高烟碱K326 定向改良材料的创制与分析
2022-08-22郑淑心闫筱筱王召军张洪映
郑淑心,闫筱筱,王召军,张洪映,崔 红
河南农业大学烟草学院,郑州市金水区文化路95号 450002
烟草生物碱是影响烟草品质的一类重要物质[1]。烟草生物碱主要分为烟碱、新烟碱、降烟碱和假木贼碱,其中烟碱含量(质量分数)占生物碱总量的90%~95%[2]。近年来,随着工业降焦技术的飞速发展,卷烟焦油量得到了有效控制,但同时卷烟产品的烟碱和致香物质含量也逐渐减少,导致卷烟产品的香气减弱[3-4]。而提高烟碱含量的烟叶可在降低焦油释放量的同时,最大限度地满足感官需求,受到工业企业的青睐。虽然,栽培措施对烟碱合成和积累有明显的调控作用[5-6],但仅依靠肥料用量、留叶数来调控烟碱,无疑会导致烟叶发育不正常以及化学成分的不协调[7-8]。近年来,CRISPR/Cas9 技术已实现了在多个物种中的应用。在烟草中,也相继实现了单基因定点突变和多基因靶点敲除,均表明CRISPR/Cas9 基因编辑系统的有效性和可行性。因此,采用基因编辑技术对烟碱合成相关基因进行编辑,实现烟碱代谢的定向调控,对培育高烟碱烤烟品种具有重要意义。
生物碱的合成过程不仅受喹啉酸磷酸核糖基转移 酶(Quinolinic acid phosphoribosyl trandaferase,QPT)、腐 胺- N -甲 基 转 移 酶(PutrescineN-methyltransferase,PMT)、鸟氨酸脱氨酶(Ornithine decarboxylase,ODC)和PIP 家族还原酶(A622)等关键酶类催化调控,同时也受茉莉酸(JA)信号途径的调节[9]。JAZ 作为JA 途径信号模块(SCFCOI1-JAZMYC2)的关键抑制因子[10-11],广泛参与调控植株免疫反应、生长发育和代谢物质的合成等过程,其中Zhang 等[12]、Howe 等[13]和Wang 等[14]的研究表明,烟草NtJAZ1参与调控烟碱的合成。另外,烟草JAZ基因沉默激活JA信号下游相关基因大量表达,显著提高植株对烟芽夜蛾的抗性[15]。但烟草NtJAZ1基因突变对烟草生长发育及品质的影响却鲜有报道。此外,拟南芥AtJAZ1基因在低温条件下能促进植株次生代谢的发生,提高植株的耐寒性,维持植株正常生长发育[16-17]。水稻JAZ可以调控碳水化合物的合成[18],在JA介导的白叶枯病抗性反应过程中发挥重要作用[19]。过表达JAZ9的水稻转基因株系通过调控离子平衡等显著提高植株的耐盐性[20]。因此,以普通烤烟品种K326为材料,通过CRISPR/Cas9技术敲除NtJAZ1基因获得定向改良的高烟碱材料,并对其农艺性状、烟碱含量、腺毛密度和叶面化学成分等进行了分析,旨在为培育优质烟叶原料提供依据。
1 材料与方法
1.1 试验材料
供试材料为本课题组保存的烤烟品种K326。K326 种子经消毒后播种在MS 培养基上,用于农杆菌的遗传转化。另外,K326突变材料播种于育苗盘上,按烟草行业标准进行育苗[21],于2020年5月移栽种植于河南许昌隔离封闭试验田。田间种植和管理按照烤烟栽培技术流程[22]和当地优质烟叶栽培规程进行。
1.2 试验方法
1.2.1NtJAZ1基因敲除载体的构建
根据茄科基因组网站(https://solgenomics.net/)已发表的烟草NtJAZ1基因两个拷贝的序列(Nitab 4.5_0000073g0270.1、Nitab4.5_0004234g0080.1),利用CRISPR2 在线软件(http://crispr.hzau.edu.cn/CRIS PR2/)设计gRNA 序列:5′-GCAGTAGAAATAACTT CTTG-3′(341~360 bp),合成Oligo 二聚体后连上CRISPR/Cas9 基因敲除载体。将该载体转化根癌农杆菌EHA105 中,根据叶盘转化法对K326 无菌叶片进行农杆菌侵染,同时以空载体进行阳性对照试验。
提取突变体和对照幼苗DNA,通过PCR扩增和靶位点测序分析其序列突变情况,使用的引物如表1所示。

表1 扩增引物序列Tab.1 Amplification primer sequences
1.2.2 植物学性状调查
在烟株现蕾期,按照烟草行业标准方法[23]分别测量烟草K326和NK-9的株高、叶数、叶长、叶宽、茎围和节距等农艺性状指标,每个烟草株系测量10株。
1.2.3 烟碱含量测定
在现蕾期(移栽后60 d)、打顶期(移栽后75 d)、和成熟期(移栽后90 d),分别选取K326 和NK-9 的中部叶(从下向上数10~12 叶位)测定烟碱含量(质量分数)。将叶片置于105 ℃杀青1 h,在85 ℃烘干至恒质量。将叶片研磨后过0.250 mm 孔径的样品筛,称取200 mg样品于甲基叔丁基醚(MTBE)中萃取24 h,转移上清液至气相色谱-氢火焰离子化检测器(7890A,美国安捷伦公司)中,按照行业标准方法[24]进行烟碱含量的定量分析,重复3次。
1.2.4 腺毛形态观察与统计
分别选取K326 和NK-9 中长度约10 cm 的叶片进行腺毛形态和密度观察。使用质量体积分数为0.2%的罗丹明B 溶液染色20~30 min,用无菌水清洗后,放置于超景深显微镜(VHX-5000,日本基恩士公司)下进行腺毛形态拍照和密度统计。
1.2.5 叶面化学成分含量测定
分别挑选5株生长状况良好的K326和NK-9,取其中部叶进行叶面化学成分含量测定,重复3次。通过二氯甲烷浸提、无水硫酸钠搅拌干燥,用定量滤纸过滤至圆底烧瓶中。经旋转蒸发后加入1 mL 内标(2.020 mg/mL的八乙酸蔗糖酯+2.542 mg/mL的十七烷醇混合溶液)定容于棕色容量瓶中,在氮气保护下将溶剂吹干。然后,加入500 μLV(N,N-二甲基甲酰胺)∶V(N,O-双三甲硅基三氟乙酰胺)=1∶1 的溶液,在75 ℃水浴中进行衍生化反应。最后,分别加入N,O-双乙酰胺和吡啶125 μ L,即为GC/MS 分析样品液。将样品液置于GC/MS分析仪(HP-5890,美国赛默飞世尔科技有限公司)中进行化学成分的定性和定量分析,具体方法参考文献[25]。
1.2.6 常规化学成分含量测定
分别选取K326 和NK-9 的C3F 烟叶于75 ℃烘箱中烘干并研磨成粉,并过0.250 mm 孔径的样品筛,称取0.25 g 烟末于三角瓶中,加入25 mL 体积分数为5%的醋酸溶液,共振荡萃取30 min,定性滤纸过滤后作为待测液,用连续流动分析仪(AA3,德国水尔公司)按照行业标准方法[26-28]测定水溶性总糖、还原糖、烟碱、氯和钾含量(质量分数),重复3次。
1.2.7 中性香气物质测定
参照赵铭钦等[29]的方法测定烤后烟叶(C3F)中性香气物质含量。
1.2.8 生物碱合成相关基因表达分析
利用植物总RNA 提取试剂盒(DP4332,天根生化科技有限公司)分别提取K326 和NK-9 的根系总RNA,利用反转录试剂盒(NovoScript®ⅡReverse Transcriptase,上海近岸科技有限公司)合成cDNA第一链。根据NCBI GenBank 中的烟碱合成相关基因(NtMYC2a、NtMYC2b、NtPMT、NtQPT、NtODC和NtA622)序列,利用Primer Premier 5.0 软件设计荧光定量PCR 引物,上下游引物见表1,以L25(L18908)为内参基因。RT-PCR 反应程序:95 ℃2 min;95 ℃15 s,58 ℃15 s,72 ℃20 s,40个循环;溶解曲线:95 ℃15 s,58 ℃15 s,20 min 内升至95 ℃,95 ℃15 s。重复3 次。
1.2.9 数据处理
数据采用SPSS 17软件进行单因素方差分析,用最小显著差数法进行差异显著性检验,用Sigmaplot 14.0软件进行图表绘制。
2 结果与分析
2.1 高烟碱K326突变材料的创制和分子鉴定
通过农杆菌介导法将NtJAZ1基因敲除载体导入K326,经过潮霉素抗性筛选后,共获得22 株阳性植株。分别利用NtJAZ1-1和NtJAZ1-2特异引物(表1)进行PCR扩增和产物测序后发现10株的gRNA序列出现碱基突变,基因编辑效率为45.45%。突变情况分析表明,NtJAZ1-1靶位点序列发生突变的株系为NK-3、NK-4、NK-8、NK-9、NK-12、NK-15 和NK-17,NtJAZ1-2靶位点序列发生突变的株系为NK-3、NK-4、NK-6、NK-9、NK-12、NK-15、NK-19 和NK-22。其中,NK-9 中均为纯合突变,且NtJAZ1-1靶位点序列在1587~1641 bp 处缺失55 个碱基,NtJAZ1-2靶位点序列在1547~1548 bp 缺失2 个碱基(图1)。

图1 NK-9株系的分子鉴定Fig.1 Molecular identification of NK-9 line
为验证NtJAZ1基因敲除对烟碱合成的影响,采用RT-PCR对相关烟碱合成基因进行表达量分析,结果如图2 所示。图2 表明,与K326 相比,NK-9 中NtMYC2a、NtMYC2b、NtPMT和NtQPT的相对表达量明显提高,但NtODC和NtA622的相对表达量无显著变化。

图2 NK-9株系相关基因的相对表达量Fig.2 Relative expressions of genes of NK-9 line
2.2 NtJAZ1 基因敲除对植物学性状及叶面化学成分的影响
分别对植株形态和农艺性状进行观察与调查,结果见表2和图3A。与K326相比,NK-9生长良好,株高、叶数和叶宽显著高于对照K326,叶长显著低于K326,茎围和节距差异不显著。腺毛形态观察和密度统计分析结果见图3B,与K326相比,NK-9腺毛总数为77.29 根/mm2,降低14.62%,长柄分泌型腺毛数量降低20.19%。对叶面化学成分分析(图3C)发现,NK-9 的叶面化学成分总量为39.70 μg/cm2,西柏烷类二萜含量为34.43 μg/cm2,明显低于K326,分别降低23.71%和25.32%。这表明NK-9 生长发育良好,NtJAZ1具有调控腺毛发育和叶面化学合成分泌的功能。

表2 NK-9株系的农艺性状指标①Tab.2 Agronomic trait indexes of NK-9 line

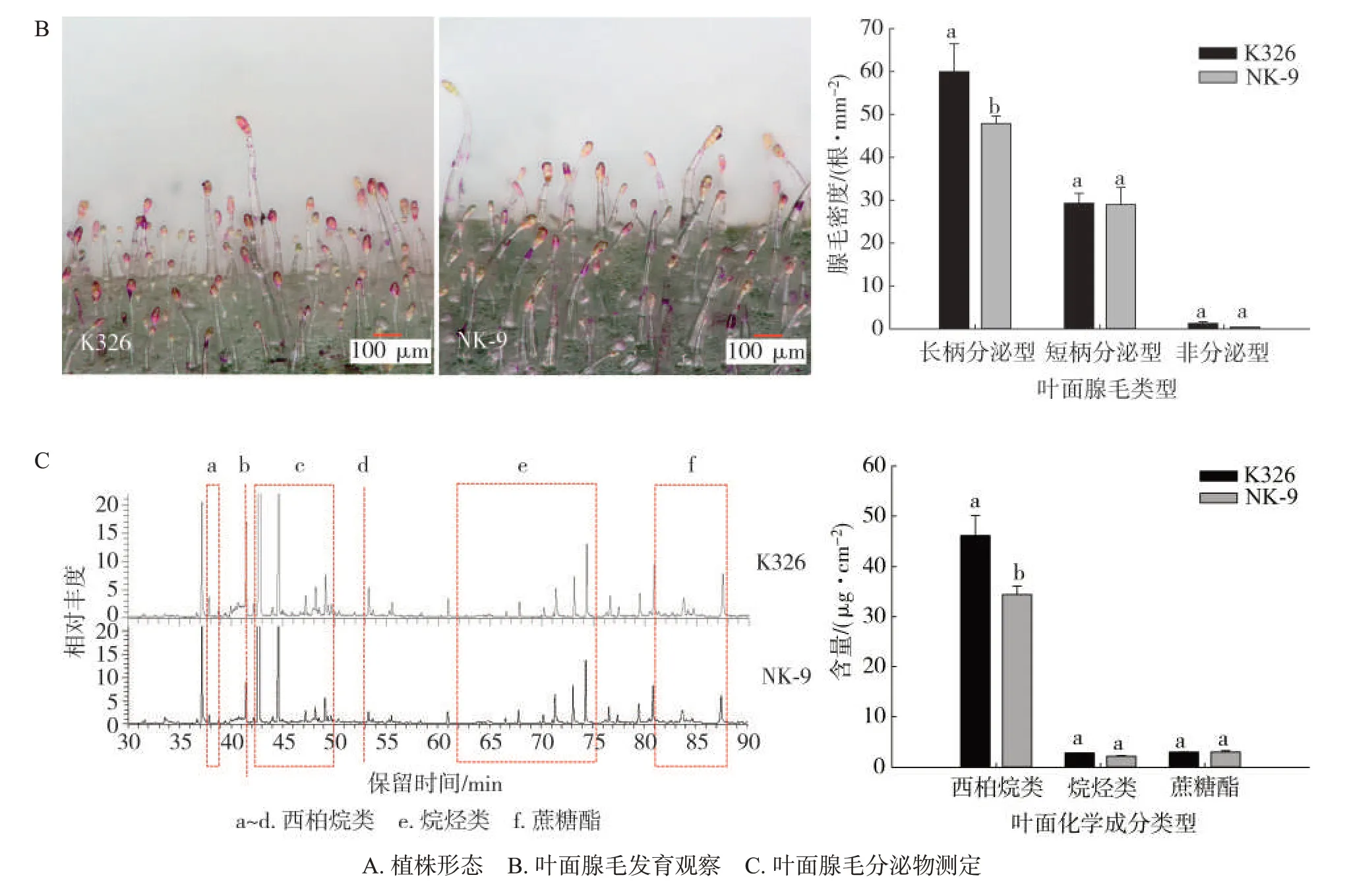
图3 NK-9株系的田间生长发育状况Fig.3 Growth and development of NK-9 line in fields
2.3 NtJAZ1基因敲除对烟叶烟碱含量的影响
分别在现蕾期、打顶期和成熟期对烟叶进行烟碱含量测定,结果见图4。与对照K326相比,随着烟株的生长发育,NK-9 烟叶烟碱逐渐积累,在现蕾期显著提高33.26%,在打顶期和成熟期分别极显著提高36.41%和39.28%。由此可见,NtJAZ1基因敲除后促进了烟株中烟碱的合成。

图4 NK-9株系的烟碱含量Fig.4 Nicotine content of NK-9 line
2.4 NtJAZ1基因敲除对烤后烟叶化学成分的影响
2.4.1NtJAZ1基因敲除对常规化学成分的影响
选取C3F 等级烟叶进行常规化学成分测定,结果(表3)显示:与对照K326 相比,NK-9 烤后烟叶烟碱含量提高44.91%,差异达极显著水平,氮碱比和糖碱比偏低,总糖、还原糖、总氮、氯和钾含量在适宜范围内。

表3 烤后烟叶化学成分含量Tab.3 Contents of chemical components in cured tobacco leaves
2.4.2NtJAZ1基因敲除对中性香气成分的影响
对C3F 等级烟叶进行香气物质含量分析,结果(表4)显示:共检测出棕色化反应产物、类胡萝卜素类降解产物、苯丙氨酸类降解产物、西柏烷类降解产物和新植二烯五大类。
从表4 可以看出,与对照K326 相比,NK-9 烟叶的新植二烯含量提高35.02%,类胡萝卜素类降解产物含量总体上增加,棕色化反应产物总量和西柏烷类降解产物含量略有降低,中性香气物质总量略高,达1060.35 μg/g。说明NtJAZ1基因敲除株系中的新植二烯和中性致香物质总量明显提高。

表4 烤后烟叶中性香气物质含量Tab.4 Contents of neutral aroma components in cured tobacco leaves (μg·g-1)
3 讨论
本试验中通过CRISPR/Cas9 技术对K326 中NtJAZ1基因序列进行编辑,导致该基因发生移码突变,翻译提前终止,获得了NtJAZ1基因敲除材料NK-9。对NK-9 根系中烟碱合成基因的表达量分析发现,调控烟碱生物合成基因NtMYC2a、NtMYC2b、NtPMT和NtQPT的相对表达量出现不同程度增加,使NK-9株系在现蕾期、打顶期和成熟期的烟叶烟碱含量分别显著增加33.26%、36.41%和39.28%。这与Zhang 等[12]和Shoji 等[30]的研究结果一致,这些结果证明,NtJAZ1基因是烟碱生物合成调控的重要靶点之一。
在提高烟碱含量的同时,保证烟株正常的生长发育对于烟草品种定向改良至关重要。本研究中对NK-9株系的农艺性状进行分析,发现NtJAZ1基因失去功能后,基本不影响烟株正常的生长发育。虽然叶长与对照相比有所减小,但株高、叶数和叶宽显著提高,对烟叶生产并无严重的不良影响。荆叶醒等[31]在小麦研究中也发现,小麦TaJAZ1基因编辑后,植株生长发育状况良好。由此推测,NtJAZ1基因在应答茉莉酸甲酯(MeJA)信号调控次生代谢的同时,其功能缺失并不影响植株正常的生长发育。当然,由于烟株发育受栽培和多种环境因素的影响,NtJAZ1基因对烟株发育的影响还需要多年多点的试验进一步验证。
NtJAZ1基因在调控烟碱合成的同时,对烟叶其他化学成分也有一定影响。本研究中对获得的NK-9 株系烤后烟叶进行分析发现,NK-9 烟叶中性致香物质总量提高,其中新植二烯和类胡萝卜素类降解产物明显增加,这对烟叶品质有积极影响。但NtJAZ1基因的敲除也导致了烟叶叶面腺毛,尤其是长柄分泌型腺毛密度的减小,使腺毛分泌物,尤其是类西柏烷二萜化合物含量降低,最终导致了烤后烟叶中西柏烷类香气物质茄酮含量减少,这对烟叶品质有不利影响。关于JAZ家族基因调控腺毛发育方面,在拟南芥[32]、番茄[33]和青蒿[34]中已有报道,但NtJAZ1基因调控烟草腺毛发育的分子机制还需进一步研究。另外,可通过多性状聚合途径,在NtJAZ1敲除的植株上对腺毛密度进行定向改良,在保证烟碱高积累的同时,促进腺毛发育和分泌物的积累,从而提高烟叶品质。
4 结论
利用CRISPR/Cas9 技术获得了NtJAZ1基因敲除的定向改良株系NK-9。田间试验表明,NK-9 株系生长发育良好,在现蕾期、打顶期和成熟期中部叶片烟碱含量分别提高33.26%、36.41%和39.28%,烤后烟叶烟碱含量提高44.91%,中性致香物质总量显著增加。这表明NtJAZ1基因敲除能显著增加烟叶烟碱含量,促进烟叶中性致香物质的积累,是进行烟碱含量定向改良的重要靶点。
