MicroRNA-219在脂毒性环境下对肝细胞脂质代谢、细胞增殖及凋亡的影响
2022-09-13徐梓馨廖红雨李慧一徐铭益
徐梓馨 廖红雨 李慧一 徐铭益
肝细胞脂质代谢、细胞增殖及凋亡等情况的变化对非酒精性脂肪性肝病(non-alcoholic fatty liver disease, NAFLD)进展至关重要[1-4]。miRNA是一种在不同物种中高度保守的非编码小RNA[5]。miRNA-122、miRNA-223、miRNA-34、miRNA-29、miRNA-194等在肝脏疾病中,发挥重要作用[6-9]。文献报道,miRNA-219(miR-219)在四氯化碳诱导的肝纤维化小鼠模型中可抑制肝星状细胞胶原沉积,进而抑制肝纤维化的进展;在肝癌小鼠模型中,miR-219可调控肝癌细胞的增殖、凋亡和迁移,发挥保护作用[10,11]。本研究通过构建脂肪肝小鼠和脂肪肝细胞模型,了解miR-219在脂毒性环境下对肝细胞脂质代谢、细胞活性与凋亡的影响。
材料与方法
一、材料与试剂
实验小鼠购于上海斯莱克实验动物中心,小鼠低脂饲料及高脂饲料均购于南通特洛菲实验饲料有限公司,胎牛血清、高糖DMEM培养基、0.25%胰蛋白酶购自美国Gibco公司,1%青霉素/链霉素双抗溶液、棕榈酸(palmitic acid,PA)、Lipo3000脂质体转染试剂、TRIzol试剂购自美国Invitrogen公司,EDU试剂盒购自广州锐博生物科技有限公司。PCR引物序列由生工生物工程(上海)股份有限公司合成,反转录试剂盒、SYBR Green PCR试剂盒,HE染色、油红染色试剂盒均购于上海翊圣生物科技有限公司。
二、方法
(一)动物模型构建 共有10只8周龄雄性SPF级C57BL/6J小鼠用于脂肪肝小鼠模型的构建。小鼠体质量为25~30 g,由上海斯莱克实验动物中心提供,饲养于上海交通大学附属第一人民医院实验动物中心。10只小鼠随机分为低脂组(low fatty diet,LFD)与高脂组(high fatty diet,HFD),每组5只,LFD组小鼠喂养普通饲料(即低脂饲料),HFD组小鼠喂养高脂饲料。喂养16周后处死小鼠,取部分肝组织经脱水、石蜡包埋或OCT凝胶包埋用于病理学检测。取适当组织进行q-PCR检测,余组织冻存于液氮中。
(二)肝组织病理检查 建模小鼠处死后,取适当大小的新鲜组织块置于4%甲醛溶液中固定48 h,脱水处理、石蜡包埋。蜡块以4 μm 切片,HE染色,光学显微镜下观察其病理改变。新鲜肝组织经过4%甲醛中固定48 h,依次经过20%、10%蔗糖溶液梯度各24 h脱水后,用OCT溶胶包埋。以10 μm厚度进行冰冻组织切片,用于油红染色,光学显微镜下检测其病理程度并拍照。
(三)原代肝细胞(primary hepatocytes,PHC)提取、培养及处理 使用两步胶原酶消化法分离8周龄雄性C57BL/6J 野生型小鼠的PHC[12]。分离得到的PHC用含10% FBS、50 mg/mL青霉素和链霉素的DMEM培养基,于37 ℃,体积分数为0.05的CO2培养箱中培养。细胞分为阴性对照组(mi-NC)和miR-219过表达组(mi-miR-219),按照说明,用miR-219模拟物(miR-219 mimic)或阴性对照(mi-NC)转染PHC,转染浓度为50 nmol/mL,转染完成24 h后,再以200 μmol PA处理PHC细胞24 h。
(四)原代肝细胞油红染色 在PHCs细胞转染完成培养24 h后更换含有200 μmol PA的完全培养基继续培养24 h,按照试剂盒说明进行细胞油红染色,光学显微镜下观察细胞脂质沉积情况。
(五)生化检测 全自动生化分析仪检测对照组和PA处理组细胞上清液中AST、ALT水平。操作按照试剂盒说明书进行。
(六)EDU检测 原代肝细胞转染完成且PA处理之后,按照EDU试剂盒说明书进行EDU孵育、细胞固定化以及ApolloR染色,在荧光显微镜下观察。
(七)QRT-PCR法检测miRNA、mRNA相对表达量 采用TRIzol法提取总RNA并纯化,进一步测定RNA样品的浓度和纯度,判断样品纯度是否符合实验要求。按照试剂说明书反转录合成cDNA后行RT-PCR。本研究主要检测PA处理的肝细胞内miR-219的表达,以及脂质代谢相关因子硬脂酰辅酶A去饱和酶1(stearoyl-CoA desaturase 1,SCD1)和脂肪酸合成酶(fatty acid synthase, FAS)[13],细胞增殖关键分子增殖细胞核抗原(proliferating cell nuclear antigen ,PCNA)和细胞周期蛋白D1(CyclinD1)[14,15],以及凋亡相关分子半胱氨酸蛋白酶-3(Caspase-3)和B细胞淋巴瘤-2(Bcl-2)的mRNA表达。对应的引物序列见表1。实验过程中以U6、β-actin分别作为miRNA和mRNA的内参对照,进行标准化转换得到各样本的拷贝数(Ct值),样品目的基因的相对表达率(relative expression,RQ)=2-△△CT。每个样品的内参基因和靶基因各设3个重复试验。
结 果
一、非酒精性脂肪性肝小鼠模型的建立
为了构建非酒精性脂肪性肝病小鼠模型,将小鼠随机分为HFD组和LFD组,分别喂养高脂饲料和普通饲料。HE染色显示LFD组小鼠肝组织小叶结构完整,中央静脉细胞排列有序,未见明显脂肪变性及炎性细胞浸润(图1A),HFD组可见明显的肝小叶结构紊乱,出现明显的脂肪泡及气球样变,伴随大量炎性细胞浸润(图1B)。油红染色显示,LFD组小鼠肝细胞形态、结构正常,细胞核居中,仅存在少量脂质沉积(图1C),HFD组小鼠肝组织可见大量明显的脂质沉积于汇管区周围(图1D)。HE染色和油红染色结果表明非酒精性脂肪性肝病小鼠模型构建成功。

表1 qRT-PCR相关引物序列
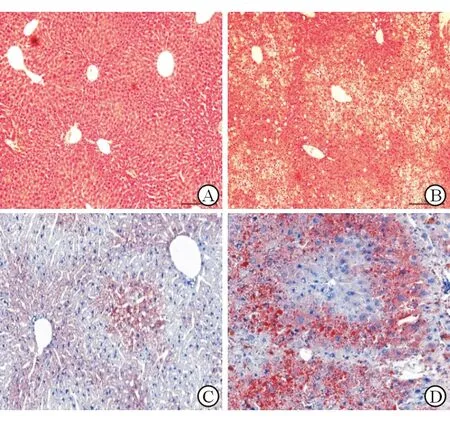
A、B:HE染色;C、D:油红染色(×100)
二、非酒精性脂肪性PHC细胞模型建立
油红染色显示,对照组PHC中红色脂滴较少,经过PA处理的PHC内红色脂肪颗粒明显增加(图2)。PA处理组PHC细胞上清液AST、ALT水平分别为12.743 U/L和7.410 U/L,高于对照组7.427 U/L和3.577 U/L,差异均有统计学意义(t=8.650、7.828,P=0.001、0.0014)。这些结果表明非酒精性脂肪肝细胞造模成功。
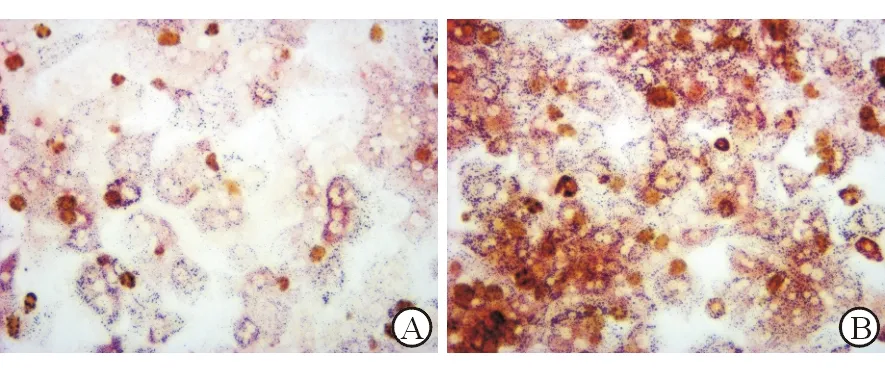
A:对照组; B: PA处理组
三、脂肪肝小鼠肝组织和脂毒性PHC中miR-219表达
在非酒精性脂肪性肝病小鼠模型中,HFD组小鼠肝组织miR-219表达水平为0.455±0.028,LFD组为1.000±0.125(P<0.05)。从野生型小鼠分离提取PHC,PA处理以模拟高脂环境,发现PA处理组PHC中miR-219的表达水平为0.676±0.064,对照组为1.000±0.190(P<0.05); miR-219的表达在高脂小鼠和PA刺激的PHC中均明显下降,说明在非酒精性脂肪性肝病时,肝细胞中miR-219的表达受到明显抑制,肝细胞功能也可能会受到影响。
四、在高脂环境下,过表达miR-219缓解PHC内脂质沉积
为进一步探究miR-219对脂毒性环境下肝细胞的脂质代谢的影响,在PA处理的PHC中过表达miR-219。结果显示miR-219在PHC内升高1000倍,过表达miR-219的PHC细胞模型成功构建。油红染色发现mi-miR219组PHC内红色脂肪颗粒明显变少(图3)。进一步检测脂质代谢相关因子SCD1和FAS的表达,发现过表达mi-R219的PHC细胞内,SCD1(mi-miR219+PA:0.539±0.048,miR-NC+PA:1.000±0.033,P<0.01)和FAS(mi-miR219+PA:0.722±0.036,miR-NC+PA:1.000±0.051,P<0.05)的mRNA表达均明显下降。这些结果表明在高脂环境下,miR-219可缓解肝细胞脂质代谢紊乱。
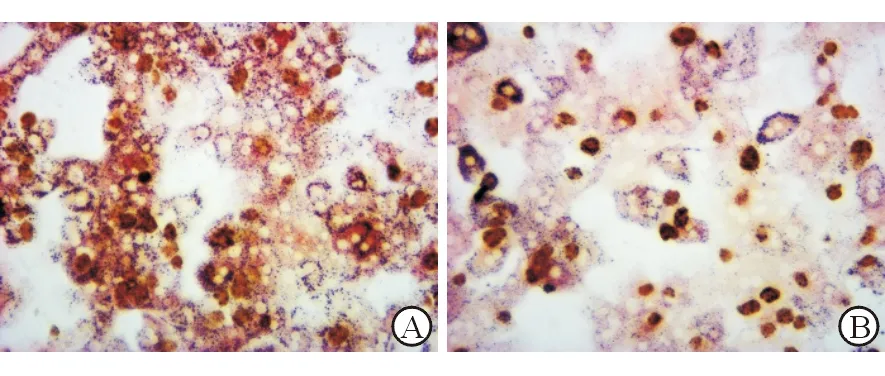
A: miR-NC+PA;B: mi-miR219+PA
五、在高脂环境下,过表达miR-219促进PHC细胞增殖并抑制细胞凋亡
为进一步探索过表达miR-219对高脂环境下肝细胞的增殖及细胞凋亡的影响,进一步检测了增殖相关因子(PCNA、CyclinD1)的表达。与miR-NC+PA组相比,mi-miR219+PA组的PHC中PCNA(mi-miR219+PA:1.652±0.185,miR-NC+PA:1.000±0.203,P<0.05)和CyclinD1(mi-miR219+PA:1.791±0.154, miR-NC+PA:1.000±0.135,P<0.05)的mRNA表达均出现明显上升。EDU检测提示过表达miR-219后,细胞增殖明显增强(图4),说明在高脂环境下,miR-219可提高PHC的活性,促进PHC增殖。
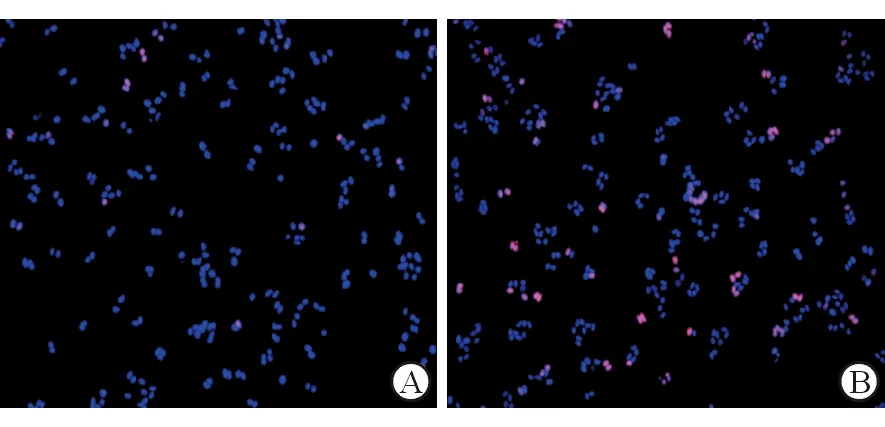
A: miR-NC+PA;B: mi-miR219+PA
此外,与对照组相比,过表达miR-219后,PHC细胞中凋亡相关基因Caspase-3表达明显下降(mi-miR219+PA:0.574±0.054,miR-NC+PA:1.000±0.35,P<0.05),Bcl-2基因表达明显上升(mi-miR291:1.535±0.109,miR-NC:1.000±0.208,P<0.05);说明在高脂环境下,过表达miR-219可明显促进肝细胞增殖,抑制肝细胞凋亡,提示mi-R219可能缓解脂毒性环境下肝细胞受到的损伤。
讨 论
Zhang等[16]发现在脂肪肝小鼠模型中,miR-103可通过靶向FAS和SCD1抑制肝脏新生脂肪生成,进而缓解NAFLD。miR-219可以通过抑制胶原沉积、促进细胞增殖、抑制凋亡等作用,在肝纤维化、肝癌中发挥作用[9-10]。
在本研究中,HE和油红染色证实成功构建了NAFLD小鼠模型。在HFD组小鼠肝组织中,发现miR-219表达水平明显下降。一些脂质(如PA)具有脂毒性,可导致肝细胞脂质代谢紊乱、活性受损、细胞凋亡增多乃至死亡[17.18]。从野生型小鼠分离提取原代肝细胞,用PA处理以模拟脂毒性环境,发现在PA处理的PHC中miR-219的表达水平也出现明显下降,这说明miR-219在高脂环境下表达受到显著抑制。
在PHC中过表达miR-219后,进一步检测肝细胞脂质代谢、细胞增殖及凋亡等情况。SCD1和FAS能够促进肝细胞脂质的合成,促进脂质在细胞内沉积,是反映脂质代谢水平的关键因子。相比对照组,过表达miR-219组PHC脂质代谢因子SCD1、FAS的表达水平出现下降,脂质沉积也明显减轻。提示miR-219可能对改善肝细胞的脂质代谢紊乱,缓解脂质沉积起关键作用。PCNA和CyclinD1是细胞DNA合成过程中的重要因子,其表达水平可反映细胞活性或细胞增殖的情况[14,15]。Q-PCR检测过表达miR-219后PHC中这两个基因的表达水平,发现PCNA和Cyclin D1的表达均出现明显上升,EDU检测也提示细胞增殖明显增强。此外,凋亡相关因子Caspase-3与细胞凋亡成正相关,促进细胞凋亡[19],而Bcl-2负调控细胞凋亡[20],过表达miR-219可明显抑制Casepase-3的表达,而促进Bcl-2的表达。这些表明过表达miR-219可缓解高脂对肝细胞活性的影响,促进增殖并抑制凋亡。
综上所述,在高脂环境下,miR-219不仅可显著缓解肝细胞脂质代谢紊乱,减轻脂质沉积,也能够减轻脂毒性对细胞活性的影响,促进细胞增殖抑制凋亡,发挥保护肝细胞的作用。但是miR-219在脂肪肝进程中起保护作用的具体机制及其下游靶基因仍需要进一步的探索。
