荒漠草原柠条灌丛堆不同发育阶段土壤细菌群落结构特征
2022-09-14王子寅刘秉儒李子豪赵晓玉
王子寅 刘秉儒 李子豪 赵晓玉
(1.北方民族大学生物科学与工程学院,银川 750021;2.国家民委黄河流域农牧交错区生态保护重点实验室,银川 750021;3.宁夏特殊生境微生物资源开发与利用重点实验室,银川 750021)
柠条锦鸡儿(Caragana korshinskii Kom.)隶属豆科(Leguminosae),多年生落叶灌木植物,主要分布于我国西北、华北等水土保持和防风固沙地区[1]。柠条具有抗逆性强、根系发达、能产生一定经济价值、改良土壤和植被生态效益等诸多优良特性[2]。柠条在生长发育过程中逐渐形成灌丛堆,它通过自身的生理代谢和与土壤微生物间的相互作用而改变土壤的物理特性,从而促进土壤的形成,并使流动沙丘向半固定、固定沙丘转变[3]。因此实施退耕还林还草工程项目以来,在我国西北干旱及半干旱荒漠草原地区大面积种植柠条,用于沙丘固定和土壤质地的改良,显著改善了局部地区的生态环境,对荒漠化治理具有重要影响[3]。
土壤微生物是影响土壤形成与演化过程的重要因素[4],能够直观反映其生态功能的变化[5]。土壤微生物作为生态系统中不可或缺的组成部分,在土壤的养分转换、有机质的分解和维持生态系统的稳定过程中起着重要作用,是生物地球化学循环过程的关键驱动者[6]。卞莹莹等[7]研究发现相较于平茬和天然柠条林,人工种植柠条地下微生物数量、活性更佳,且土壤结构和肥力较好。以土壤细菌为主的土壤微生物数量较多、分布较为广泛,参与了土壤中多数养分的循环转化,推动着生态系统的物质循环和能量转换[8]。因此,通过对柠条灌丛堆土壤细菌群落结构的分析,可以更好地理解柠条灌丛在不同发育阶段的养分状况,为促进柠条灌丛堆的可持续发展提供重要依据。
以往对柠条的研究主要集中在其土壤水热条件[9-10]、生物力学性质[11-12]以及土壤微生物[13-14]等方面,但对于生长期、成熟期和衰退期3种不同发育阶段的柠条灌丛堆土壤细菌群落结构及多样性的研究少有报道。鉴于此,本文采用高通量测序技术,通过研究柠条灌丛堆不同发育阶段土壤细菌的群落结构和多样性,明确土壤细菌群落对不同发育阶段土壤养分变化的影响,以期为防风固沙区域柠条林的可持续发展提供一定的理论依据。
1 材料与方法
1.1 材料
试验所选取柠条灌丛堆位于宁夏盐池县南部荒漠草原(37°22'-37°24'N,106°57'-106°59' E),北邻毛乌素沙地,位于鄂尔多斯台地和黄土高原的过渡地带,海拔约为1 500-1 600 m,属典型的中温带大陆性季风气候,干旱少雨,年降水量约为280-300 mm,集中于7-9月,约占全年降水量的70%[3]。年气温约为7-9℃,有效积温约为3 000℃,无霜期在150 d左右[15]。植物种类以柠条锦鸡儿(Caragana korshinskii Kom)、 赖 草(Leymus secalinus(Georgi)Tzvel)、 白 草(Pennisetum centrasiaticum Tzvel)、 糙隐子草(Cleistogenes squarrosa(Trin.)Keng)、长芒草(Stipa bungeana Trin)等干旱、中干旱灌木草本为主,灰钙土、风沙土是研究区主要土壤类型[3]。
1.2 方法
1.2.1 试验设计 于2020年8月下旬植物生长旺盛时期,随机选取15个外形规则、生长态势均匀一致但大小有梯度差异的柠条灌丛堆(选取最大、最小灌丛堆基本可以代表当地最大、最小灌丛堆)[3]。参考熊小刚等[16]的方法将选取的15个灌丛堆按照高度及水平差异将其划分为生长期(growing)、成熟期(mature)、衰退期(recession)3类灌丛堆,每组分别选取5个灌丛。采样时分别在沙堆顶部(U)、中部(M)及底部(D)3个部位采集土样,按东南西北4个方向的0-10 cm土层,将土样等量混匀后作为一个样品,然后将每个样品分成两份,一份立即放入冰盒带回实验室用于微生物特征的测定,另一份于室内风干后测定土壤理化性质[3]。
1.2.2 土壤理化性质的测定 土壤样品采集完毕经室内自然风干后,测定土壤电导率(electrical conductance,EC)、pH、有机碳(soil organic carbon,SOC)、总磷(total nitrogen,TP)、速效磷(available phosphorous,AP)、总氮(total nitrogen,TN)等指标[3]。采用水土比为2.5∶1的电位计法测定土壤pH值;采用便携式电导率仪测定土壤EC;采用TOC测定仪测定土壤SOC;采用硫酸-高氯酸消煮-钼锑抗比色法测定土壤TP;采用碳酸氢钠浸提-钼锑抗比色法测定土壤AP;采用硫酸钾-硫酸铜-硒粉消煮,定氮仪自动分析法测定土壤TN[17]。
1.2.3 样本DNA提取、16S rRNA基因的PCR扩增及高通量测序 样品送至美吉生物医药科技(上海)有限公司,进行土壤样品的DNA提取、16S rRNA基因的PCR扩增以及高通量测序。选择细菌V4区域的16S rRNA进行PCR扩增,通用引物为515FmodF(5'-GTGYCAGCMGCCGCGGTAA-3') 和806RmodR(5'-GGACTACNVGGGTWTCTAAT-3')[18]。
1.2.4 统计分析 使用美吉生物云平台(www.majorbio.com)进行稀释曲线、韦恩图、细菌群落相对丰度图及功能预测等图形绘制。使用IBM SPSS Statistics 24.0软件对土壤理化性质等数据进行单因素方差分析(One-way ANOVA)。使用Origin 2021软件对土壤细菌多样性及丰富度进行 Paired Comparison Plot。
2 结果
2.1 不同发育阶段柠条灌丛堆土壤理化性质比较
随着柠条灌丛堆发育阶段的变化,如表1所示,土壤养分在不同发育阶段之间存在显著性差异(P<0.05),土壤pH在7.6左右,呈弱碱性。AP和EC在成熟期表现为最大值;SOC、TP和TN没有显著性差异;SOC、TP、AP、TN以及EC在柠条灌丛堆不同发育阶段均表现为先升高后降低。
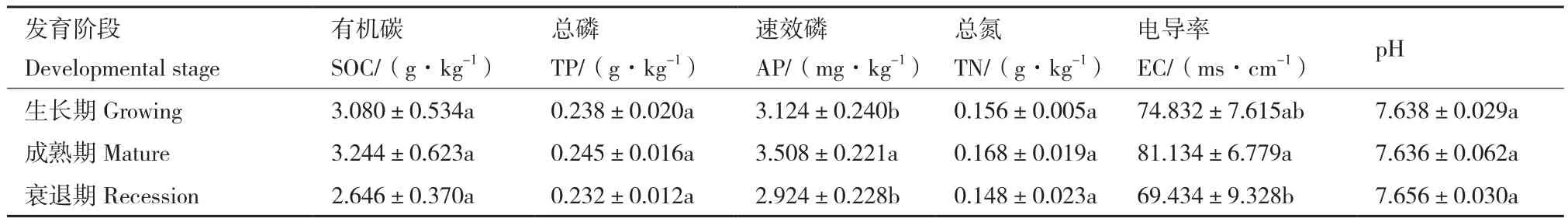
表1 不同发育阶段柠条灌丛堆土壤理化性质Table 1 Soil physical and chemical properties of Caragana korshinskii Kom.Nebkhas at different developmental stages
2.2 高通量测序结果及Alpha多样性评估
高通量测序结果如表2所示,不同发育阶段样品序列数在56 050-69 731之间,平均测序序列数为62 670,覆盖度均在95%以上。不同发育阶段柠条灌丛堆土壤细菌类群之间无显著性差异,但3种发育阶段之间柠条灌丛堆多样性(Shannon指数)和丰富度(ACE和Chao 1指数)数值均表现为成熟期(mature)大于生长期(growing)和衰退期(recession)(P<0.05)。
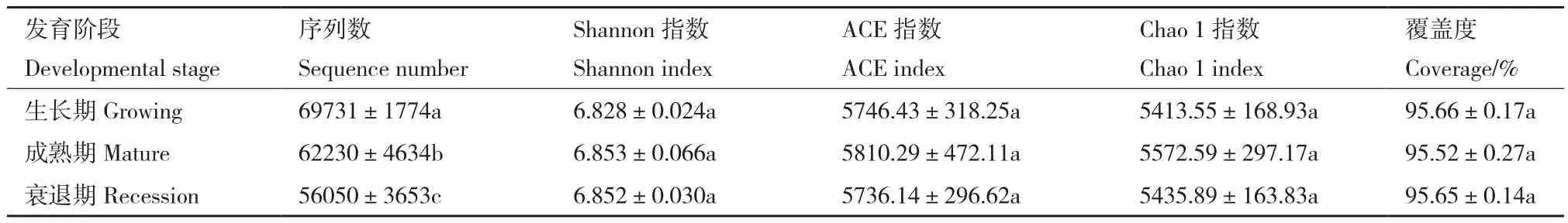
表2 不同发育阶段柠条灌丛堆土壤细菌群落多样性Table 2 Soil community diversity of C.korshinskii Kom.nebkhas at different developmental stages
2.3 土壤群落结构组成分析
Veen图可以较为直观的表现不同发育时期柠条灌丛堆土壤细菌群落OTU的组成差异及其共有物种的情况。经对有效序列进行质控、拼接和优化,通过 RDP 数据库的注释以及在97%的相似水平下的OTU进行生物信息统计分析,由图1可知,共获得15 383个细菌OTU,在生长期、成熟期、衰退期土壤中分别获得12 513、12 362和12 201个细菌OTU。生长期、成熟期、衰退期3个阶段共有OTU数为9 369,只占OTU总数的60.90%;3个阶段特有的OTU数量分别为1 160、1 037和862,分别占OTU总数的7.54%、6.74%和5.60%;生长期和成熟期共有的OTU数量为985(6.40%),生长期和衰退期共有的OTU数量为999(6.49%),成熟期和衰退期共有的OTU数量为971(6.31%)。
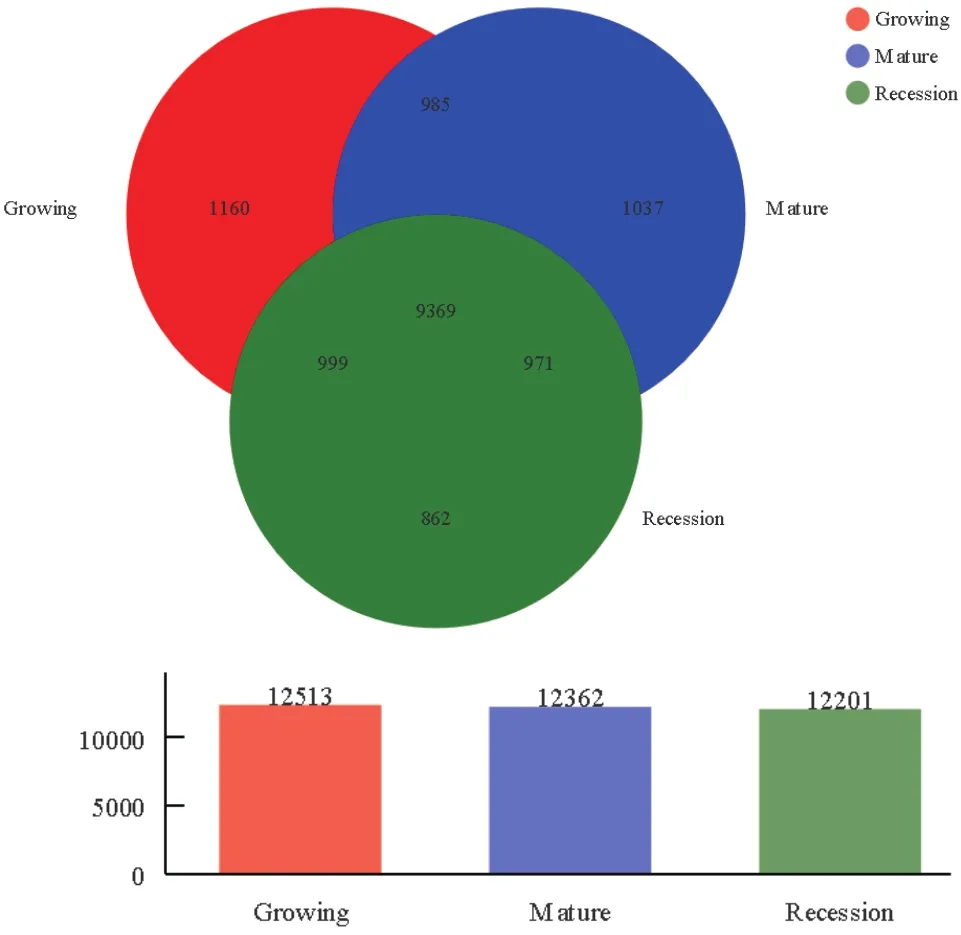
图1 不同发育阶段柠条灌丛堆土壤细菌群落韦恩图Fig.1 Venn diagram of soil bacterial community at different developmental stages of C.korshinskii Kom.nebkhas
高通量测序结果显示,按照最小样本序列数抽平后,柠条灌丛堆不同发育阶段土壤细菌隶属50门146纲364目590科1 111属。在不同分类水平上统计柠条灌丛堆不同发育阶段的物种丰度,通过群落柱形图可以直观的研究群落的结构组成,并且能够反映各发育阶段中优势物种的分布比例。如图2所示,不同发育阶段土壤细菌群落中优势菌群有放线菌门(Actinobacteriota)、变形菌门(Proteobacteria)、酸 杆 菌 门(Acidobacteriota)、 绿 弯 菌 门(Chloroflexi)、泉古菌门(Crenarchaeota)、浮霉菌门(Planctomycetota)、芽单胞菌门(Gemmatimonadota)、黏球菌门(Myxococcota)、拟杆菌门(Bacteroidota)等9种,约占微生物总数的90.00%-93.82%。其中占比大于5%的有放线菌门占比27.50%-90.62%,变形菌门占比14.35%-20.75%,酸杆菌门占比10.93%-14.30%,绿弯菌门占比9.34%-11.20%,泉古菌门占比6.46%-8.98%,芽单胞菌门占比4.92%-6.34%,浮霉菌门占比4.25%-5.66%。
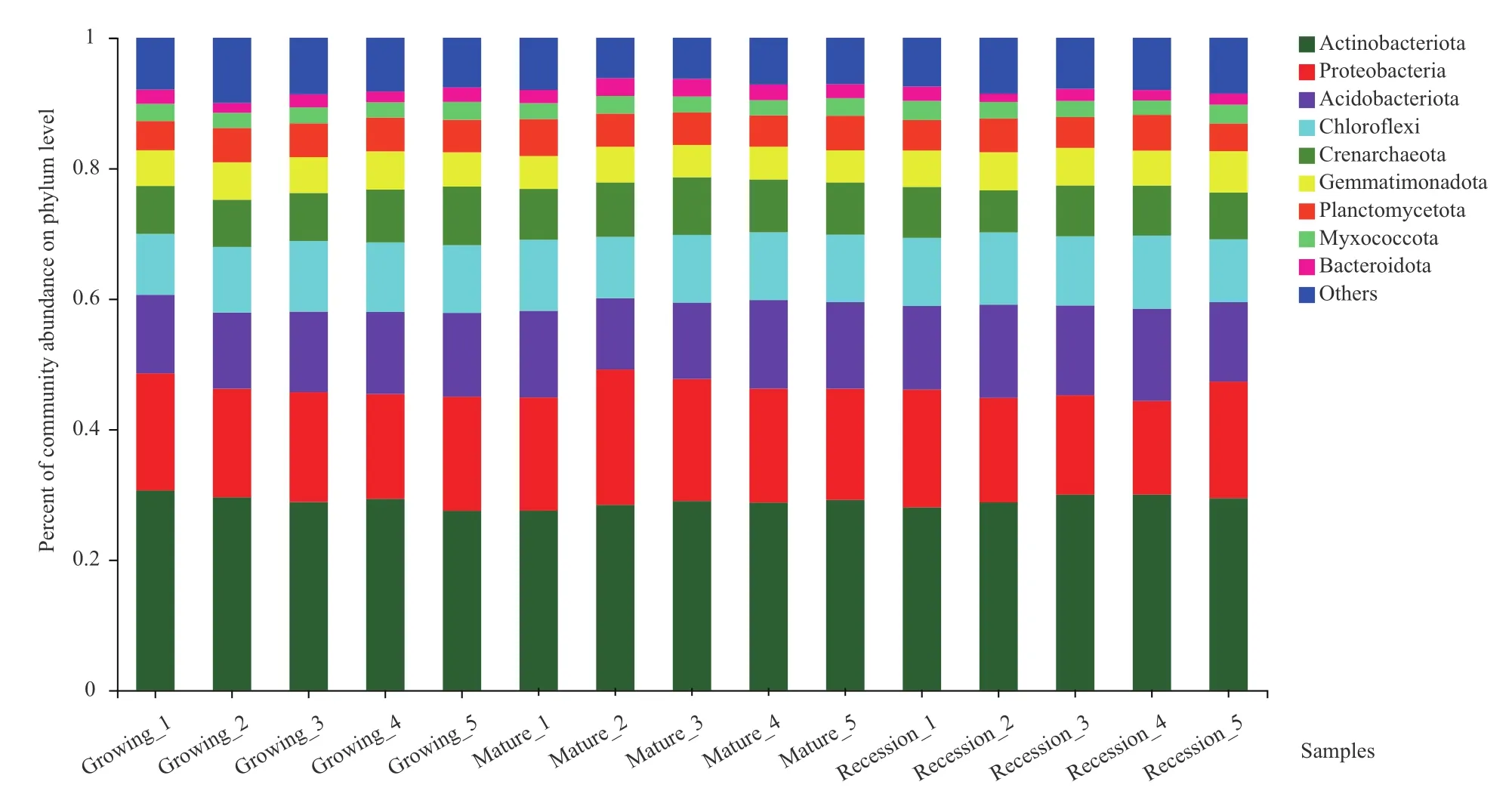
图2 门分类水平上细菌群落相对丰度图Fig.2 Relative abundance of bacterial community at the phylum level
为了寻找不同发育阶段柠条灌丛堆土壤细菌群落结构中具有显著差异的微生物门,基于各发育阶段群落丰度数据,选取门分类水平下Top50优势物种,对不同发育阶段土壤细菌相对丰度差异通过单因素方差(one-way ANOVA)分析,来评估物种丰度差异的显著性水平,从而得到具有显著性差异的物种,并且通过丰度降序,列出了生长期、成熟期和衰退期3个发育阶段中存在显著差异性的前11种细菌(图3)。结果发现,前11种细菌分别是Gemmatimonadota(芽单胞菌门)、Bacteroidota(拟杆菌门)、Firmicutes(厚壁菌门)、unclassified_k__norank_d__Bacteria、Methylomirabilota、Patescibacteria(髌骨细菌 门 )、Entotheonellaeota、GAL15、NB1-j、WPS-2以及Dadabacteria。其中,Patescibacteria和Entotheonellaeota的丰度差异性达到非常显著水平,NB1-j丰度差异性达到了极显著水平,Gemmatimonadota、Bacteroidota、Firmicutes、unclassified_k__norank_d__Bacteria、GAL15、WPS-2和 Dadabacteria的丰度差异性均达到显著水平。
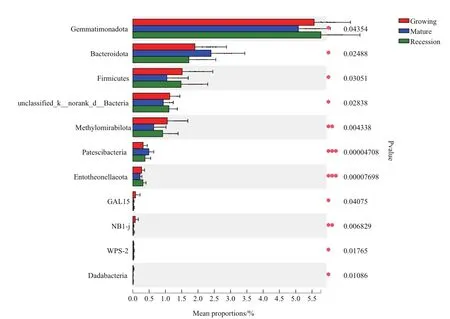
图3 门水平物种相对丰度差异分析Fig.3 Analysis on the relative abundance differences of dominant fungi at the phylum level
2.4 不同发育阶段柠条灌丛堆土壤细菌优势类群与土壤理化性质的相关性
通过Person相关性热图分析土壤理化性质对不同发育阶段柠条灌丛堆土壤细菌门水平群落组成的影响(图4)。结果显示,在门分类水平上,AP、SOC、TP、EC、pH、TN均与土壤细菌总丰度前20的优势类群存在相关关系,AP和SOC是影响不同发育阶段柠条灌丛堆土壤细菌群落结构的主要影响因子。其中,AP对 Bacteroidota、Bdellovibrionota、Proteobacteria、Patescibacteria、Thermoplasmatota、unclassified_k__norank_d__Bacteria、Firmicutes、Methylomirabilota、Gemmatimonadota、Actinobacteriota和Nitrospirota门呈非常显著相关关系,对Crenarchaeota门呈极显著相关关系,对Entotheonellaeota门呈显著相关关系;SOC对Bacteroidota、Proteobacteria、Thermoplasmatota、unclassified_k__norank_d__Bacteria、Firmicutes、Methylomirabilota门呈非常显著相关关系,对Crenarchaeota、Bdellovibrionota、Patescibacteria、Gemmatimonadota、Armatimonadota门呈极显著相关关系,对Nitrospirota、Acidobacteriota、Chloroflexi门呈显著相关关系,另外EC、pH、TP、TN也显著影响多个门菌群的丰度。
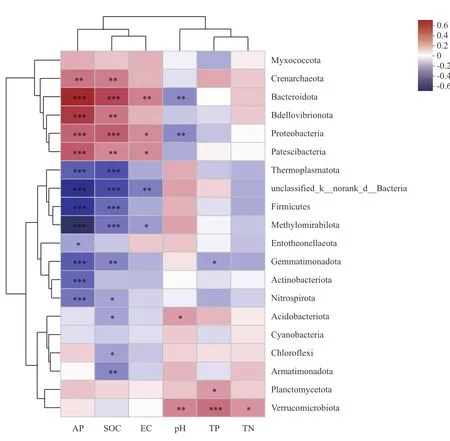
图4 门水平的土壤细菌优势类群与环境因子相关性热图Fig.4 Heat map of the correlation between dominant ant groups of soil bacterial and environmental factors based on phylum level
2.5 土壤细菌网络分析与PICRUSt功能预测
物种相关性网络图主要反映某一环境条件下各分类水平的物种相关性。选用Pearson模型,设定参数为相对系数绝对值≥0.5、P<0.05,对生长期、成熟期和衰退期柠条灌丛堆土壤细菌中相对丰度排名前30的属进行单因素网络分析,如图5所示。本研究中不同发育阶段柠条灌丛堆土壤细菌的单因素网络呈连通状态,网络直径是4,节点间平均最短路径长度为1.773,共210个边,对应29个节点。前30个物种OTU,分别属于泉古菌门(Crenarchaeota)、放线菌门(Actinobacteriota)、酸杆菌门(Acidobacteriota)、变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、芽单胞菌门(Gemmatimonadota),网络中心性数值最高的节点为变形菌门(Proteobacteria)的Sphingomonadales属,与其他门类20个属进行互作,大多数为负相关。
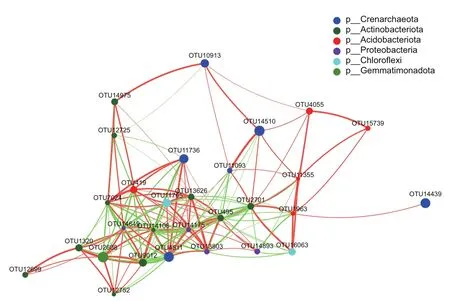
图5 土壤细菌网络图Fig.5 Network of soil bacteria
为了解不同发育阶段柠条灌丛堆土壤细菌群落功能,采用PICRUSt软件,将测序所得到的16S rRNA数据,基于 KEGG(kyoto encyclopedia of genes and genomes)数据库进行对比预测,在一级功能层共获得 6 类通路:细胞过程(cellular processes)、环境信息处理(environmental information processing)、遗传信息处理(genetic information processing)、人类疾病(human diseases)、代谢(metabolism)、有机系统(organismal systems)和目前分类学尚未分类的通路。其中代谢、遗传信息处理和环境信息处理是一级功能层中最主要的组成部分,丰度值分别为68 277 089.01-69 450 987.46、6 067 770.89-6 107 170.17和4 298 763.26-4 400 881.69,占比分别为78.76%-78.86%、6.93%-7.00%和4.95%-4.99%。二级功能层共有46类通路,其中相对丰度大于1%的有18类,如表3所示。在生长期、成熟期、衰退期3个阶段,土壤细菌群落结构中一级和二级通路的数量和多样性差异不显著。
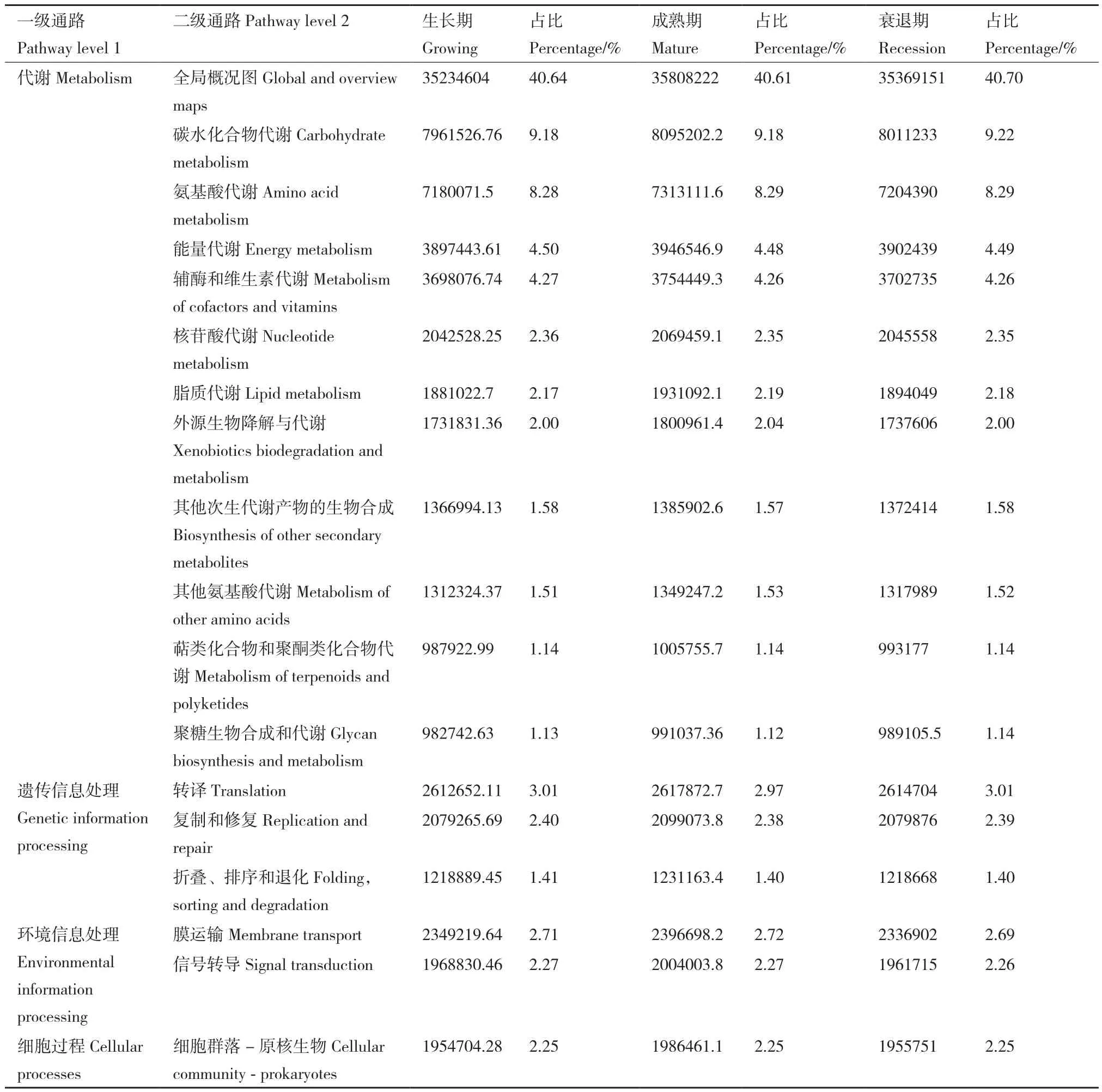
表3 土壤细菌群落中相对丰度>1%的二级功能丰度表Table 3 Secondary functional abundance table with relative abundance > 1% in the soil bacterial community
3 讨论
在荒漠草原干旱与半干旱地区,植物灌丛堆可以通过堆积枯落物等手段富集自己所需养分。杨阳等[19]研究发现柠条灌丛堆对土壤养分有明显的富集作用。一方面,柠条灌丛堆的发育能够明显增加荒漠地区沙面粗糙度,在发育过程中通过逐渐抵挡风沙侵蚀、截取大气降尘等方法,增加土壤黏粒的含量[20],从而改变土壤容重和保水性,进而增加土壤养分的储存量[21];另一方面,通过根际分泌物的释放、根系等衰老组织的分解以及枯落物的堆积,可以为微生物提供有利的生存环境,从而加快土壤养分的积累[22]。本研究中不同发育阶段的柠条灌丛堆土壤AP和EC存在显著性差异,表现为成熟期>生长期>衰退期,即随着柠条灌丛堆的发育,其对土壤养分的富集呈现先增加后降低,在成熟期表现为最大的现象,其他土壤养分如SOC、TP、TN、pH虽然不存在显著性差异,但同样在数值上表现为成熟期>生长期>衰退期,与陈鸿洋等[23]、裴世芳等[24]研究结果一致。说明在柠条发育过程中由于成熟期生长旺盛,更新周期短,并且其冠层结构茂密更有利于堆积枯落物,最终使得成熟期柠条灌丛堆的土壤养分显著高于生长期和衰退期。
土壤中细菌菌群特征受到土壤理化性质、气候、植被类型、生理[25]等多种因素的影响。本研究从柠条灌丛堆总体土壤细菌类群特征上分析,检测出土壤细菌隶属50门146纲364目590科1 111属,不同发育阶段检测出的细菌总体门种类及数量相近,没有显著差异;各发育阶段共有的OTU占比达60%以上,特有的OTU数量低于10%;以及本研究兼顾数据的可信度,未深入分析属种层次。原因可能由于样地之间没有显著的地理隔离,加之气候干旱等因素的影响。研究表明,不同类型的土壤细菌群落的优势种类基本是相似的,包括放线菌门(Actinobacteriota)、变形菌门(Proteobacteria)、酸杆菌门(Acidobacteriota)、绿弯菌门(Chloroflexi)、浮 霉 菌 门(Planctomycetota)、 芽 单 胞 菌 门(Gemmatimonadota)、拟杆菌门(Bacteroidota)等[26]。这些菌门在冻土、森林、湿地、荒漠等地形中也常被作为优势菌门被检出[27-32]。本研究中测定出荒漠草原区柠条灌丛堆土壤中丰度最高的4个的细菌门类分别是放线菌门、变形菌门、酸杆菌门、绿弯菌门,4个门类共同丰度占总检出数量的60%以上,优势菌门同样为以上菌门,且优势程度明显。
土壤环境和土壤细菌群落的组成有着密不可分的联系,沈聪等[14]研究表明,柠条根际土壤pH值、AN和TK与细菌群落关系密切,而在本研究中,门水平的土壤细菌优势类群与环境因子相关性热图表明,AP、SOC、TP、EC、pH、TN均与土壤细菌总丰度前20的优势类群存在相关关系,其中AP是影响细菌群落组成结构的最主要因子,说明土壤中P含量的增加能够影响微生物的群落结构,这与纳小凡等[33]的研究结果一致。PICRUSt功能预测通过与KEGG数据库进行对比,将微生物的变化与生物体功能紧密联系[34]。一级功能层中代谢、遗传信息处理和环境信息处理三类丰度较高,能使柠条灌丛堆土壤细菌代谢更加旺盛、长势更好,从而提高其群落结构的多样性。二级功能层中主要有全局概况图、碳水化合物代谢、氨基酸代谢、能量代谢、辅酶和纤维素代谢、转译等46类通路,证明其功能的丰富性。但目前PICRUSt功能预测仍然存在较多局限,随着科技的进步,后续可通过宏基因组学等方法进一步详细研究柠条灌丛堆不同发育阶段土壤细菌的群落功能。
4 结论
荒漠草原区柠条灌丛堆不同发育阶段土壤细菌隶属于50门146纲364目590科1 111属,主要优势门为放线菌门、变形菌门、酸杆菌门和绿弯菌门。不同发育阶段成熟期的土壤养分含量最高,其细菌的多样性和丰富度也处于最高水平。AP和SOC是影响土壤细菌群落结构的主要影响因子,TP、EC、pH、TN也有一定影响。不同发育阶段土壤细菌功能丰富,在代谢、遗传信息处理和环境信息处理方面表现活跃。