中国汉族人群SNPH 基因多态性与精神分裂症的关联研究
2022-08-16陆佳晶黄欣欣黄茹燕吕钦谕易正辉
巫 珺 陆佳晶 黄欣欣 黄茹燕 吕钦谕,△ 易正辉,
(1上海交通大学医学院附属精神卫生中心精神科 上海 200030;2复旦大学附属华山医院精神科 上海 200040;3上海市虹口区精神卫生中心精神科 上海 200083)
精神分裂症(schizophrenia,SZ)是一种复杂的精神疾病,据估计全球有2 100 万人患有SZ,给社会与个体带来了极大的负担[1]。SZ 临床症状复杂,以阳性症状、阴性症状、认知功能损害作为主要核心症状,并伴随有情感障碍、意志行为障碍等其他临床表现[2-3]。目前认为SZ 起病受到遗传、生物和环境因素的交互影响。其中SZ 的神经递质失调假说是目前最为重要的假说。而神经递质的失调涉及多种蛋白质的协调和动态的相互作用。其中,可溶性N-乙基马来酰亚胺敏感因子附着蛋白受体(soluble N-ethylmaleimide-sensitive factor attachment protein receptor,SNARE)蛋白的复合物起着核心作用[4-5]。SNARE 复合蛋白或其调节因子的变化能够影响囊泡的募集、对接、膜融合和再循环[6],进而影响SZ 患者关键脑区不同神经递质的突触后信号转导[7],影响神经递质的释放,最终参与SZ 的发生[8-9]。
伸展蛋白(syntaphilin,SNPH)是SNARE 蛋白的重要调节因子。SNPH 蛋白在大脑特异性表达,特别是影响突触可塑性的区域表达丰富。其表达启动于嗜铬细胞分裂细胞系(pheochromocytomaderived cell line,PC12)细胞的神经元分化的诱导,并分化为两个细胞亚群,分别以线粒体外膜和突触质膜为靶点[10]。已知SNARE 复合蛋白是由突触融合蛋白-1、突触小泡蛋白以及突触相关蛋白 25(synaptosomal associated protein 25,SNAP-25)相互作用而形成。SNPH 蛋白能够在膜融合阶段与SNAP-25 竞争性结合游离的突触融合蛋白-1,来抑制SNARE 复合体的形成。体外实验显示,对于培养基中的海马神经元,SNPH 蛋白的瞬时过表达显著减少了神经递质的释放,而将SNPH 蛋白引入突触前颈上神经节的神经元则可抑制突触传递。总之,SNPH 蛋白能够作为一个分子钳,抑制SNARE复合体的装配,来调节突触囊泡的胞排作用[11]。并且蛋白激酶A(protein kinase A,PKA)磷酸化可以作为SNPH 蛋白的“关闭”开关,通过环磷酸腺苷(cyclic adenosine monophosphate,cAMP)依赖的信号转导途径阻断其与突触融合蛋白-1 的结合,抑制SNPH 蛋白的功能,说明SNPH的调节功能具有动态平衡[12]。并且,SNPH 蛋白也与神经退行性变存在关联[13]。也有证据显示,SZ 患者血浆神经源外泌体提取液中线粒体的SNPH 蛋白显著高于健康对照组[14],提示SNPH 蛋白参与SZ 发生的途径具有多样性。
SNARE 复合体与多种精神疾病的发生存在关联,除SZ 外尚包括双相情感障碍、注意缺陷多动障碍等[15-16]。因此,作为SNARE 复合体的重要抑制因子,SNPH基因可作为SZ 的候选基因。目前尚无SNPH基因多态性是否与SZ 存在关联的相关报道。因此本研究假设SNPH基因多态性与SZ 发生存在关 联。 我 们 选 取SNPH基 因 的 5 个 位 点(rs3764714、 rs3795139、 rs3803947、 rs3803949、rs6134520)进行多态性检测,通过病例对照的关联分析来探讨SNPH基因多态性与中国汉族人群SZ发生的关联性,以进一步明确SNPH基因与SZ 多态性的关系。
资料和方法
病例组本研究共纳入SZ 患者389 例,来源于2015 年9 月至2018 年9 月在上海交通大学医院院附属精神卫生中心住院或门诊患者。入组标准:(1)符合《国际疾病分类》第10 版(ICD-10)SZ 的诊断标准;(2)年龄小于65 岁;(3)汉族人群;(4)患者本人及其监护人签署知情同意书。排除标准:(1)目前或既往符合ICD-10 中除SZ 以外的其他精神疾病患者;(2)具有明显的自杀、自残及危害他人倾向的患者;(3)有严重躯体疾病的患者;(4)存在物质及药物滥用者。
对照组本研究共纳入正常对照433 例,来源于广告招募的社区志愿者。入组标准:(1)年龄小于65 岁;(2)汉族;(3)签署知情同意书;(4)健康者与患者无血缘关系。排除标准:(1)有精神疾病及精神疾病家族史;(2)有严重躯体疾病史;(3)孕妇和哺乳期妇女。
本研究获上海市精神卫生中心伦理审查委员会批准(批件号:2017-19R)。
SNP 位点的选择SNPH基因数据从千人基因组 数 据 库(http://grch37.ensembl.org/index.html)中查到并下载,使用HaploView 软件以及SNP 在线网 站(http://gvs.gs.washington.edu/GVS150/)筛选 标 签SNP(tag SNPs),针 对SNPH基 因(NC_000020.11,chr1:1266292~1309327)及 基 因 上 游2 000 bp,以连锁不平衡系数(r2)>0.8,微小等位基因频率(minor allele frequency,MAF)>0.05 的原则进行挑选,共挑选出5 个标签SNPs,即rs3764714[chr20:1306461(GRCh38.p12)],rs3795139[chr20:1305289(GRCh38.p12)],rs3803947[chr20:1308322(GRCh38. p12)] ,rs3803949 [chr20:1308693(GRCh38. p12)] ,rs6134520 [chr20:1307579(GRCh38.p12)]。其中rs3795139 位于外显子区域。rs3764714、rs3803947、rs3803949、rs6134520 位于3’端UTR 区。
由于不同基因座位的各等位基因在人群中以一定的频率出现。在某一群体中,不同座位某两个等位基因出现在同一条染色体上的频率高于预期的随机频率的现象,称为连锁不平衡。应用HaploView 4.2 软 件 对SNPH基 因5 个SNP 位 点 进行连锁不平衡分析(linkage disequilibrium,LD),单倍型频率<3.0%将不被纳入后续分析。以D’值来衡量各位点之间的连锁不平衡程度,建议D’>0.8可以构成一个单倍型区块。分析结果显示:SNPH基因的rs3764714、rs3795139、rs3803947、rs3803949、rs6134520 5 个SNP 位点可以构成一个单倍型区块(图1)。

图1 5 个SNPs 的连锁不平衡分析结果Fig 1 Results of linkage disequilibrium analysis of 5 SNPs loci
DNA 提取抽取病例组及对照组人群外周静脉血5 mL,置于EDTA 抗凝管中抗凝,采用血液基因组DNA 提取试剂盒(上海莱枫生物科技有限公司)提取全血DNA 后,于-80 ℃冰箱中冷冻保存。
基因型检测应用TaqMan 荧光探针基因分型技术对rs3764714、rs3795139、rs3803947、rs3803949、rs6134520 位点的SNP 进行检测。TaqMan SNP Genotyping Assays 试 剂 盒 、TaqMan SNP Genotyping Mix 试剂盒及PCR 扩充应用7900 HT荧光实时定量PCR 仪购自美国ABI 公司。PCR 扩增反应体系为5 μL,包括2×Taqman Master Mix 试剂2.0 μL,40×Taqman Genotyping Assay 试 剂0.05 μL,DNA(15~20 ng/μL)2 μL,H2O 0.95 μL。PCR 反 应 条 件:95 ℃10 min 预 变 性;95 ℃15 s,60 ℃1 min,共进行50 个循环,每例样本均重复检测3 次。 采 用 SDS version 2.1 软 件 中 Allelic Discrimination 程序进行基因分型,通过检测不同等位基因FAM 和VIC 荧光强度来判断样本的基因型,并将结果保存。
统计学方法选择位点前,使用SPSS 26.0 对病例组和对照组一般人口统计学资料进行比较,其中计数资料采用χ2检验,计量资料应用独立样本t检验。并使用Haploview 统计软件进行两两位点之间的LD,用D’值来度量各个位点间的连锁不平衡程度。选择位点后,应用在线SHEsis 软件(http://analysis.bio-x.cn/SHEsisMain.htm)进行单个位点的关联分析(等位基因、基因型频率的比较)及位点之间的单倍型分析,通过计算比值比(odds ratio,OR)与95%CI 以了解等位基因和疾病的关联强度(OR>1 是危险因素;OR<1 是保护因素),并对各组间基因型频率分布进行H-W 平衡吻合度检验。采用SNPstats(https://www.snpstats.net/snpstats/start.htm)在线软件对不同位点的遗传模式进行分析。关联分析结果采用Bonferroni进行校正检验,即每个SNP 的P值乘以所分析的遗传标记数目,以校正后的P<0.05表示位点与疾病之间的关联有显著性。
应用Quanto 1.2.4 软件进行计算样本的统计效能,5 个SNP 中rs3764714 的微小等位基因频率(MAF=0.056)最小。假设OR=1.5,加性模式为参考,疾病一般人群患病率为1%,病例组389 例,对照组433 例,计算结果显示:统计效能为0.99。
其中,在线Shesis 软件主要具备3 种分析功能,首先可以进行单位点分析,以进行等位基因频率和基因型频率统计为主,其次可对位点进行两两D’和r2的计算,最后可以对单倍型进行分析。 而SNPstats 是一个从遗传流行病学的角度设计的网络应用程序,能够进行关联分析。Quanto 1.2.4 软件是一个计算效能或所需样本大小的程序,用于基因、环境因素、基因-环境(G×E)相互作用或基因-基因(G×G)相互作用的关联研究。使用各个软件时均按照软件要求进行数据编码。
结 果
一般人口学资料病例组共389 例,其中男性208 例(53.5%),女性181 例(46.5%);年龄13~65岁,平均(36.74±18.27)岁。正常对照组共入组433例,其中男性224 例(51.7%),女性209 例(48.3%);年龄17~64 岁,平均(36.22±13.00)岁。病例组与对照组性别(χ2=0.329,P=0.566)和年龄(t=0.473,P=0.636)差异均无统计学意义。
Hardy-Weinberg 平衡检验5 个位点(rs3764714、rs3795139、rs3803947、rs3803949、rs6134520)在病例组和对照组中均符合H-W 平衡(P>0.05),可以被纳入后续的分析。
SNPH基因5 个位点等位基因和基因型分布的比较结果表明,病例组(SZ 组)与对照组(HC 组)中 5 个 SNP (rs3764714、rs3795139、rs3803947、rs3803949、rs6134520)等位基因及基因型分布差异无统计学意义(表1)。
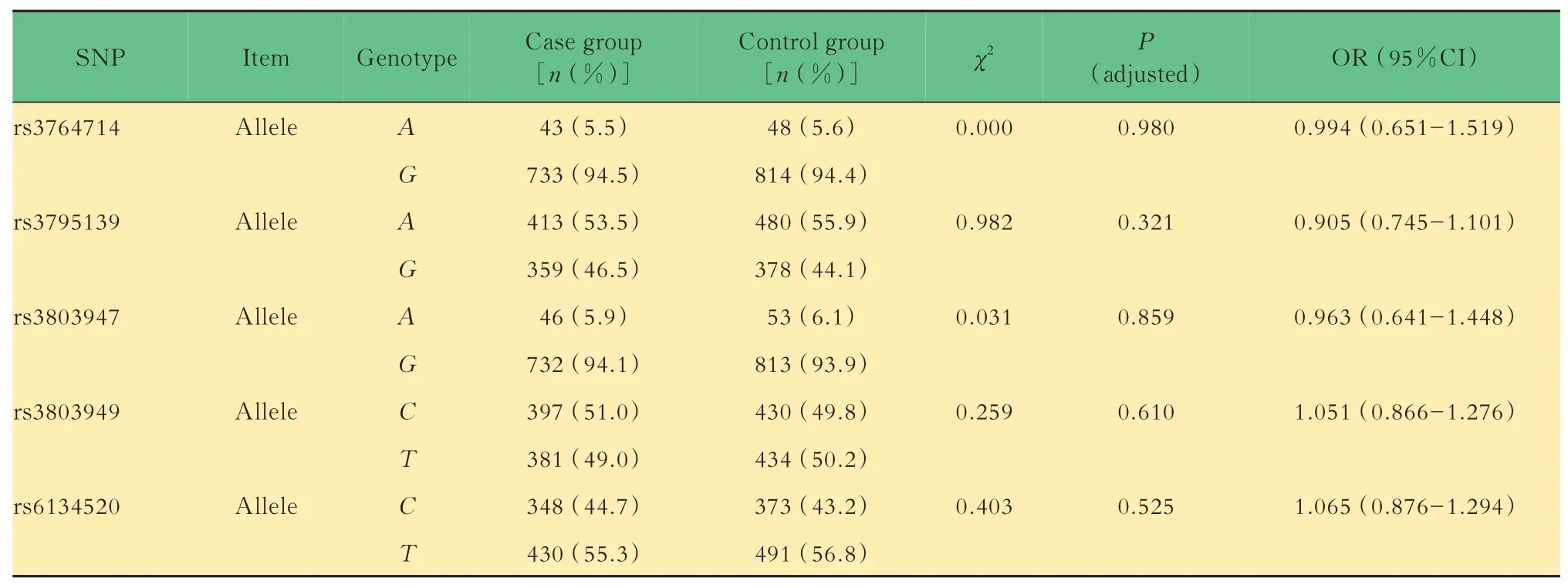
表1 病例组与对照组等位基因和基因型比较Tab 1 Comparison of allele frequencies and genotype frequencies of case group and control group
不同遗传模式下基因型分布比较在共显性、隐性、显性及加性遗传模式下,病例组与对照组rs3764714、 rs3795139、 rs3803947、 rs3803949、rs6134520 基因型分布差异均无统计学意义(表2,其中加性模式为参考)。

表2 病例组与对照组不同遗传模式下基因型的比较Tab 2 Comparison of genotype distribution in different models between case group and control group
病例组与对照组的单倍型分析应用SHEsis 在线软件对病例组(SZ 组)及对照组(HC 组)间单倍型进行 分 析,发 现 由rs3764714、rs3795139、rs3803947、rs3803949、rs6134520 位点组成的单倍型A-A-A-C-T,G-A-G-T-T,G-G-G-C-C频率>3%,后续分析发现3个单倍型在两组间的分布差异无统计学意义(表3)。
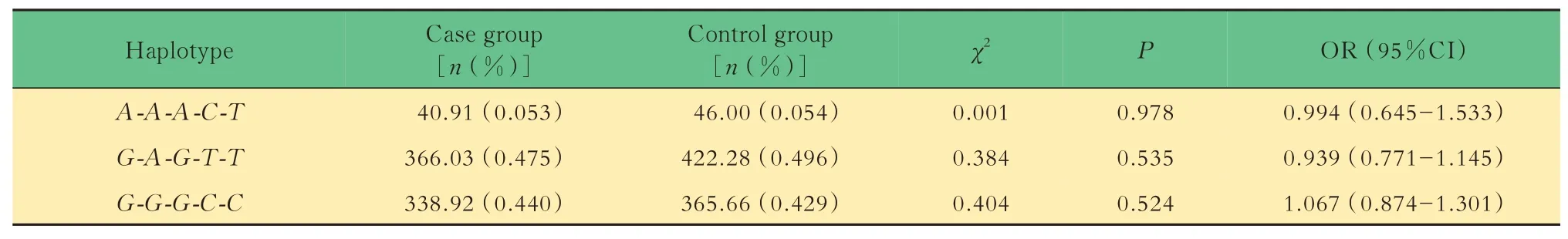
表3 病例组与对照组单倍型分布的比较(rs3764714,rs3795139,rs3803947,rs3803949,rs6134520)Tab 3 Comparison of Haplotype distribution of case group and control group(rs3764714,rs3795139,rs3803947,rs3803949,rs6134520)
讨 论
SZ 的神经递质假说是目前用以解释SZ 发病原因的主流观点,而SNARE 复合体对神经递质的释放过程起着关键作用,并最终影响SZ 的发生发展。因此,作为SNARE 复合体组装的抑制因子,SNPH蛋白同样在神经递质传递过程中扮演着重要的角色,SNPH基因可作为SZ 遗传学研究的候选基因之一而受到关注。
在本研究中,5 个位点在病例组和对照组中均符合H-W 平衡,可以进行下一步分析。而病例组和对照组在年龄与性别方面差异均无统计学差异,数据较为匹配。根据统计效能检验,本研究统计效能为0.99,统计效能较好。研究结果发现,在中国汉族人群SZ 患者和健康人群之间,SNPH基因5 个SNP位 点(rs3764714、rs3795139、rs3803947、rs3803949、rs6134520)等位基因及基因型频率分布差异均无统计学意义。在共显性、隐性及加性遗传模式下,两组之间5 个SNP 位点基因型频率分布差异均无统计学意义。这表明该基因的多态性可能与SZ 的发生没有关联,其特定的等位基因或者基因型可能不会导致SZ 的发病风险增加。针对SNPH基因5 个SNP 位点进行的连锁不平衡分析显示,该5 个位点可构成一个单倍型区块(D’>0.8)。然而,在单倍型 分 析 中,由rs3764714、rs3795139、rs3803947、rs3803949、rs6134520 位点组成的单倍型A-A-A-CT、G-A-G-T-T和G-G-G-C-C频率分布在病例组与对照组间差异无统计学意义(P=0.978、0.535、0.524),说明这3 种单倍型可能不会增加SZ 的发病风险。本研究结果显示,在中国汉族人群中SNPH基因可能不是SZ 发生的易感基因。
本研究选取的SNPH5 个SNP 位点在SZ 相关研究中均未见报道,但有研究显示,SNPH 蛋白与多种神经系统疾病相关。首先,SNPH 蛋白可能与脑白 质 病 变(white matter lesions,WMLs)有 关[17]。WMLs 是一种伴有脱髓鞘和认知功能下降的脑血管疾病。其次,体外实验证实,树突中SNPH过度表达会诱导N-甲基-D-天冬氨酸兴奋毒性、减少线粒体钙摄取和阻止线粒体自噬,导致线粒体功能的损害,最终与神经退行性疾病多发性硬化(multiple sclerosis,MS)相关[18]。可见SNPH 蛋白在大脑的结构与功能中具有一定的调节作用。但是对于同样伴随有认知功能下降的SZ,SNPH 蛋白作为其神经递质假说中的重要中介因素,相关的研究却极少,因此本研究具有一定的创新性和开拓性。
本研究在以下几个方面具有局限性。首先,所选5 个点属于一个区块,进行基因型分析时可能由于LD 的存在而造成混杂。候选基因法选点时应考虑编码SNPs(cSNPs),即在基因上首先考虑CDS区域(外显子)及启动子等,本研究选点时在编码SNPs(cSNPs)上考虑不足。多个位点位于3′UTR区,且仅1 个SNP 在外显子上,本研究对功能SNPs的探究不足。虽然3′-UTR 区可能在生物复杂性的调节中发挥重要作用,但更多3′-UTR 区的SNPs 高度保守,编码区的SNPs 功能普遍更为重要。其次,仅选取了SNPH基因的5 个SNP 位点进行关联分析,即使这5 个位点是经过筛选得到的,也不足以反映SNPH基因的全部信息,在后续研究中需扩大进行关联分析的SNP 位点规模,以进一步探索SNPH基因的相关信息。此外,本研究仅仅初步探讨了SNPH基因多态性与SZ 的相关性,该基因多态性与SZ 相关受体、递质、蛋白的关联,以及其他基因与SNPH之间的相互作用需要进一步研究,以明确SNPH基因多态性在SZ 发生机制中的作用。进一步验证SNPH基因多态性与SZ 的关联需要设计更加严谨的实验方案,增加样本量,保证入组患者的同质性,进行重复性实验。
作者贡献声明巫珺 论文构思、撰写和修订,病例收集,数据采集,基因分型。陆佳晶 数据统计和分析,论文撰写。黄欣欣,黄茹燕 病例收集,数据统计和分析。吕钦谕,易正辉 数据分析,论文构思和修订。
利益冲突声明所有作者均声明不存在利益冲突。
