<1)[12],群体间未出现生殖隔离,各群体间仅乌江-猫营河群体的基因流(Nm)>1。根据线粒体DNA碱基突变率每百万年(Ma)1%~2.5%估算[13],种群间分化时间以猫营河-都柳江群体的分化时间最早(0.06 Ma~0.15 Ma),乌江-猫营河群体最晚(0.03 Ma~0.07 Ma),各群体间分化时间交叉在0.06 Ma~0.07 Ma。
2.4 4个月鳢群体的聚类
利用Genbank中斑鳢(Channamaculata,GenBank:MN057083.1)、乌鳢(Channaargus,GenBank:KU852447.1)作为同属外群同源序列,对所有月鳢单倍型进行NJ聚类和UPMGA聚类(图1)。从单倍型NJ树可看出,6个倍型分别聚为2支,其中,Hap3、Hap4、Hap5聚为1支,Hap1、Hap2、Hap6聚为1支,最后2支聚在一起。以布展值50为划分标准,长江流域与珠江流域月鳢种群内部单倍型遗传分化不明显,结合各群体单倍型分布情况及各群体间遗传距离的分析结果,长江流域及珠江流域月鳢群体之间种群分化不明显。月鳢群体与同属外种群明显分开,但与斑鳢遗传关系更近,与杨春英等[14]的研究结果一致。
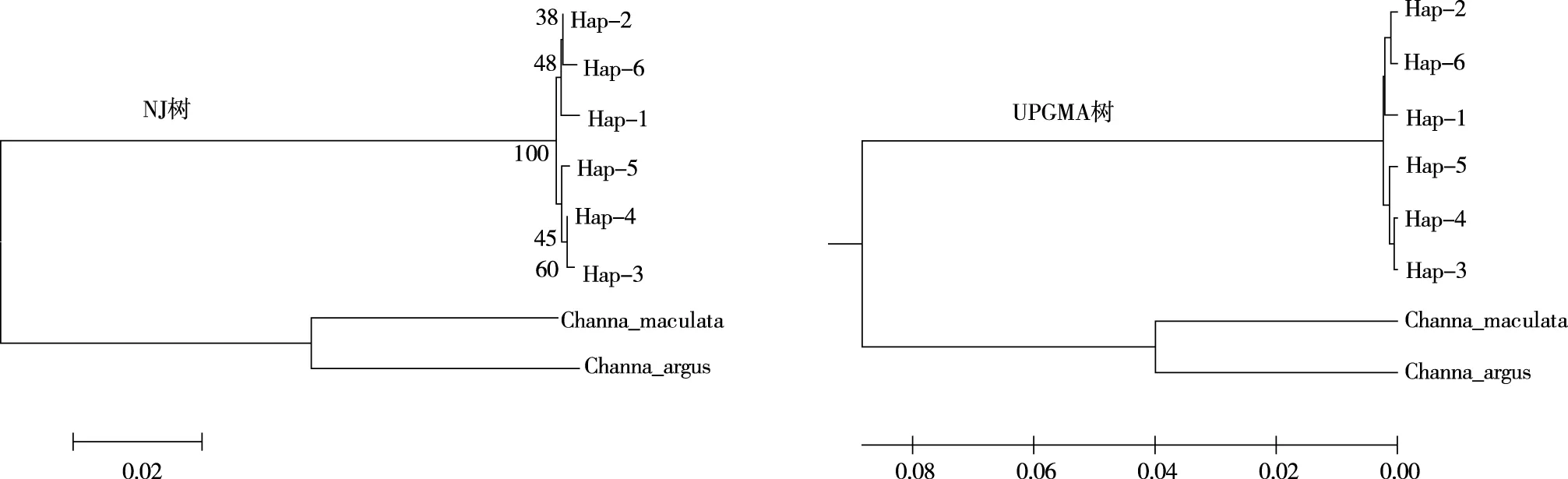
图1 4个月鳢群体基于cytb单倍型的NJ树及UPGMA树Fig.1 NJ tree and UPGMA tree of C. argus population with four-month-age based on Cyt b haplotype
月鳢单倍型UPGMA树聚类趋势和NJ树聚类趋同,均支持贵州月鳢种群的遗传关系划分为两大支系。
3 讨论
核苷酸多样性指数是判断群体遗传多样性水平的重要指标。研究结果显示,贵州境内长江流域(乌江、清水江)和珠江流域(都柳江、猫营河)4个月鳢群体遗传多样性水平呈乌江群体>猫营河群体>都柳江群体>清水江群体,总体呈长江流域>珠江流域。从单倍型数量上看,乌江群体出现4种单倍型,猫营河群体与都柳江群体均为2种单倍型,而清水江群体仅1种。根据GRANT等[15]对核苷酸多样性指数(π)<0.005则群体遗传多样性水平低的判断,贵州月鳢群体(π=0.001 98)整体遗传多样性处于较低水平。从近年来对相关水系鱼类遗传多样性的研究结果看,贵州境内月鳢遗传多样性低于小口白甲鱼[16]、清水江鲤[17]、都柳江鲇、光倒刺鲃[18]等经济鱼类。生物种群瓶颈效应、建群效应、栖息地破碎化以及遗传漂变等因素都有可能引起生物遗传多样性偏低,20世纪90年代有水产科技人员指出,月鳢面临资源衰减的困境[19]。根据本次对贵州月鳢采样的分布结果看,月鳢的保有数量相比20年前大大减少,人类活动很大程度上影响月鳢种群数量,这有可能是造成月鳢遗传多样性水平低的原因。清水江月鳢种群的主要遗传多样性参数数值均为0,且倍型单一,除人类活动外可能与采集到的月鳢为稻田养殖区的样本有关。黔东南地区月鳢主要在稻田养殖区,稻田养鱼相较河流有更大的局限性,造成月鳢群体的小范围地理隔离,从而影响群体遗传多样性水平。目前没有贵州月鳢遗传多样性的相关研究报道,随着长江流域开展“十年禁捕”工作,相信月鳢资源会得到一定恢复,今后可适时增加样品采集地以及使用其他分子标记如CoI、D-loop、ITS等再次对月鳢遗传多样性开展研究。
研究显示,贵州境内长江流域和珠江流域月鳢群体有一定的遗传分化,但群体间遗传距离不大,种群分化不明显。根据种群间分岐时间估算,贵州月鳢群体间分化时间最早在15万年前,晚于苗岭山脉成为长江流域和珠江流域诸水系分水岭的时间[20],说明地理隔离是造成两江流域月鳢群体遗传分化的重要因素。乌江月鳢群体和猫营河月鳢群体之间遗传分化程度不高,存在一定程度的基因交流,这可能是因为猫营河处于珠江流域和长江流域分水岭,两流域水系时有相互掠水现象,地理隔离不明显;也可能与贵州境内曾经开展过月鳢的人工养殖有关,推测造成两群体遗传分化的主要原因可能是生殖隔离。月鳢遗传分化程度高,基因交流不频繁,还可能与其生活习性相关。月鳢是广温、杂食性鱼类,对环境适应能力强,游泳距离短,不需要生殖洄游、索饵洄游及越冬洄游,甚至在稻田里面就能生存,造成一定范围的地理隔离和生殖隔离。
4 结论
采用PCR技术对贵州境内长江流域(乌江、清水江)和珠江流域(都柳江、猫营河)的4个月鳢群体(共130尾月鳢样本)进行遗传关系及遗传背景研究。结果表明,贵州月鳢Cytb基因946 bp序列的A+T含量为53.7%,高于G+C含量(46.3%),其碱基组成表现为C的含量最高、G的含量最低,呈较强的反G偏倚性。在130个月鳢样本中存在6种单倍型,总体核苷酸多样性指数为0.001 98,单倍型多样性指数为0.734。4个月鳢群体间平均遗传距离为0.002 29,以猫营河-都柳江群体的遗传距离最大(0.003 08),乌江-猫营河群体的最小(0.001 31)。总体看,贵州两江流域境内月鳢遗传多样性水平低,长江流域和珠江流域月鳢各群体种群分化不明显。可以利用各月鳢群体间遗传分化程度高的有利条件,加强各群体间亲本的交叉繁殖,尽量避免群体内部近亲繁殖,提升月鳢整体遗传多样性水平,达到保护贵州土著鱼类资源及开发利用的目的。
