干燥方法对水蜜桃微观结构变化的影响
2022-09-27袁越锦韩思明徐英英杨佳琪张国安施俊文
袁越锦 ,韩思明,徐英英,杨佳琪,张国安,施俊文
(1.陕西科技大学机电工程学院,陕西西安 710021;2.轻工业西安机械设计研究院有限公司,陕西西安 710086)
水蜜桃肉质鲜美,香味浓甜,营养丰富,且桃肉甘酸性温,具有生津活血、解烦止渴等功效,深受市场欢迎。但水蜜桃不易储存,通过干燥能够使它的水分减少,延长保存时间,增加经济效益。
水蜜桃细胞之间存在细胞间隙,水分和营养物质能够通过间隙输送,以维持细胞的生长。干燥过程伴随的高温高压等条件不仅会改变细胞和细胞间隙的原始形状,还会损失水蜜桃的营养物质。干燥方法不当会严重影响细胞间和细胞内的传热传质过程进而影响干燥效率和干燥品质。因此对于干燥过程的研究不应仅聚焦于复水、感官、色泽等方面,更应向细胞微观结构方向深入研究。
目前,将显微技术和图像处理技术结合在一起能够定量测定细胞微观结构变化的参数。描述果蔬微观结构变化的参数主要包括细胞截面积、截面周长、直径、圆度、孔隙率和分形维数等。Segura等通过分析对流干燥下苹果片的细胞面积、周长、直径及圆度等参数,认为干燥过程能显著改变细胞及细胞间隙的形状且苹果片的宏观变化与细胞微观变化行为相同,但是干燥过程中物料的宏观变化与微观变化的关系未定量描述。Wang 等利用光学显微镜和image-pro-plus 软件获得了黄桃经不同温度热风干燥后的细胞截面积频率分布曲线,认为高温会导致细胞形态难以维持。常剑等结合石蜡切片及显微观测技术分析了热风干燥过程中马铃薯、苹果、胡萝卜的微观结构并获得三种物料不同水分比下细胞结构图像和微观结构参数(面积、周长、当量直径等)的分布规律,还得到了微观结构参数和水分比的线性关系,然而由于干燥过程中微观结构参数的演化趋势较为复杂,因此该线性关系无法准确描述水分比和微观参数之间的关系。近年来,发现利用计算机模拟干燥过程在优化干燥工艺方面有极大的作用,因此许多研究者利用此方法来研究干燥过程中物料的微观结构变化。但是建立模拟所需的微观模型和数学模型必须获得物料微观结构参数,然而任意水分比下微观结构参数的测量步骤繁琐,因此可通过拟合得到水分比和细胞微观结构参数的函数关系,然后通过该方程更容易获得任意水分比下的微观结构参数。
为了描述不同干燥方法下水蜜桃微观结构的变化规律,结合石蜡切片制备、显微观测及图像处理等技术,获得了五种不同干燥方法对应的不同含水率下水蜜桃切片的显微图像和微观结构参数分布曲线且分析了微观结构参数的变化规律。通过多项式拟合建立了水分比与微观结构参数的拟合方程,将水分比与该方程结合起来能够预测干燥过程中微观结构参数。该研究结果为探究不同干燥方式下水蜜桃微观结构参数随水分比的变化规律和建立水蜜桃在干燥过程中的数学模型提供重要参考。
1 材料与方法
1.1 材料与仪器
实验用的水蜜桃 产于上海,品种为上海水蜜。其质量范围250±10 g,最大直径范围7±0.5 cm,每100 g 桃肉中约含水85 g,脂肪0.1 g,蛋白质0.8 g,碳水化合物7 g,粗纤维4.1 g,还包括维生素C 及胡萝卜素等其他营养物质总和约3 g,且要求果型圆润、硬度适中、品质新鲜;番红溶液(1 g 番红溶入100 mL 50%酒精)、蒸馏水、冰醋酸、甲醛(40%)、二甲苯、石蜡、中性树胶、固绿溶液(固绿0.1 g 溶入100 mL 95%酒精)、无水乙醇等 天津市科密欧化学试剂有限公司。
DZF-6090 型真空干燥箱 上海一恒科学仪器有限公司;SFY-60 型红外线水分测定仪 深圳市冠亚电子科技有限公司;JY104FC 型捷宇高拍仪 福建捷宇电脑科技有限公司;DHG 型热风干燥箱 上海一恒仪器有限公司;HH-3 型水浴锅 常州天瑞仪器有限公司;TF-LFD-1 型冷冻干燥箱 上海田枫实业有限公司;E100 型显微镜 尼康。
1.2 实验方法
1.2.1 水蜜桃干燥工艺 以水蜜桃为物料,选择HAD(60 ℃、风速1 m/s)、VD(60 ℃、0.08 MPa)、FD(预冻时间3 h、预冻温度-30 ℃)、HA-VD(热风/真空干燥条件同上、转换点含水率50%)及FHAD(冷冻/热风干燥条件同上、转换点含水率50%)五种不同的干燥方法进行实验。将同批次的新鲜水蜜桃处理干净后进行厚度为5 mm 的切片处理,然后在水蜜桃表皮下的相同位置(皮下2~7 mm 区域)进行取样(切为长10 mm、宽10 mm、厚5 mm的小块)。
对样品小块分别进行HAD、VD、FD、HA-VD和F-HAD 处理,实验时将样品小块分成两组先后进行以上五种工况下的干燥实验。第一组样本小块在同一个工况下进行5 次实验,然后在5 次实验过程中的同一干燥时间计算的5 个水分比求平均值,将该平均值作为样本小块在该时间的水分比。每次干燥过程中每隔5 min 对样品小块称重5 次,然后求得平均质量并记录;称重后将样本继续干燥且前后称重两次获得的平均质量差值小于0.001 g 则认为干燥结束(此时样本的质量为绝干质量),以此来计算每次实验中不同干燥时间的含水率和水分比。通过减小称重时间间隔,多次称重及增加同一工况下的实验次数能够更准确地获得干燥过程中样本小块的水分比到达0.8、0.6、0.4、0.2 时所需的干燥时间。第二组样本小块在干燥时根据第一组实验结果(即对应的水分比到达0.8、0.6、0.4、0.2 的干燥时间)再进行取样,由于干燥过程中物料收缩不均匀,因此不同干燥方式对应的每个水分比下取5 个显微观测样本。最后,将第二组取得的所有样品制作成显微观测所需的石蜡切片。
1.2.2 水蜜桃组织石蜡切片制备及显微图像处理 为了能够清晰地在显微镜下观察干燥后的水蜜桃组织的微观结构,采用石蜡切片法将第二组不同水分比下的样本制成石蜡切片标本(具体流程如图1 所示)。首先,将第二组干燥实验中取得的所有显微观测样品浸入甲醛(40%,5 mL)—冰醋酸(5 mL)—酒精(70%,90 mL)固定液至少24 h,接着将样本浸入不同质量分数的酒精(35%酒精中1.5 h—75%酒精中4 h—85%酒精中2 h—90%酒精中2 h—95%酒精中1 h—无水乙醇中1 h)中脱水;然后使用二甲苯对样品进行透明处理,接着将样品放入60 ℃融化石蜡(1 h,重复3 次)中浸蜡。其次,在热风温度为60 ℃的干燥箱中进行包埋,包埋之后将样品放入切片机切片,厚度为5 μm 左右;再将切片在60 ℃烘箱内烘干后依次浸入二甲苯(20 min,重复3 次)、无水乙醇(5 min,重复2 次)、75%酒精(5 min)和蒸馏水中脱蜡;将脱蜡后的切片先浸入番红染液染色后,再浸入不同浓度的酒精溶液(50%酒精中5 s—70%酒精中5 s—80%酒精中5 s—90%酒精中5 s)脱色;接下来使用固绿染液染色60 s,再利用醇苯(5 min)、二甲苯(5 min,重复2 次)对切片进行透明。最后,将透明后的切片放置于载玻片上,在30 ℃烘箱内烘干后盖上盖玻片,并用中性树胶封片。通过显微镜放大100 倍进行观察和拍照,然后利用Matlab 和Photoshop 软件捕捉细胞结构的原始图像。

图1 石蜡切片制备流程图Fig.1 Flow chart of paraffin section preparation
图2a 为冷冻干燥过程捕捉的细胞原始图像,在Matlab 中调用rgb2gray 函数先将显微镜拍摄的原始图片转换成灰度图像,如图2b;然后调用graythresh和im2bw 函数对细胞图像进行阈值分割,将灰度图像进行二值化表达,以此识别细胞的边缘轮廓,如图2c;最后利用photoshop 软件消除二值图像内部杂质,填充边缘及孔隙并调整图像亮度和对比度,最终图像如图2d。

图2 图像处理过程Fig.2 Image processing process
1.3 数据处理及测量指标
对得到的每张原图进行图2 中的图像处理过程得到对应的最终图像,再利用Matlab 软件中bwlabel、regionprops、bwperim 等函数测量和计算所有最终图像上每个细胞截面积和截面周长,并根据如下公式利用Matlab 编程计算水蜜桃细胞的圆度,壁面粗糙度及孔隙率。
细胞圆度S:

式中:A 为图像上的细胞截面积,m;L 为图像上的细胞截面周长,m。
细胞壁面粗糙度r:
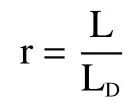
式中:L 为细胞图像上的实际周长,m;L表示与其细胞相似度一致的正多边形的周长,m。
孔隙率e:在截面上,多孔介质内部孔隙和多孔介质总面积的比值:

式中:A为多孔介质内部细胞面积,m;A为多孔介质内部总面积,m。
含水率:干燥过程中某一时刻称取5 次物料的质量并求得平均质量,然后利用该时刻的平均质量结合物料绝干质量计算该时间点样品的含水率w,其计算公式如下:

式中:m为t 时刻取样物料的平均质量,kg;m 为物料的绝干质量,kg。
水分比:干燥过程中样品某一时刻的含水率与样品初始含水率之比(即w/w)。
将所有计算结果以文本格式自动保存,利用Excel 软件对编程计算得到的不同干燥方式中每个水分比下5 个石蜡切片最终图像对应的微观结构参数求取平均值。将微观结构参数平均值的总分布范围划分为n 个小区间,统计每个细胞微观参数在不同的小区间内的细胞数量,再使用该小区间内的细胞数量除以每张图像上的细胞总数得到此小区间内的微观结构参数分布频率,计算公式如下:

式中:p为第m 个小区间内微观结构参数的分布频率;k为第m 个小区间内的细胞数量;K为每张图片上的细胞总数;1≤m≤n 且m、n 为整数。最后将所有小区间内的微观结构参数分布频率进行统计,并使用Origin 软件绘制各微观结构参数的频率分布曲线图。
2 结果与分析
2.1 不同干燥方式下水蜜桃细胞截面积的变化规律
从图3 中可以看出,新鲜水蜜桃的细胞截面积分布范围是600~78000 μm。HAD 过程中,面积为40000~78000 μm的细胞数量明显降低,面积为0~45000 μm的细胞数量略有增加,同时出现少部分面积大于80000 μm的细胞。这说明HAD 能够使水蜜桃细胞显著收缩,但是高温会导致部分细胞的细胞壁被破坏,从而导致几个细胞破裂后融合在一起形成“大细胞”。与HAD 相比,VD 能更有效保持物料的原始形状。水分比由1 降至0.6 的过程中细胞截面积变化幅度小,细胞截面积峰值处于区间8000~10000 μm内;继续干燥至水分比为0.2 时,细胞截面积在15000 μm下的细胞数目明显增加,小于20000 μm的细胞约占60%。这主要是因为水分的大量蒸发导致细胞失去支撑,再加上负压的共同作用造成了细胞截面积减小。FD 能最大程度地维持细胞的原始形状。从水分比为1 下降至0.8 时,细胞截面积平均值增加6%,这主要是因为在冷冻过程中细胞内水凝结成冰导致细胞略有膨胀;当水分比从0.6 下降至0.4 时,细胞内的部分冰晶由于真空条件直接升华,造成细胞内外存在压差,故细胞截面积有细微减小;水分比继续下降至0.2 时,细胞截面积平均值又有所增大,原因是细胞内剩余的冰晶持续升华为气体,气体向外扩散导致对细胞的“膨胀”作用加剧。

图3 不同干燥方法下细胞截面积频率分布曲线图Fig.3 Frequency distribution curve of cell area under different drying methods
HA-VD 条件下水分比从1 降至0.5 这一阶段为HAD 过程,面积为16000~78000 μm的细胞数量明显下降,面积在600~16000 μm的细胞数量明显增加,因此细胞收缩明显。转入VD 过程后,当水分比0.4 时细胞截面积最大值为6000 μm,水分比从0.4 降至0.2 时细胞截面积大于25000 μm的细胞数量增长8%,细胞截面积均值增大,可能是由于HAD过程破坏了细胞壁,使得VD 过程加剧了细胞破裂变形,再加上VD 过程细胞收缩率降低共同导致大细胞数目增多。F-HAD 条件下,水分比从1~0.6 这一阶段为FD 过程。水分比降至0.6 时,由于冰晶升华,因此20000 μm以下的细胞占比超过60%。转入HAD 过程后,主要是由于FD 过程中冰晶升华破坏了大量细胞结构,因此HAD 会导致大量面积较小的细胞破裂后在发生融合,故0~17000 μm的细胞数量明显减少,其融合导致面积在17000 μm以上的细胞数量显著增加。
随着干燥过程的进行,分布曲线的偏移量,曲线峰值增大或减小的程度各不相同,可能的原因是干燥过程中细胞内的水分转移不均匀导致细胞的收缩变形是不连续的。
2.2 不同干燥方式下水蜜桃细胞截面周长的变化规律
如图4,水蜜桃鲜样细胞截面周长主要分布范围为0~1300 μm。HAD 过程中,周长频率分布曲线峰值变化不大,但是水分比至0.2 时由于细胞破裂、融合和变形导致细胞截面周长大于750 μm 的细胞数量增加了6%。VD 过程中细胞发生形变但基本上不发生细胞破裂及细胞融合。当水分比至0.8 时细胞截面周长在1000 μm 以下的细胞数量减少了10%;水分比从0.8 降至0.2 的过程中,截面周长主要分布区间变为100~1780 μm,整个区间内几乎大部分曲线均右移但移动幅度不大,因此几乎所有细胞的截面周长略有增大。FD 过程中细胞截面周长呈增大趋势。水分比0.8 时,细胞截面周长均值增长10%左右;水分比0.6 时,细胞截面周长频率分布曲线左移,表明细胞失去较多水分皱缩。水分比降低至0.2 时与鲜样细胞相比细胞截面周长在500 μm 以上的细胞数量重新增加。
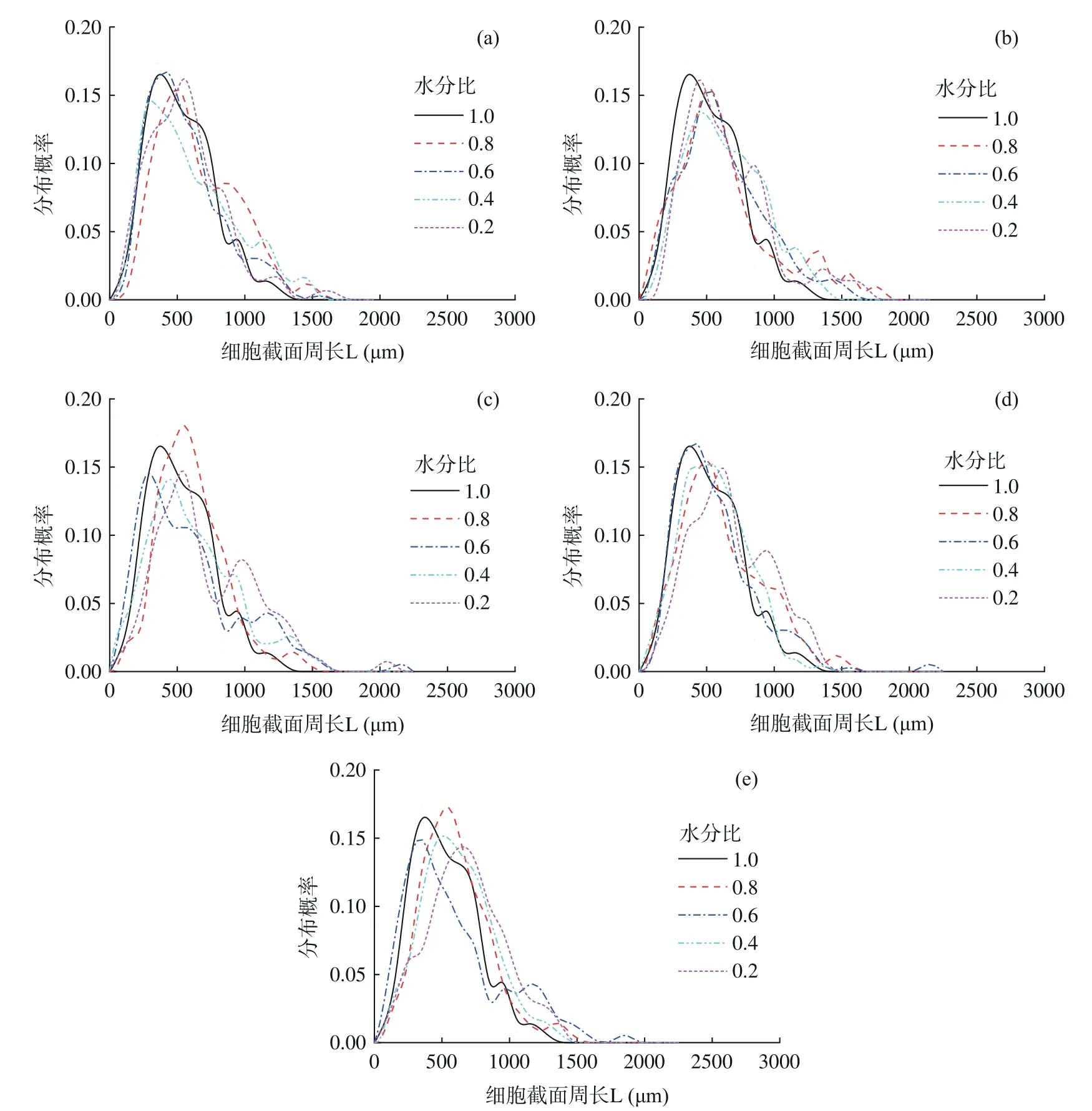
图4 不同干燥方法下细胞截面周长频率分布曲线图Fig.4 Frequency distribution curve of cell perimeter under different drying methods
HA-VD 过程中水分比降至0.2 时,有50%左右的细胞截面周长超过了670 μm,与最初未干燥的水蜜桃细胞相比提升了23%。F-HAD 过程中,当水分比为0.2 时与鲜样细胞相比,频率分布曲线右移明显,这主要是由于后半程的HAD 使细胞皱缩形变导致所有细胞截面周长变大。
2.3 不同干燥方式下水蜜桃细胞圆度的变化规律
从图5 可得水蜜桃鲜样细胞大部分圆度分布在0.4~0.9 之间,圆度大于0.85 的细胞占比约26%。HAD 过程中,从整体上看水蜜桃细胞圆度是减小的。水分比为0.2 时的细胞圆度均值相比于水分比0.4时有所增加,导致这一现象的原因是细胞破裂后又发生了融合。HAD 会导致细胞收缩,变形及细胞壁折叠,这都是细胞圆度发生变化的原因。VD 过程中,从整体上看细胞圆度是降低的。当水分比为0.8 时,细胞圆度分布范围主要为0.3~0.8,随着干燥过程的进行圆度频率分布曲线持续左移,这表明细胞圆度会随着水分比的持续降低而降低,水分比为0.2 时曲线峰值对应的圆度为0.35。FD 过程中,水分比从1 降为0.8 时细胞圆度变化幅度不大,75%以上的水蜜桃细胞圆度大于0.4,当水分比从0.8 降至0.6 时细胞圆度大幅度降低。水分比从0.6 降至0.2 这一过程中细胞圆度变化幅度较小,水分比0.2 为时细胞圆度主要分布在0.2~0.7 之间。

图5 不同干燥方法下细胞圆度频率分布曲线图Fig.5 Frequency distribution curve of cell roundness under different drying methods
HA-VD 过程中细胞圆度总体上呈减小趋势,但水当分比为0.2 时,圆度大于0.5 的细胞数量比VD 多10%左右,造成此现象的原因主要是前半程采用了不同的干燥方法。这表明相比于VD,热风干燥能更好的维持细胞圆度。F-HAD 过程中水分比1~0.5 之间为FD 过程,细胞圆度减小幅度大,其频率分布曲线左移并与FD 过程类似,当转入HAD 过程后,高温使得细胞变形、收缩和破裂,因此导致细胞圆度持续减小。
2.4 不同干燥方式下水蜜桃细胞壁面粗糙度的变化规律
由图6 可知,水蜜桃鲜样细胞壁面粗糙度主要分布在1~2 之间。干燥过程中随着水分比的降低,细胞壁将产生褶皱。HAD 过程中水分比从1 降至0.6 这一阶段,细胞壁快速皱缩导致细胞变形较大,当水分比降至0.6 时,曲线峰值对应的壁面粗糙度为1.5,此时壁面粗糙度主要分布在1.1~2.5 之间,这表明壁面粗糙度明显增加;在水分比从0.6 降至0.2 时这一过程中,由于部分细胞破裂后发生融合,因此曲线左移但移动幅度较小,这表明壁面粗糙度略微减小。VD 过程中水分比为0.8 时,70%的水蜜桃细胞壁面粗糙度在1.2 以上,曲线峰值对应的壁面粗糙度为1.3。与HAD 相比,VD 过程中细胞壁面粗糙度频率分布曲线向右移动更加缓慢。水分比为0.2 时,曲线峰值对应的壁面粗糙度为1.45,壁面粗糙度在1.26~1.75 之间的细胞占比超过60%。FD 相比其它干燥方式其平均壁面粗糙度最小且水分比从1 降至0.8 这一过程中壁面粗糙度变化幅度不大;水分比从0.8 降至0.6 这一过程中曲线右移明显,因此壁面粗糙度明显增大;水分比从0.6 至干燥结束,此阶段壁面粗糙度变化不明显;因此对于FD 过程,壁面粗糙度增大主要发生在前半程,后半程干燥过程几乎不变。

图6 不同干燥方式下细胞壁面粗糙度分布频率曲线图Fig.6 Frequency distribution curve of cell wall surface roughness under different drying methods
HA-VD 前半程壁面粗糙度的变化与单一HAD过程几乎相同;当水分比降至0.6 以下时,曲线峰值对应的壁面粗糙度都在1.3~1.5 之间;由图6(d)与单一HAD 相比,HA-VD 的壁面粗糙度均值介于HAD和VD 之间,这表明组合干燥进一步降低了细胞壁面粗糙度。F-HAD 整个过程中曲线一直右移,这表明壁面粗糙度持续增大,与单一HAD 过程相比其壁面粗糙度均值增大幅度明显降低。
2.5 不同干燥方式下水蜜桃细胞截面孔隙率的变化规律
由图7 可得,在HAD 过程中孔隙率先增大后减小,这主要是由于高温会导致细胞收缩并破坏细胞壁,随后发生破裂和融合;但这与香蕉、芒果等水果干燥时的孔隙率变化趋势正好相反,其趋势为先略微减小后增加。造成这种差异的原因主要是香蕉、芒果等水果干燥末期细胞的破裂融合程度没有水蜜桃剧烈。VD 和FD 过程中孔隙率一直增大。FD 过程中水分比为0.8~0.4 时孔隙率增长较快,这是由于冰晶升华的过程中细胞主要发生形变且细胞截面积减小,导致孔隙率的增大。FD 与VD 过程均主要发生细胞变形,但是细胞截面变形小且细胞破裂和融合的程度远低于HAD。
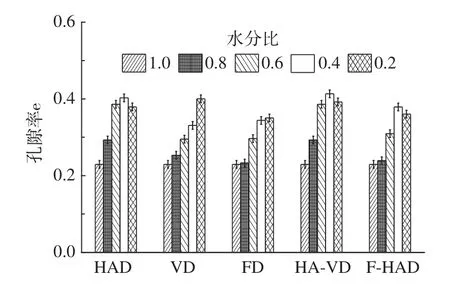
图7 细胞截面孔隙率变化直方图Fig.7 Cell cross-section porosity histogram
HA-VD 前半程孔隙率的变化与HAD 过程类似,转入VD 后,大部分细胞截面缩小的情况下,细胞截面积均值却增大(即小细胞数量减少,大细胞数量增加,导致最后阶段孔隙率减小),这一趋势与VD 略有不同。整个F-HAD 过程中孔隙率的变化与HAD类似,水分比为0.2 时孔隙率略有减小,这是由于FD后的稀疏孔隙分布在HAD 高温作用下细胞发生收缩,破裂及融合,故导致大细胞数量增加,因此孔隙率减小。
2.6 不同干燥方式下水分比与微观结构参数的关系
在干燥过程中,为了能够较易获取任意水分比下的微观结构参数,将水分比与微观结构参数进行拟合,只需测量干燥过程中任意时刻样本的水分比后代入拟合公式即可获得此时刻细胞的微观结构参数,拟合结果如图8。由于水蜜桃细胞微观结构参数与水分比呈非线性关系,故选取下列拟合公式:

式中:Y 表示细胞微观结构参数均值比;X 为水分比;a,b,c,d 为系数,见表1。
所有干燥过程完成后水蜜桃细胞截面积均值有增有减,截面周长均值、壁面粗糙度均值、孔隙率都增大,而细胞圆度均值减小。根据表1,Y 和X 的(除A/A(HAD)及L/L(HA-VD)外)均在0.9 以上。由图8 及表1 中的拟合系数综合分析,不同干燥方式对细胞大小和形状参数均值影响程度不同,其中包含HAD 干燥过程中对细胞截面积均值的影响较为剧烈;VD 和FD 对细胞截面周长均值影响程度更大;VD 对于细胞圆度均值影响最大而HAD 则最小;VD 对于细胞壁面粗糙度均值影响较小而HAD则最大;VD 对孔隙率的影响程度最大且干燥过程中孔隙率一直在增加。

图8 不同干燥方式下水蜜桃细胞微观结构参数均值比与水分比拟合曲线Fig.8 The correlation between the cell parameter ratios and the moisture ratio during different drying methods by polynomial fitting

表1 不同微观参数的拟合系数及决定系数Table 1 Fitting coefficients and determination coefficients of different microscopic parameters
由表中的可得,HAD 过程中圆度、壁面粗糙度和孔隙率,VD 过程中细胞截面周长、圆度及孔隙率,FD 过程中的细胞截面周长、壁面粗糙度和孔隙率,HA-VD 过程中的圆度、壁面粗糙度及孔隙率,以及F-HAD 过程中的细胞截面积,周长,孔隙率和壁面粗糙度与水分比的拟合效果均较好,以上参数均可作为建立不同干燥方式下宏微观关系式时可选择的水蜜桃细胞微观结构参数的代表。
3 结论
选择细胞截面积,截面周长,圆度,壁面粗糙度和孔隙率为微观结构参数,研究了不同干燥方法下水蜜桃细胞微观结构的变化规律,得出如下结论:
水蜜桃微观结构发生变化的阶段一般是在HAD前半程(水分比1~0.5)以及VD 和FD 的后半程(水分比0.5 以下)。选取水分比作为干燥过程中的宏观参数,在不同干燥方式下建立了水分比与细胞微观参数的拟合方程。结果表明细胞截面积均值的变化程度为:FD<HAD<VD<HA-VD<F-HAD,截面周长均值变大的幅度依次为:HAD<HA-VD<VD<F-HAD<FD,细胞圆度均值变小的程度依次为:HAD<FD<FHAD<HA-VD<VD,细胞壁面粗糙度均值增大幅度依次为:F-HAD<VD<FD<HA-VD<HAD,孔隙率增大的程度依次为:FD<F-HAD<HAD<HA-VD<VD。因此与单一干燥方法相比,相对应的组合干燥能够显著抑制细胞截面周长,圆度,壁面粗糙度及孔隙率的变化。
不同干燥方法对细胞大小和形状参数均值的影响程度明显不同,其中,HAD 干燥过程中对细胞截面积均值的影响较为剧烈;VD、FD 及F-HAD 对细胞截面周长均值影响程度更大但差异不大;HAD 对于细胞圆度均值影响最小而VD 则最大;VD 对于细胞壁面粗糙度均值影响较小而HAD 则最大;VD 对孔隙率均值的影响最大,但是不同干燥方法对干燥结束后孔隙率的影响差异不大。
