宁夏河东沙区丝棉木树干液流昼夜变化及其受气象因子的影响1)
2022-09-08彭苓韩磊韩永贵柳利利
彭苓 韩磊 韩永贵 柳利利
(宁夏大学,银川,750021)
树干液流可反映树木体内的水分传输状况,是土壤-植物-大气连续体(SPAC)水流路径中的一个关键链接,是研究树木耗水特性和水分传输机制的重要指标[1],树干液流有99.8%以上消耗于蒸腾[2]。大多数研究者针对树干液流的研究集中在白天,而针对夜间液流的研究较少[3-4],夜间液流不仅可以补充因白天蒸腾造成的水分亏缺[5],还能为树木器官间呼吸提供氧的传递机制,维持树木体内水分平衡[6]。太阳辐射直接影响植物的光合作用,太阳辐射强度增加,光合作用及蒸腾速率增加[7],茎干液流速率增加,气温是通过影响植物的生理活动和植物叶面温度来影响蒸腾,进而影响树干液流,植物通过加强蒸腾作用来降低叶面温度,以免高温灼伤,当空气干燥时,相对湿度较低,使得空气水汽压差变大,水分更容易从植物体散失[8]。温淑红等[9]通过研究发现,林木夜间液流主要受风速影响,而孔喆[10]的研究发现夜间液流受饱和水汽压差影响较大,风速则抑制夜间液流速率。随着沙地植被的恢复和重建,防护林蒸腾耗水的研究越来越被广泛关注。
宁夏河东沙区位于毛乌素沙地西南缘,鄂尔多斯台地东北部,西临黄河,南靠黄土高原[11],在空间上是我国北方农牧交错区最具有过渡性地域特征的沙地[12],该区域气候干旱,生态脆弱,营建防护林是最主要的防风固沙措施,但受传统造林理论的局限,随着林木的逐年生长,林水矛盾日益突出,林地退化问题严重[13]。丝棉木(EuonymusbungeanusMaxim.)抗性强[14],尤其抗盐碱、干旱能力较强,在干旱半干旱缺水地区和盐碱地造林中广泛应用[15]。而对丝绵木的研究,多以药用价值、育苗栽培、光合生理特性为主[16-18],对丝绵木的蒸腾耗水特性鲜有研究。为了解宁夏河东沙区典型防护林树种蒸腾耗水特征,本文以宁夏河东沙区主要造林树种丝绵木作为研究对象,采用热扩散茎流计实时监测林木树干液流,分析树干液流昼夜变化特征及其影响因素,为揭示林木蒸腾耗水调控机理提供依据,同时为干旱沙区防护林结构优化提供依据。
1 研究区概况
研究区位于宁夏回族自治区灵武市白芨滩国家级自然保护区(105.59°~106.37°E,37.60°~38.01°N),地处毛乌素沙地西南缘,属典型的大陆性干旱气候,气候特征表现为太阳辐射强、日照充足,降水少、蒸发强烈。其多年平均气温为8.8 ℃,年日照时间为3 080.2 h,年均降水量为209.7 mm,年均蒸发量为1 800 mm。降雨主要集中在6—9月,约占年均降水量的72.4%,降水时空分布不均匀。气候干旱,植被类型单一,以荒漠草原植被为主[19],防护林树种主要有丝绵木、刺槐(Robiniapseudoacacia)、新疆杨(Populusbolleana)等。
2 材料与方法
2.1 树干液流测定
选取4株生长发育良好、无病虫害的丝绵木作为研究对象。采用热扩散式茎流计(SF-L,Ecomatic Ltd.,German)对生长状况基本一致的丝绵木进行持续监测,茎流计外接太阳能板和12 V胶体电池保证24 h不间断供电。传感器分别由2根直径1.5 mm、长35 mm探针组成。为避免阳光直射引起热效应[20],探针安装在树干北部1.3 m高处,并外裹铝箔。数据采集器(CR-1000,Campbell Scientific Co.Ltd.,USA)与气象数据同步,采用Vantage Pro 2自动气象站(Davis Inc.,USA)每隔15 min自动记录1次数据,使用生长锥确定样木边材断面积。
根据Granier[20]建立的热扩散探针上下探针的温度差与树干液流密切相关的原理,利用两者关系的经验公式将温度差转换为树干液流:
(1)
式中:Js为单株样木液流密度(g·cm-2·s-1);ΔT是加热探针和数据传感探针之间的瞬时温差(℃);ΔTmax为1 d中最大的温度差值(℃)。
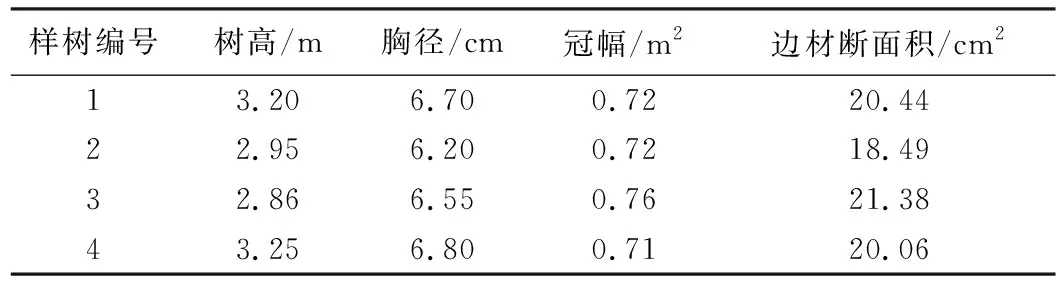
表1 样树基本信息
2.2 环境因子
选择林内空旷的平地,在样地周边安装自动气象监测站,采用美国Vantage Pro 2自动气象站,每15 min记录1次太阳辐射(Rs,W·m-2)、气温(Ta,℃)、相对湿度(Rh,%)、风速(W,m·s-1)、降水量(P,mm)等气象数据,并在试验地布设雨量筒测定区域降雨量,检验自动气象站降雨数据。饱和水汽压(Vpd,kPa)采用大气温度和相对湿度实测值进行计算[21],计算公式如下:
(2)
采用时域反射仪(TDR)每隔7 d测定丝绵木林地土壤含水量,通过烘干法进行检验,每隔10 cm进行1次取样,直到100 cm深度。平均土壤含水量为各层含水量的土层厚度加权平均值。
(3)
式中:Swc为土壤含水率(%),m0为烘盒质量(g),m2为烘前土样和烘盒质量(g),m1为烘后土样和烘盒质量(g)。
2.3 数据分析
以太阳辐射≤5 W·m-2对连续观测的丝绵木树干液流数据进行昼夜区分[22], 从20:00—06:00为夜间,06:00—20:00为日间,分析丝绵木树干液流的昼夜变化规律,通过气象数据筛选典型晴天进行相关性研究。采用SPSS21.0对液流密度进行方差分析,并对树干液流密度与各环境因子进行Pearson相关性分析;利用Orgin2018进行绘图。
3 结果与分析
3.1 环境因子变化特征
丝绵木树干液流密度与生长季(5—9月)各气象因子变化情况如图1所示。研究期内,气象因子季节变化较大,降水集中在6和8月,晴天时间多,温湿度季节变化明显,液流密度季节性差异大。5、7和9月土壤含水率保持较为稳定,6、8月降水量较多,土壤含水率较高,各月平均土壤含水率由大到小依次为6月、8月、7月、5月、9月(表2)。
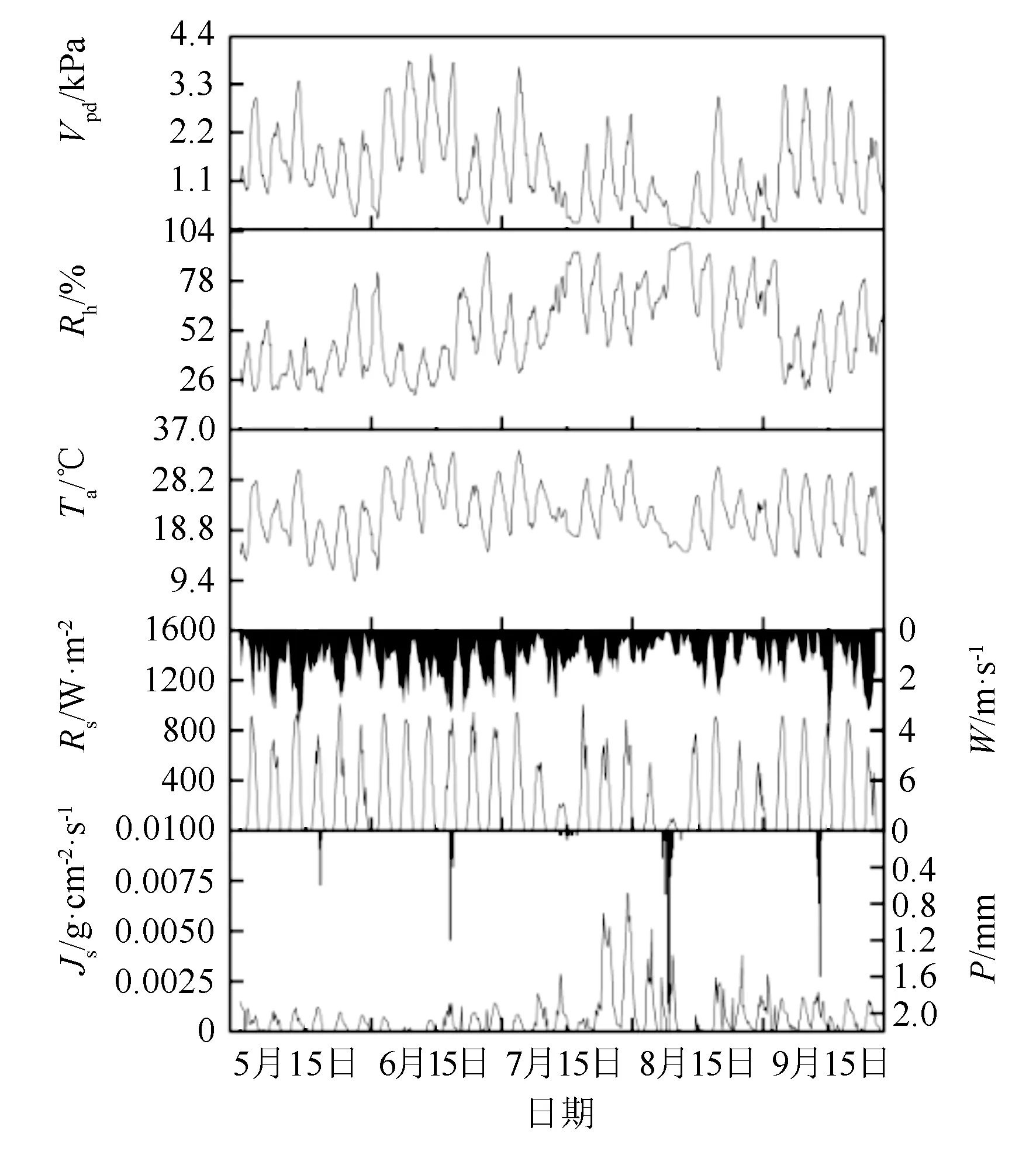
图1 丝绵木树干液流密度与生长季(5—9月)各气象因子变化情况
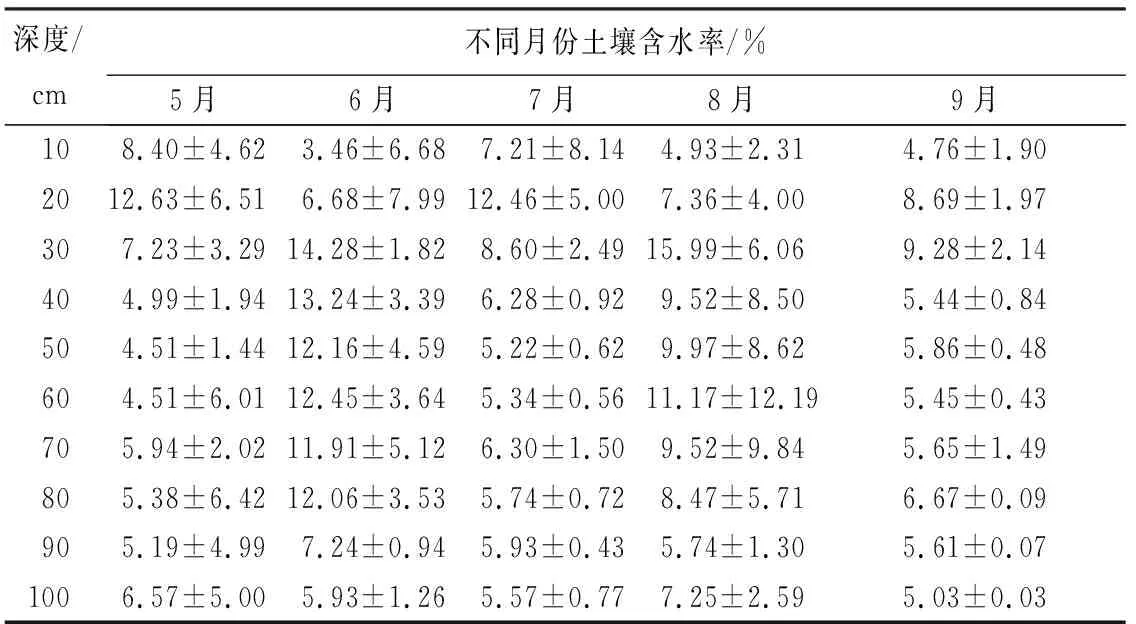
表2 不同深度土壤含水率
3.2 树干液流变化特征
丝绵木样木液流密度随太阳辐射变化而变化,树干液流的日变化呈典型的单峰曲线(图2),树干液流密度随着太阳辐射的升高而上升,12:00—13:00处达到峰值,之后随太阳辐射减小温度降低而逐步降低,并在20:00后逐步达到较低水平。此外,液流密度与太阳辐射变化存在时滞,约30 min。丝绵木前半夜的液流密度普遍高于后半夜(图3),说明夜间液流的变化主要存在前半夜。受太阳辐射驱动,树干液流于清晨06:00左右启动。从表3可知,7月丝绵木液流量最高,6月最低,从液流总量来看,6月与其他各月具有显著差异,而夜间液流量由大到小表现为8月、6月、7月、5月、9月,其中8月与5、6、7、9月具有明显差异;各月夜间液流占整日液流比例,由大到小分别为6月、8月、5月、9月、7月,土壤含水率增加夜间液流所占比例增加,夜间液流占整日液流量最高比例可达22.43%,因此,夜间液流是林木液流活动的重要组成部分,在研究中不可忽视。
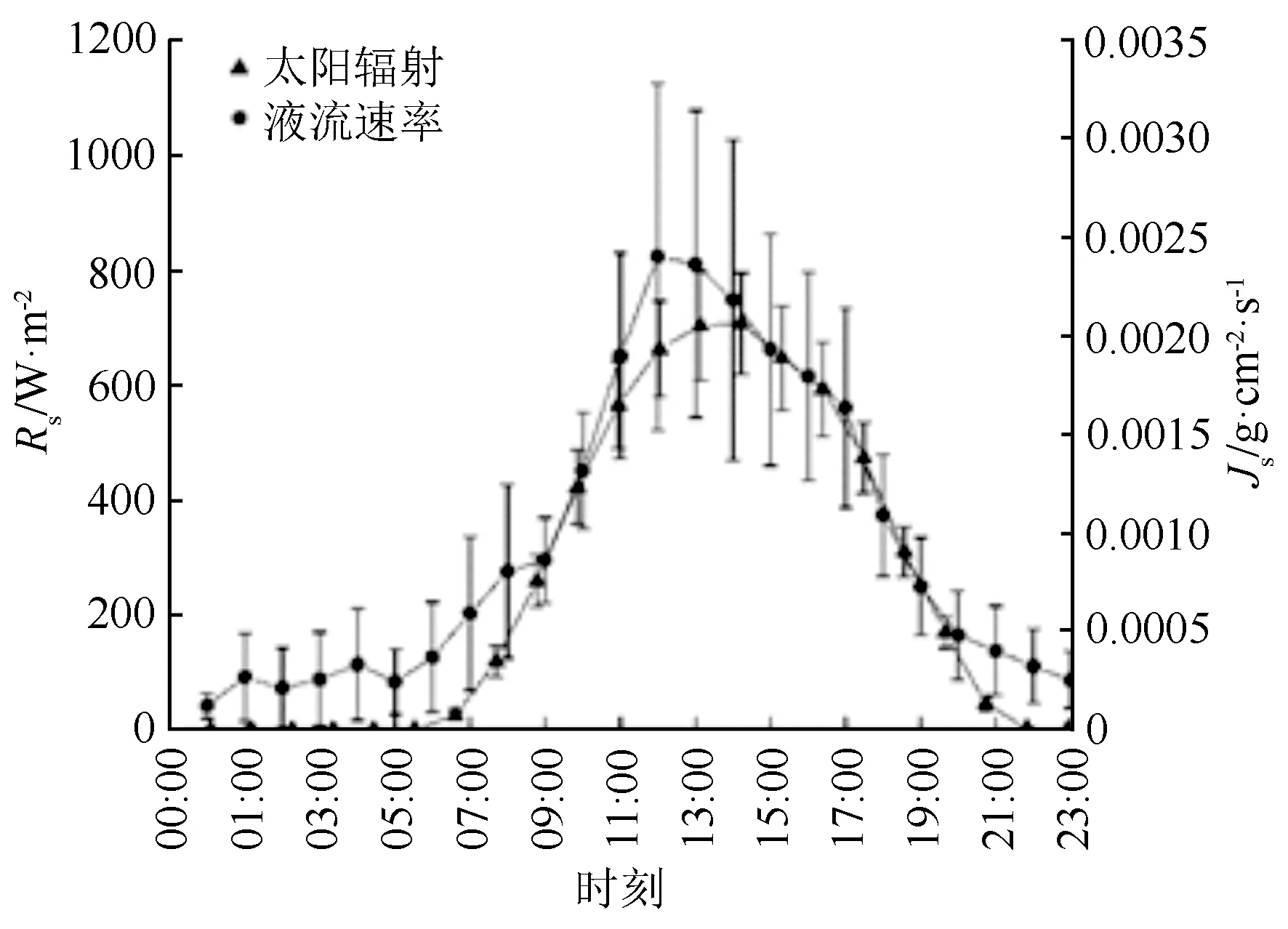
图2 丝绵木液流密度与太阳辐射变化
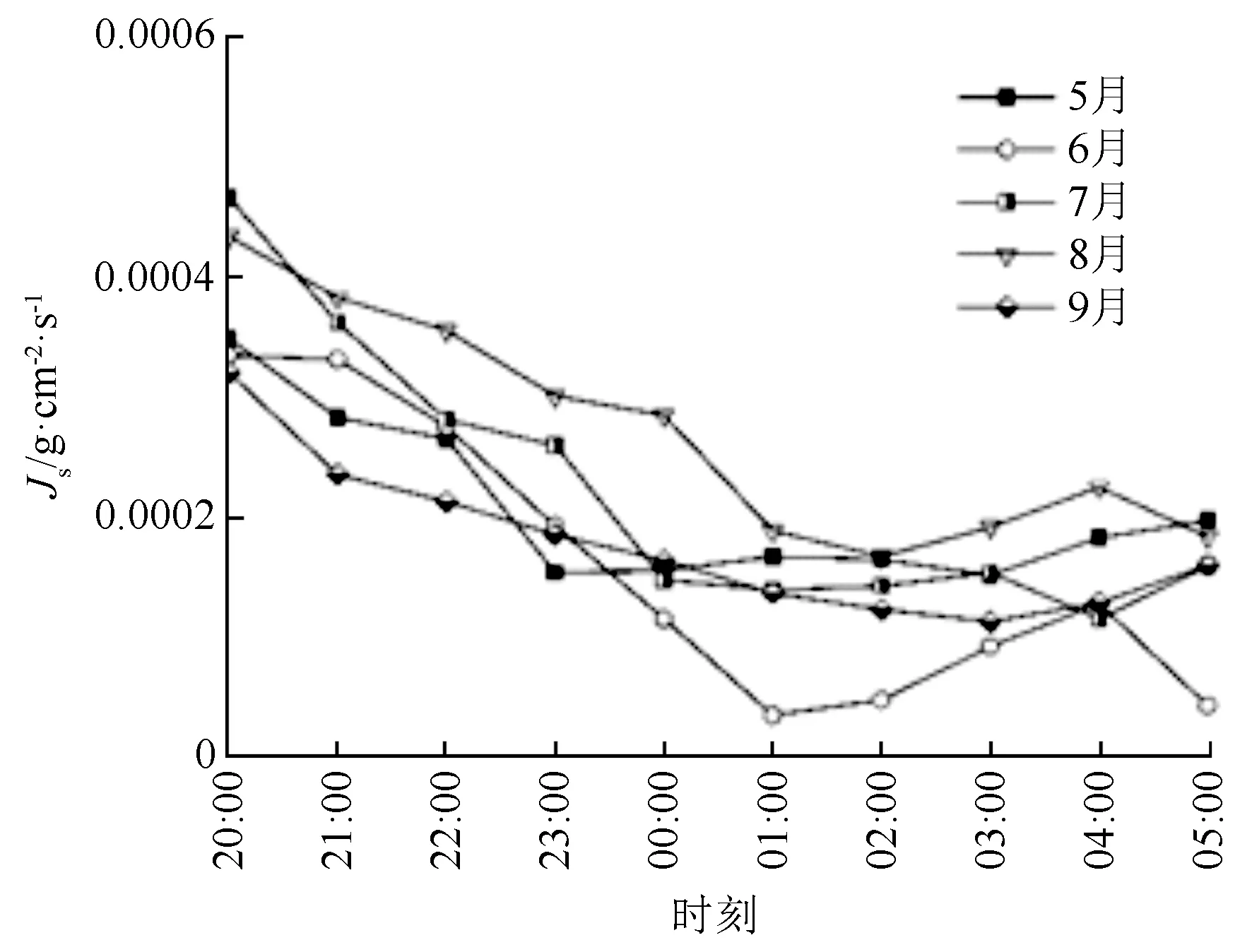
图3 丝绵木夜间液流密度均值变化
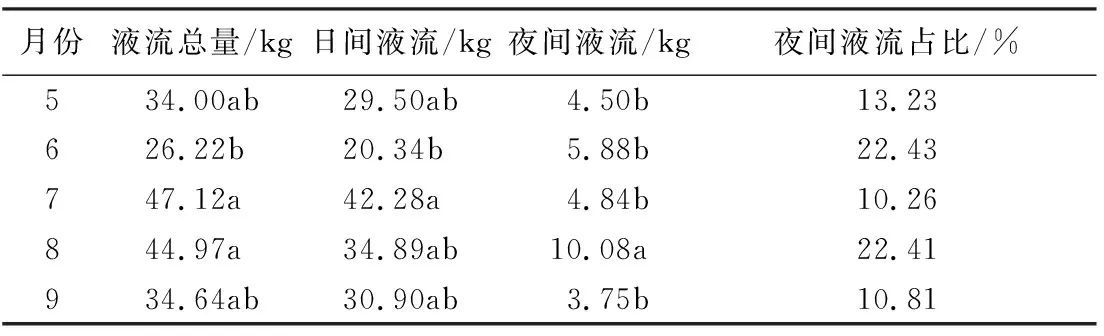
表3 丝绵木各月液流量变化
3.3 环境因子对树干液流的影响
3.3.1 昼夜液流与环境因子的关系
如表4所示,为丝绵木昼夜液流密度与气象因子的Pearson相关分析,日间液流密度与Rs、Ta、Rh、Vpd均表现极显著相关性,而夜间液流密度仅与Ta存在极显著相关,说明影响日间液流的主要因素是Rs、Ta、Rh、Vpd,夜间液流受Ta的影响较大。
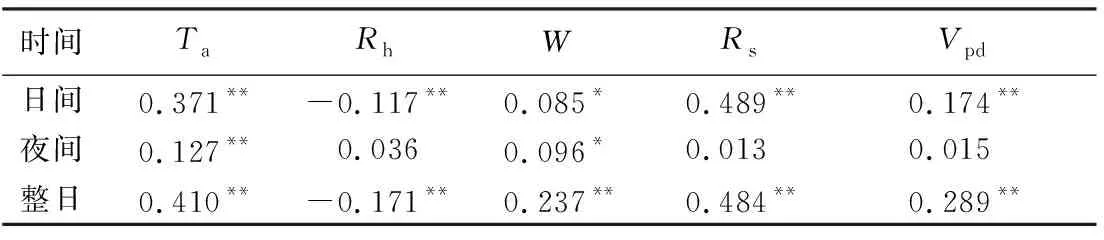
表4 丝绵木昼夜液流密度与气象因子的Pearson关系
3.3.2 日间液流对环境因子的响应
如图4、图5所示,丝绵木日间液流密度与Rs和Vpd关系,Rs和Vpd呈现正相关关系。Rs增大,液流密度随之增大,增速随Rs增加而减小;液流密度随Vpd增大而增大,并在1.7 kPa左右达到峰值,超过1.7 kPa时,液流密度回落并趋于稳定。
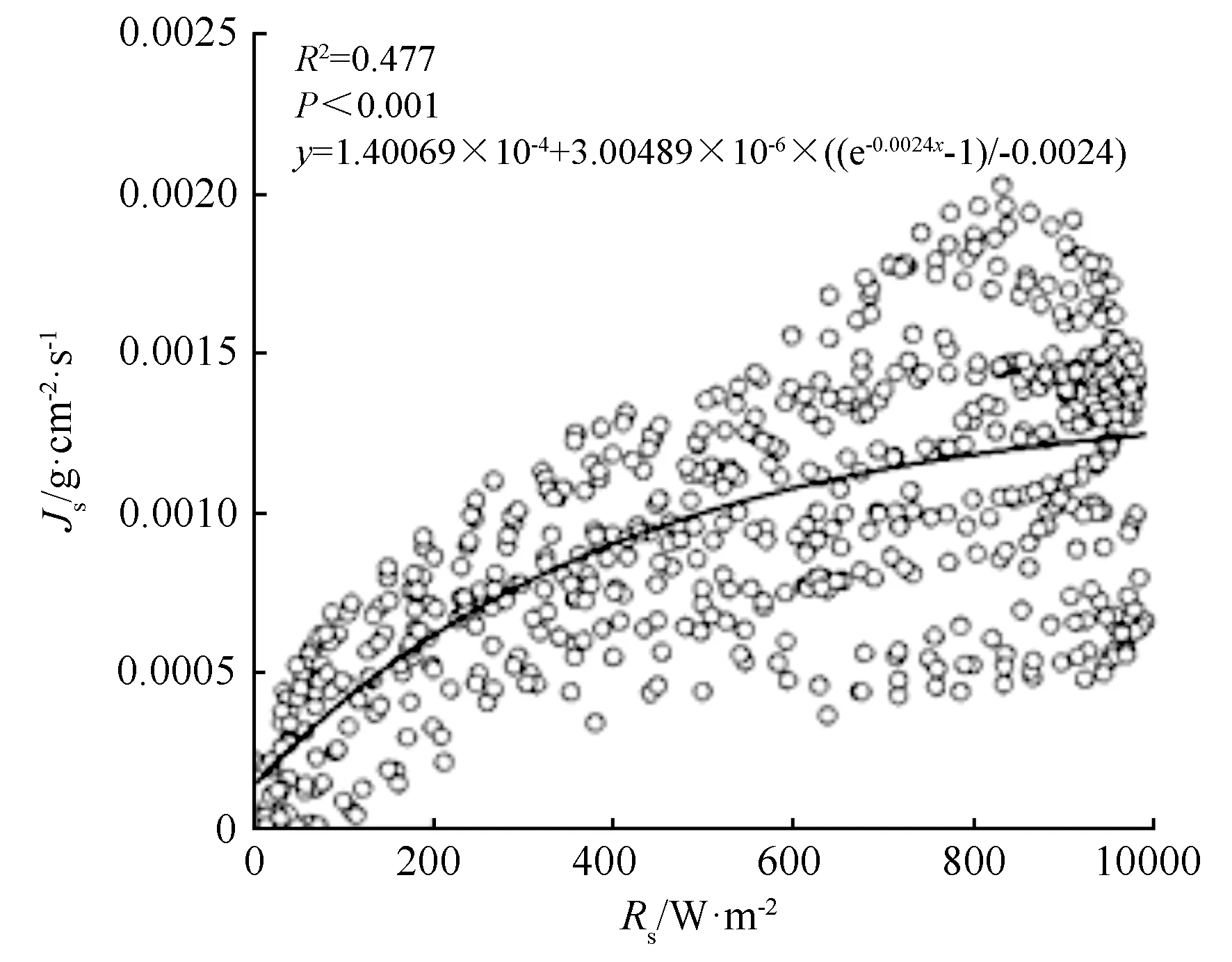
图4 丝绵木日间液流密度与Rs的关系
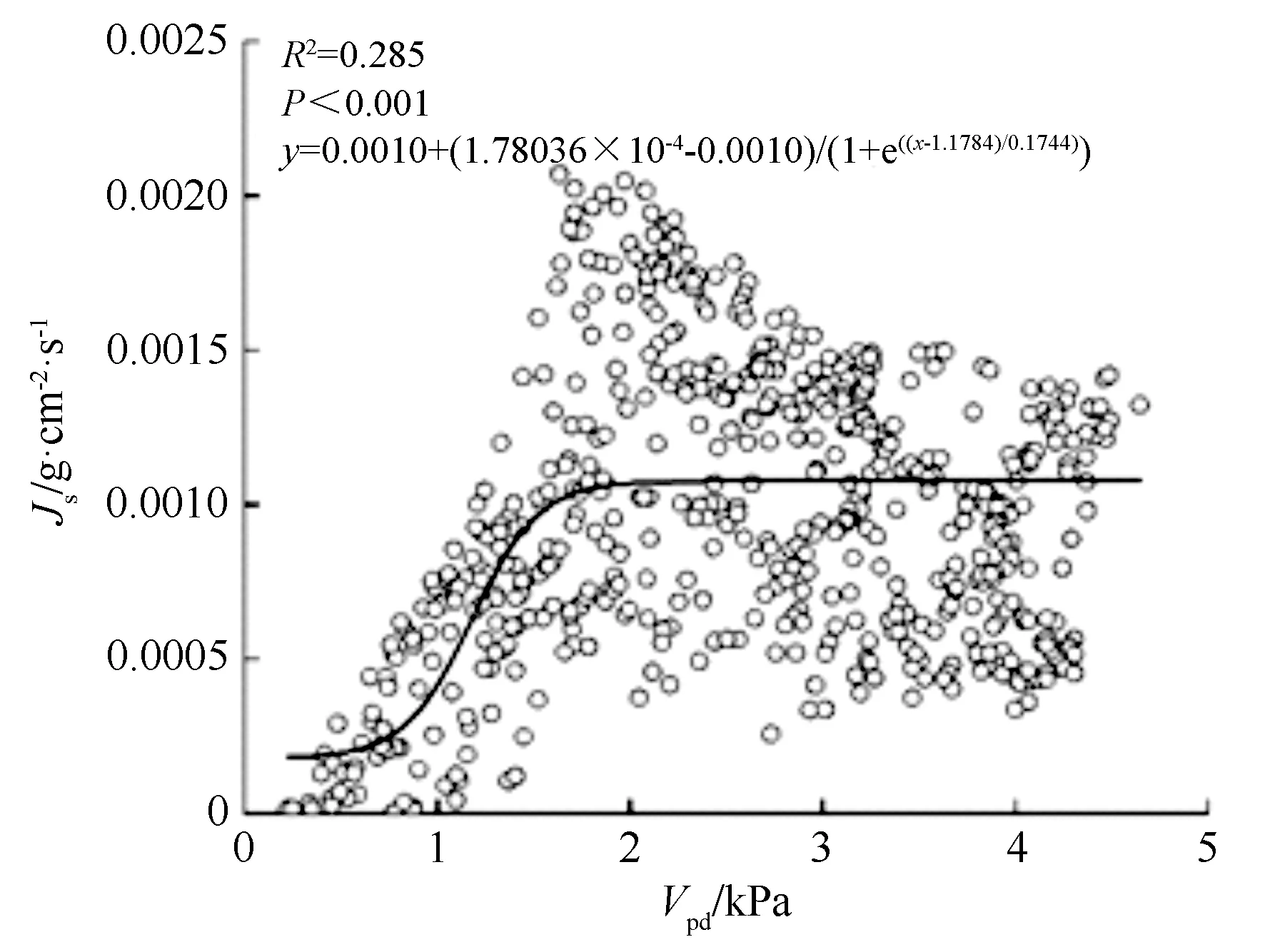
图5 丝绵木日间液流密度与Vpd的关系
3.3.3 夜间液流对环境因子的响应
在夜间影响因素中,根据Pearson相关系数的最优相关结果,选择Ta按1 ℃一个组别,分别对夜间液流进行方差分析,结果如表5所示,不同Ta组别下,夜间液流的总体均值显著地不同,说明在不同温度水平下,夜间液流具有明显差异;从图6可以看出,夜间液流与Ta呈现明显正相关关系,夜间液流随温度升高而增大。

表5 夜间液流密度与Ta的方差分析
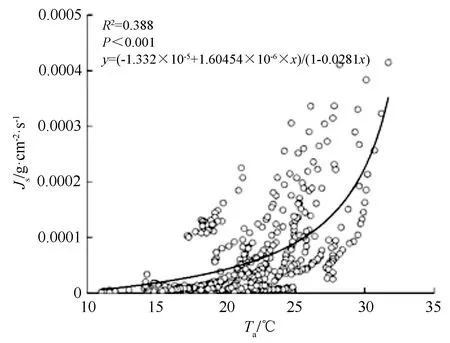
图6 夜间液流与Ta的关系
4 结论与讨论
树干液流是研究树木蒸腾过程、水分状况及估算单株耗水量的有力工具[23],树干液流受大气环境因素、生理生化特征、土壤水分环境等因素影响,呈现不同的昼夜变化节律。生长季丝绵木树干液流液在白天和夜间表现出较大的差别,白天液流速率呈现先增高后降低的趋势,夜间液流表现为前半夜较后半夜活跃,前半夜液流速率呈明显的下降趋势,后半夜则趋于平稳并在黎明前(06:00)开始启动。此外,树干液流启动时间与大气环境因子存在时滞效应[24-25],本研究发现,丝绵木树干液流启动时间、峰值时间与太阳辐射存在时滞现象,约30 min。谢恒星等[26]研究发现,槐树树干液流较太阳总辐射和光合有效辐射滞后10 min,较风速滞后20 min,王华等[27]在研究北京常见树种生长季时滞效应中发现,树干液流的变化与太阳辐射和饱和水汽压差显著相关且树干液流均滞后于太阳辐射、提前于饱和水汽压差,时滞时间在10~70 min;田原等[28]发现兴安落叶松(Larixgmelini)的树干液流与太阳辐射存在140~150 min滞后;而李雯雯等[29]研究结果表明,日尺度液流变化与环境因子的响应不存在时滞情况。王慧梅等[30]认为,液流时滞产生原因是当前树干液流密度的高低并不是由当前环境因子所直接决定,很可能受之前的环境因子或现存环境因子对之后的树干液流密度的明显影响,某一时刻的树干液流是受之前环境因子的相互影响,从而产生时滞现象,不同树种对环境因子变化的响应方式上的差异直接导致了液流速率格局的差异[31]。
丝绵木树干液流昼夜差异显著,日间液流是夜间液流的3~9倍,夜间液流占全天液流量的10%~22%,夜间液流不可忽视。张婕等[4]在研究北京山区元宝枫夜间液流活动特征及影响中发现,夜间液流月平均贡献率为12%,赵春彦等[1]则发现胡杨(Populuseuphratica)夜间液流占整日累计液流量的比例为26.2%~45.7%。在某些特定环境下,夜间液流所占的比例甚至更高[32]。越来越多研究表明,夜间液流一部分用于夜间蒸腾,一些植物不仅在白天维持着较高的蒸腾作用,在夜晚也保持着较高的气孔导度和气孔蒸腾[33];也有研究认为,林木夜间液流用于补充木质部白天蒸腾引起的木质部水分亏缺,这主要是由于树木存在的水容调节能力,使树体能够在夜间补充白天的蒸腾失水,避免树木体内产生气穴和栓塞[34]。此外,丝绵木夜间液流季节差异明显,由大到下依次为6月、8月、7月、5月、9月,丝绵木作为直根系树种[35],对水分敏感性强,在土壤含水率较高的月份(6月),有利于植物通过夜间液流补给日间耗水。
环境因子虽对树木的树干液流有着极其重要的影响,但树种差异性同样是液流变化不可忽视的重要因子之一[36]。李双等[37]对干旱区雨养多枝柽柳(Tamarixramosissima)液流对环境因子的响应研究表明,太阳辐射是影响柽柳白天液流速率的主导因子,温淑红等[9]则发现太阳辐射和气温对山桃(Amygdalusdavidiana)树干液流的影响较大,其次是空气相对湿度和风速。本研究发现丝绵木日间树干液流与太阳辐射、温度、饱和水汽压差呈极显著正相关关系,与大气相对湿度呈极显著负相关关系。太阳辐射作为蒸腾作用的主要驱动力,决定植物叶片气孔的张开程度,无论在何种时间尺度上,太阳辐射和大气温度都是影响树干液流的主要环境因子[38-39]。王志超等[40]研究得出尾叶桉(Eucalyptusurophylla)无论是旱季还是雨季,其夜间液流密度均与夜间饱和水汽压差呈极显著正相关,而王艳兵等[41]发现华北落叶松夜间树干液流与饱和水汽压差并不相关。本研究结果表明,丝绵木夜间液流密度与大气温度具有显著的相关性,夜间液流随温度升高而升高,在不同温度水平下具有显著差异。在气候变化背景下,夜间温度、降雨量等环境条件的变化将会影响到夜间液流过程以及夜间液流在蒸腾与茎干储水间的分配[32],使得植物蒸腾对水量平衡影响的不确定性也更大。地区和树种对树干液流分布规律及其与气象因子的关系影响很大[42],系统研究气象因子与树干液流的关系对于量化林木蒸腾耗水、阐释林木蒸腾调控机制意义重大。
