霍山石斛类黄酮-3-O-糖基转移酶(DhUF3GT)基因克隆与原核表达分析*
2022-08-26瞿彩丽刘雨馨李永华汪文杰
瞿彩丽,刘雨馨,李永华,汪文杰,余 坤,龚 玲
(湖北中医药大学药学院 武汉 430065)
霍山石斛(Dendrobium huoshanenseC. Z. Tang et S.J.Cheng)被赞誉为“九大仙草”之首,是药用石斛中的珍品,具有滋补肝肾、补气养阴、益津明目等功效[1]。由于霍山石斛对生长环境严苛,加之掠夺性采挖,野生资源几乎难觅踪迹[2]。自上个世纪六七十年代,地处于大别山腹地中心的湖北省英山县就已经开展了霍山石斛的仿野生人工培育,优良种质选育及人工种植技术相对成熟[3],在一定程度上缓解了霍山石斛资源匮乏等问题。然而,目前霍山石斛人工栽培的产量远远无法满足日益扩大的市场需求。这严重制约了霍山石斛深度的开发利用及产业化发展。因此,在加大霍山石斛人工种植技术研究的同时,有必要加快对霍山石斛关键活性成分的生物合成机制研究。
霍山石斛作为药用石斛中的珍品,富含新西兰牡荆苷、夏佛塔苷、槲皮素、柚皮素、芹菜素等类黄酮物质[4-5],除能清除体内的氧自由基外,还有较强的抗氧化、降血糖、抗肿瘤、抗炎等药理活性[6-7]。现代药理研究已证明,植物中大多数类黄酮化合物主要以类黄酮-O-糖苷或C-糖苷形式发挥广泛的药理作用[8-9]。如芹菜素7-O-葡萄糖苷能抗痉挛、抗炎[10];矢车菊素-3-O-葡萄糖苷能增强人肝癌HepG2 细胞的抗肿瘤活性[11];山奈酚3-O-葡萄糖苷对HIV-1表现出强抑制性[12];柚皮素7-O-葡萄糖苷具有抗癌、抗糖尿病等药理作用[13]。
类黄酮糖基转移酶(Flavonoid glycosyltransferase,UFGT)是类黄酮化合物生物合成下游途径中的最后一个合成酶,主要负责类黄酮糖基化修饰过程,能将具有活性的糖供体转移到黄酮醇、花色素、黄酮、异黄酮和黄烷醇等苷元上,形成种类多样的类黄酮糖苷类物质[14](图1)。因此,它被认为是类黄酮生物合成途径中的关键酶。近年来,研究者们从许多色泽鲜艳的植物叶片、茎、花朵中提取到丰富的花青苷及其合成酶UFGT[15-16],也认识到通过挖掘高效、专一的类黄酮UFGT 合成高质优产的类黄酮糖苷是类黄酮糖苷药源的重要途径。研究者通过随机突变人参皂苷Rh2 生物合成的关键酶编码基因UGTPg 45,获得了催化活性最高的糖基转移酶UGTPg 45-HV突变蛋白,使人参皂苷Rh2 的产量有效提高10 倍以上[17]。Wang 等发现野黄芩中的类黄酮3-O-糖基转移酶Sb3GT1 能催化17种黄酮醇转化成相应的3-O-糖苷,转化率高达到98%以上[18]。这为黄芩活性成分的体外生物合成提供了实验依据。尽管,研究者们前期做了大量关于霍山石斛多糖生物合成机制调控研究[19],但有关霍山石斛类黄酮糖基化反应的关键——类黄酮糖基转移酶UFGT 的研究却鲜少报道。
因此,本研究根据课题组前期霍山石斛转录组测序数据,筛选了霍山石斛类黄酮-3-O-糖基转移酶(DhUF3GT)基因,采用RT-PCR 技术克隆其基因序列,并进行生物信息学分析及其原核蛋白表达研究,以期为霍山石斛类黄酮-3-O-糖基转移酶的功能以及类黄酮化合物生物合成途径的阐明奠定基础。这也为将来利用生物工程等手段培育高含量黄酮成分的霍山石斛品种和促进霍山石斛产业可持续开发提供重要的理论基础。
1 材料与方法
1.1 材料
实验材料为课题组前期组织培养的霍山石斛新鲜叶片,经湖北中医药大学药学院余坤教授鉴定,为兰科石斛属植物霍山石斛Dendrobium huoshanenseC.Z. Tang et S. J. Cheng。用无菌清水将叶片洗净后消毒,滤纸吸干水分,作为提取霍山石斛RNA 的材料。
1.2 试剂
本实验所用试剂见表1。
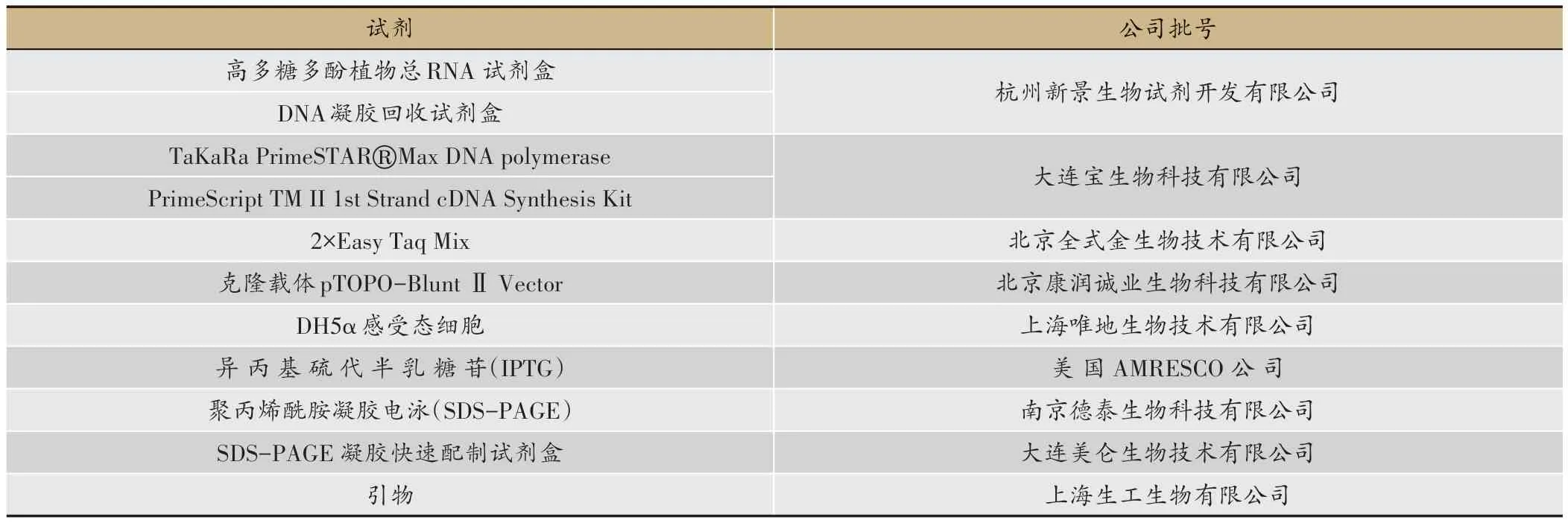
表1 实验试剂列表
2 方法
2.1 霍山石斛总RNA 的提取及cDNA 的合成
采用高多糖多酚植物总RNA 提取试剂盒提取霍山石斛叶片总RNA,并在1%琼脂糖凝胶电泳下检测总 RNA 的完整性。ND2000(Denovix DS-11+,S-03512)检测霍山石斛总RNA 的质量,选择A260/A280为 1.8~2.0 的总 RNA 按照 PrimeScript TM II 1st Strand cDNA Synthesis Kit反转录体系进行cDNA的合成。
2.2 霍山石斛DhUF3GT 基因序列的克隆
根据课题组前期霍山石斛转录组数据获得的DhUF3GT基因序列,运用在线软件NCBI ORF Finder设计特异性引物:GT-Z:CGGACAGGCAGTGGTAGGAAGAG;GT-F: TAAGAGAGAGTTCAGAGGGGGCT。以反转录的cDNA 为模板进行RT-PCR 扩增。cDNA 扩增在TaKaRa PrimeSTAR®Max DNA polymerase 作用下进行,反应条件为 98℃、3 min;98℃、10 s;55℃、15 s;72℃,30 s。2~4 步反应 30 个循环;72℃延伸 10 min;4℃永久。扩增产物经1%琼脂糖凝胶电泳检测后,切胶回收目的条带,并与pTOPO-Blunt ⅡVector 载体连接,转化至DH5α感受态细胞中,选择单克隆菌落并进行菌液PCR 验证,将阳性克隆菌液送至上海生工生物有限公司进行测序。
2.3 霍山石斛DhUF3GT基因生物信息学分析
霍山石斛DhUF3GT基因序列采用NCBI在线软件BLAST进行相似性搜索;采用DANMAN软件寻找开放阅读框(ORF),并对其氨基酸序列进行蛋白同源性分析;采用ExPASy、SOPMA 分析蛋白的理化性质;采用SignalP 4.1 Server 预测信号肽;采用 STRING 软件,以拟南芥蛋白数据库作为参考分析DhUF3GT 蛋白与其他蛋白的互作关系;蛋白的磷酸化和糖基化位点采用NetPhos 3.1 Server和Net NGlyc 1.0 Server进行预测;蛋白亲/疏水性采用ExPasy-ProtScale 分析;采用PSORT、SOPMA 和Swiss-Mode Workspace 分别进行蛋白的亚细胞定位和二、三级结构预测;采用PyMOL 软件对三级结构进行可视化处理;采用TMHMM Server v.2.0 进行蛋白跨膜结构域的预测;使用MEGA6.0 软件,应用Neighbor-joining(N-J)的方法建立NJ tree。
2.4 霍山石斛DhUF3GT基因的原核表达分析
根据测序后的结果设计特异性引物:正引物GAGCTCATGCCATCCACCGCCGCCTTCGCC;反 引 物GTCGACTCAAGTCTCCCCGGACTCGTTCAAC(下划线分别为SacⅠ、SalⅠ酶切位点),进行PCR 扩增。PCR产物与pTOPO-Blunt ⅡVector 的载体连接,构建重组质粒DhUF3GT-pTOPO,并提取阳性克隆重组质粒DNA。
理论计算所用原料为贵州赫章鲕状赤铁矿,化学成分如表1所示:鲕状赤铁矿原矿中的铁主要以赤铁矿的形式存在,少部分以褐铁矿形式存在,脉石成分主要为高岭石。
采用SacⅠ、SalⅠ限制性内切酶分别对DhUF3 GT-pTOPO 和表达载体pET28(a)进行双酶切,切胶回收目的基因片段与原核表达载体pET28(a),通过T4连接酶进行连接,菌落PCR 验证。重组表达质粒DhUF3GT-pET28(a)经快速质粒 DNA 提取试剂盒提取后,转化至Rosetta 表达感受态细胞中,IPTG 16℃诱导表达,通过12%SDS-PAGE电泳分析。
3 结果
3.1 霍山石斛总RNA提取结果
霍山石斛总RNA 提取后经凝胶电泳分析,结果如图 2,在约 1500 bp 和 1000 bp 处可见 28S 和 18S 两条清晰条带。总 RNA 的 A260/A280 为 1.986(1.8 至 2.0 之间),证明其质量和完整性均较好,无DNA 污染,且浓度为58.012 mg·μL-1。
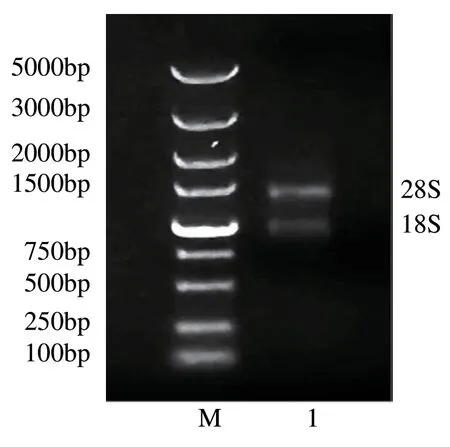
图2 霍山石斛总RNA凝胶电泳图
3.2 霍山石斛DhUF3GT基因序列的克隆
根据转录组数据中筛选的DhUF3GT基因设计专一性引物,以霍山石斛叶片RNA 逆转录的cDNA 为模板进行PCR 扩增,得到了霍山石斛DhUF3GT基因的片段。如图3显示,该凝胶电泳结果在约1300 bp处有一明显条带。将该片段条带切胶回收后进行克隆测序,最终测序结果为1289 bp,与前期转录组数据一致,表明该扩增片段是预期目的基因。
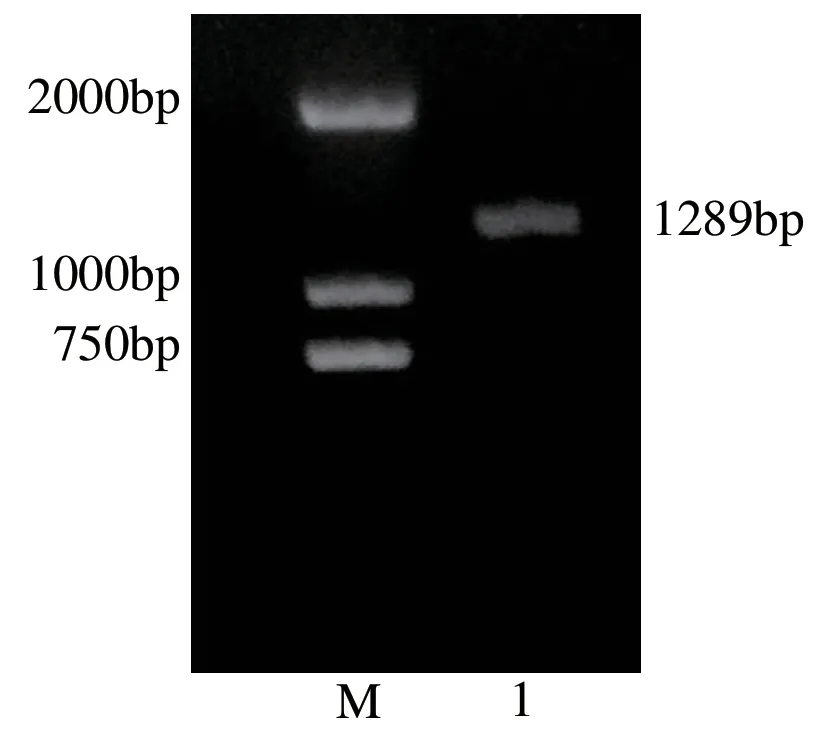
图3 霍山石斛DhUF3GT基因的凝胶电泳图
3.3 霍山石斛DhUF3GT基因生物信息学分析
3.3.1 序列分析
将DhUF3GT基因序列采用DNAMAN 在线软件进行ORF 框查询,发现该序列的开放阅读框为1239 bp,编码了412 个氨基酸(图4)。在NCBI 中注册,GenBank 中的登录号为OM460827。通过BLAST 搜索比对得到与该序列相似性最高的是同为兰科草本植物的铁皮石斛(Dendrobium catenatum)的糖基转移酶基因。

图4 霍山石斛DhUF3GT基因的cDNA序列
3.3.2 霍山石斛DhUF3GT基因编码蛋白质的理化性质分析
利用在线软件Protparam 对霍山石斛DhUF3GT基因编码的氨基酸进行序列分析,结果显示DhUF3GT蛋白总平均疏水性为-0.001,具有亲水性和不稳定性,为亲水性蛋白。理论相对分子质量为45.668 kD;分子式为 C2060H3216N552O584S19;等电点 PI 为 5.98;不稳定指数为48.17;脂肪族指数为92.82。序列所包含的412个氨基酸中,丙氨酸Ala 和亮氨酸Leu 含量最高,分别占比9.7%,其次是谷氨酸Glu 和半胱氨酸Cys 分别占8.0%和1.0%,其中负极性残基(Asp+Glu)45个,正极性残基(Arg+Lys)37个。
使用NCBI Concserved Domain 在线软件分析发现DhUF3GT基因编码的蛋白有3 个活性位点(色氨酸Trp-22,天冬酰胺Asp-23,组氨酸His-44),为糖基转移酶GTB 超家族成员(Glycosyltransferase_GTB-type super family)(图5a)。
使用在线软件,导入拟南芥蛋白数据库分析DhUF3GT 蛋白与其他蛋白的互作关系,得出DhUF3GT 与AT5G64540互作性最高,得分为0.903,其次是与AT1G14710(属富羟脯氨酸糖蛋白家族的氧化还原酶蛋白)和AT1G55270(属半乳糖氧化还原酶家族的半乳糖氧化酶)互作性较高,另外还与AT1G6020 0、AT2G29210、TIL 有相互作用。综上证明,DhUF3GT蛋白可能与其他氧化还原酶蛋白共同完成了类黄酮苷类物质的合成(图5b)。

图5 霍山石斛DhUF3GT基因编码蛋白质的理化性质分析
3.3.3 霍山石斛DhUF3GT 蛋白的磷酸化/糖基化位点、信号肽、亲/疏水性、跨膜区预测
经 NetPhos 3.1 Server 和 Net NGlyc 1.0 Server 软件分别预测,结果显示DhUF3GT 蛋白有20 个Ser磷酸化位点、9 个 Thr 磷酸化位点及 2 个 Tyr 磷酸化位点(图6a),以及两个糖基化位点(96、407)(图 6b)。经SignalP 4.1 Server 分析,DhUF3GT 蛋白为胞内蛋白(图6c),无信号肽。利用ExPasy-ProtScale 分析该蛋白的亲/疏水曲线,发现疏水性最强出现在第223 个氨基酸处,最高值为2.333;第147 个氨基酸的亲水性最强,得分为-2.856(图6d),且整个蛋白质链的大部分估计值小于0,故可以得到霍山石斛DhUF3GT 蛋白为亲水性蛋白。TMHMM Server v.2.0 软件分析显示,该蛋白没有跨膜区。PSORT 的亚细胞定位发现DhUF3GT 蛋白在叶绿体基质、类囊体膜和类囊体间隙,估值分别为0.938、0.775 和 0.750,因此推测 DhUF3GT 蛋白可能存在于叶绿体中。

图6 霍山石斛DhUF3GT蛋白
3.3.4 霍山石斛DhUF3GT 蛋白的二级和三级结构预测
采用SOPMA 和Swiss-Mode Workspace 在线软件估测DhUF3GT 蛋白的二级结构与三级结构。结果显示该蛋白二级结构中α-螺旋结构占41.99%,β-转角结构占5.83%,延伸链占14.81%,无规则卷曲占37.38%。研究可得出DhUF3GT 蛋白的二级结构主要属于α-螺旋(图7),三级结构经PyMOL 软件可视化处理,结果如图8。

图7 霍山石斛DhUF3GT蛋白的二级结构

图8 霍山石斛DhUF3GT蛋白三级结构及保守结构域
3.3.5 霍山石斛DhUF3GT基因编码氨基酸的同源性及系统发育树分析
DhUF3GT基因编码氨基酸的多重比较发现(图9),霍山石斛DhUF3GT基因编码蛋白与铁皮石斛(Dendrobium catenatum)、石斛兰杂交种(Dendrobiumhybrid cultivar)、小兰屿蝴蝶兰(Phalaenopsis equestris)、海枣(Phoenix dactylifera)、油棕(Elaeis guineensis)及小麦(Triticum aestivum)中的糖基转移酶蛋白共同指数分别为 98.79%、89.76%、79.23%、48.95%、47.32%、48.06%,且都在C 末端含有1个高度保守的PSPG 结构域(图9)。选取这6 个物种的糖基转移酶氨基酸序列采用 MEGA6.0 构建 NJ 树,发现霍山石斛 DhUF3GT 蛋白与同科植物铁皮石斛的亲缘关系最高(图10)。

图9 霍山石斛DhUF3GT氨基酸序列与其他植物的多重比对
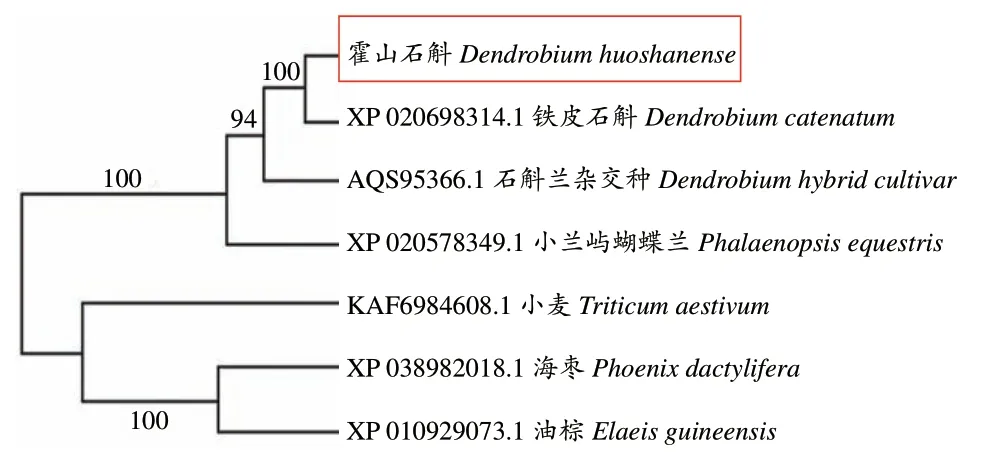
图10 霍山石斛DhUF3GT氨基酸序列NJ树
3.4 霍山石斛DhUF3GT基因原核表达
3.4.1 重组表达质粒构建及验证
将构建的DhUF3GT-pET28(a)重组质粒进行双酶切,结果显示(图11):双酶切后在约5000 bp 和1300 bp 处有明显条带,且1300 bp 处条带与霍山石斛DhUF3GT条带位置一致,证明重组质粒构建成功。
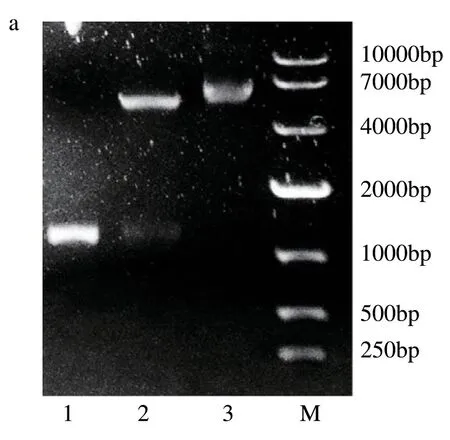
图11 重组质粒DhUF3GT-pET28aD及双酶切验证
3.4.2 SDS-PAGE蛋白电泳检测
蛋白诱导后经电泳检测结果显示(图12),与pET28(a)空载体表达总蛋白相比,诱导后的总蛋白、上清及沉淀都在45 KDa处有明显条带,证明该蛋白成功表达。且上清蛋白条带较沉淀中蛋白深,说明原核表达蛋白主要存在于上清中。综上证明,霍山石斛DhUF3GT蛋白在原核细胞中成功表达,且表达趋于折叠正确的活性蛋白形式。
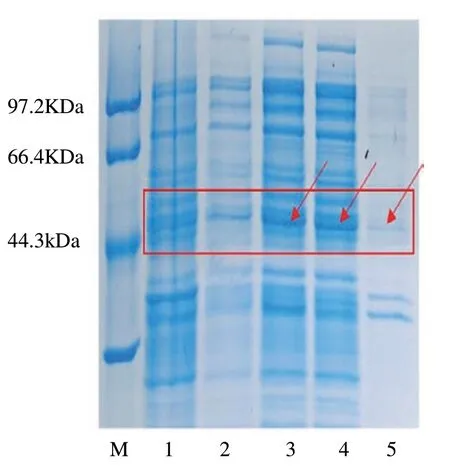
图12 蛋白SDS-PAGE凝胶电泳图
4 讨论
目前,整个霍山石斛产业的发展呈“头重脚轻”的趋势,即药材炮制技术(枫斗)工艺成熟、历史悠久,但中成药、新型饮片、保健食品等深加工产品数量稀少,技术创新非常薄弱[20-21]。与同属的铁皮石斛、紫皮石斛、金钗石斛相比,霍山石斛在本草考证、功能主治、化学成分和药理研究等深入宣传推广上差异化不显著,其独特的文化内涵也尚未充分挖掘[22]。归根究底,霍山石斛产业的发展受到上游基础研究不充分的严重制约,如①药效学物质基础、作用机制研究薄弱;②有别于其他品种的独特化学成分和药理活性未深入研究[23],专一的质控标准缺乏。霍山石斛类黄酮合成途径和调控机制研究尚未充分解析,这严重影响了下游产业的深度研发与创新。
植物类黄酮的生物合成以丙二酰辅酶A 和香豆酰CoA 为底物,在一系列酶的作用下形成黄酮、黄酮烷、黄酮醇、花青素等苷元和黄酮苷(图1)。随着生物信息学的飞速发展,大量类黄酮-3-O-葡萄糖基转移酶(3GT)基因在拟南芥、矮牵牛、金花茶、丹参等重要模式植物和园艺作物中被发现[24-27]。如在拟南芥(Arabidopsis thaliana)中,山柰酚、槲皮素、异鼠李素等在UGT78D1 作用下,生成3-O-鼠李糖苷[24];矮牵牛中的黄酮醇、花色素在PGT8 催化下能分别生成黄酮醇3-O-葡萄糖苷、花色素3-O-葡萄糖苷[28]。金花茶中CnUFGT14和CnUFGT15基因参与黄酮醇、花青素的修饰反应[26];丹参的SmUF3GT基因调控了花青素苷的合成[29]。由此可见,3GT基因为类黄酮糖苷修饰合成的关键基因。
本研究从霍山石斛中成功克隆出类黄酮-3-O-糖基转移酶(DhUF3GT)基因序列,经在线生信工具分析发现该基因片段包含一个长度为1239 bp 的开放阅读框,能编码412 个氨基酸。DhUF3GT 蛋白属于亲水性
稳定蛋白,理论相对分子质量为45.668 KD,等电点(PI)为5.98。氨基酸同源性分析显示,霍山石斛与其他植物糖基转移酶氨基酸序列相似性较高,与同科植物铁皮石斛相似性可达98.79%,且DhUF3GT 蛋白在C 端具有UGT 的特征保守结构域“PSPG Box”。这证明了本次克隆得到的序列准确,属于糖基转移酶UGT家族基因一员。
有研究表明,PSPG 序列中特定位点的氨基酸决定了该糖基转移酶糖基供体的选择。在霍山石斛DhUF3GT 蛋白的PSPG 保守序列中,第22 位为色氨酸、第23 位为天冬酰胺和第44 位为组氨酸,表明其可能利用葡萄糖和半乳糖来合成花青素苷等类黄酮物质。由此推测,霍山石斛DhUF3GT与其他同家族的糖基转移酶有着相似的葡萄糖基和半乳糖基的糖基化修饰活性[23]。
据相关研究报道,植物的糖基转移酶一般定位于细胞质基质,主导了植物次生代谢产物的合成[25]。霍山石斛DhUF3GT基因经PSORT 的亚细胞定位分析,该基因主要分布在叶绿体中。由于类黄酮转移酶选择糖基供体或受体时具有特异性,导致亚细胞定位部位有所不同,该研究结果还需要进一步的实验证实。
霍山石斛与铁皮石斛具有较高的亲缘关系,两者均含有丰富的类黄酮化合物[30]。近年来,随着铁皮石斛基因组图谱的成功绘制,Ren 等[31]成功挖掘及鉴定出一个新颖的黄酮碳苷糖基转移酶基因,能体外合成牡荆素、异牡荆素等化合物。Yu 等[32]发现铁皮石斛UDP-葡萄糖黄酮-3-O-葡糖基转移酶基因(DoUFGT1、DoUFGT2)(MH663506、MH663505) 在红皮品系中的表达水平显著高于绿皮品系。与此同时,红皮品系中的飞燕草素3,5-O-二葡萄糖苷(Delphinidin 3,5-O-diglucoside)和花青素3-O-葡萄糖苷(Cyanidin 3-O-glucoside)含量要明显高于绿皮品系。而总黄酮含量和抗氧化性也明显高于绿皮茎。这些研究充分证明类黄酮糖基转移酶(UFGT)在石斛属植物黄酮合成中发挥了重要作用。
类似的研究,Feng 等[33]发现花青素类物质在红色玫瑰花瓣中显著积累,而白色突变体花瓣中的黄酮醇含量却显著高于红色花瓣。经转录组分析发现,9 个UFGT基因在白色玫瑰中显著上调,而另外6 个UFGT基因却显著下调。这表明类黄酮糖基转移酶(UFGT)能平衡类黄酮物质的合成。
本研究初步探索了霍山石斛类黄酮-3-O-糖基转移酶(DhUF3GT)原核表达条件。该蛋白能在原核细胞中成功表达,且表达趋于折叠正确的活性蛋白形式。鉴于霍山石斛和铁皮石斛亲缘关系最近,初步可推测参与类黄酮糖基化反应的该蛋白在结构表征和功能上与铁皮石斛具有高度的同源性。下一步实验将进一步筛选特异性好、催化活性较高的UF3GT,为构建霍山石斛类黄酮苷的生物合成路径打下基础。
5 总结
本研究成功克隆出霍山石斛类黄酮-3-O-糖基转移酶(DhUF3GT)基因序列,并获得其序列特征及原核表达蛋白。这为霍山石斛DhUF3GT基因功能的进一步研究和类黄酮化合物生物合成机制的阐述奠定了良好的实验基础。同时,本研究为今后采用生物技术提高霍山石斛药材黄酮成分含量,扩大霍山石斛产业化生产提供了重要的理论依据。
