饲粮能量水平对安格斯公牛生长性能、养分表观消化率和血清生化指标的影响
2022-08-09卢连强史长笑郝日康陈志明崔亚芳李英齐唐建鹏曹兵海苏华维
卢连强 史长笑 郝日康 陈志明 崔亚芳 李英齐 唐建鹏 曹兵海* 苏华维*
(1.中国农业大学动物科学技术学院,动物营养学国家重点实验室,北京 100193;2.商都县畜牧业服务中心,乌兰察布 013450)
安格斯牛是优秀的肉牛品种,具备耐粗饲、适应能力强、饲料报酬高、生长快以及肉品质好等特点[1]。美国、加拿大、澳大利亚和英国等国家普遍饲养安格斯阉牛,并利用阉牛肌内脂肪沉积能力强的特点,采用高精料育肥方式,生产带有大理石花纹的牛肉。
能量是肉牛生长发育的第一限制性营养因素。目前国内外已有不少关于安格斯牛饲粮能量水平的研究。Fortin等[2]通过自由采食和限制采食量(自由采食的65%~70%)的饲喂方式发现,低能量摄入组安格斯公牛、阉牛和母牛的皮下和肌间脂肪沉积率普遍较低。王秉龙等[3]研究发现,降低饲粮能量与粗蛋白质(CP)的比例(能蛋比),安格斯公牛平均日增重(ADG)和育肥效益显著提高,料重比显著降低(前期综合净能约为6.64 MJ/kg,CP水平分别为12.13%、11.47%和10.81%;后期综合净能约为6.94 MJ/kg,CP水平分别为11.63%、11.10%和10.63%)。Keady等[4]研究发现,提高饲粮精粗比(10∶90和77∶23)能提高安格斯杂交阉牛ADG和干物质采食量(DMI),进而提高饲料效率,降低料重比。郭延俭[5]研究发现,随着饲粮精粗比的提高(4∶6、6∶6和7∶3),安格斯杂交牛瘤胃pH逐渐下降,挥发性脂肪酸(VFA)浓度逐渐上升。Barber等[6]研究发现,提高饲粮能量水平 [代谢能(ME)为11.39和12.39 MJ/kg],安格斯阉牛胴体脂肪沉积增加,但ADG差异不显著。Schoonmaker等[7]研究安格斯杂交阉牛发现,饲喂相同谷物占比的饲粮,玉米组比大麦组ADG和DMI显著提高,料重比差异不显著;饲粮玉米青贮50%组(低能组)与15%组(高能组)相比,ADG差异不显著,DMI显著提高,料重比有升高趋势,经济效益显著下降。
通过以上文献可以发现,这些研究对象多为杂交牛、阉牛和母牛。而我国普遍饲养的是公牛,目前国内外还缺乏适合安格斯公牛饲养模式的饲粮能量水平推荐标准。因此,本研究探索了不同饲粮能量水平对安格斯公牛生长性能、养分表观消化率和血清生化指标的影响,为制定安格斯公牛的饲粮配方提供参考。
1 材料与方法
1.1 试验设计和饲养管理
试验选取(12.7±0.9)月龄和体重[(371.7±20.8) kg]相近的安格斯公牛36头,随机分成3个组,每组4个重复,每个重复3头牛(饲养在同一小栏)。3个组的公牛分别饲喂低(LE)、中(ME)和高(HE)3种能量水平的饲粮,饲粮参考NRC(2016)配制,3组饲粮CP水平同为13.5%,ME分别为10.28、10.88和11.49 MJ/kg。饲粮组成及营养水平见表1。试验预试期14 d,正试期120 d。
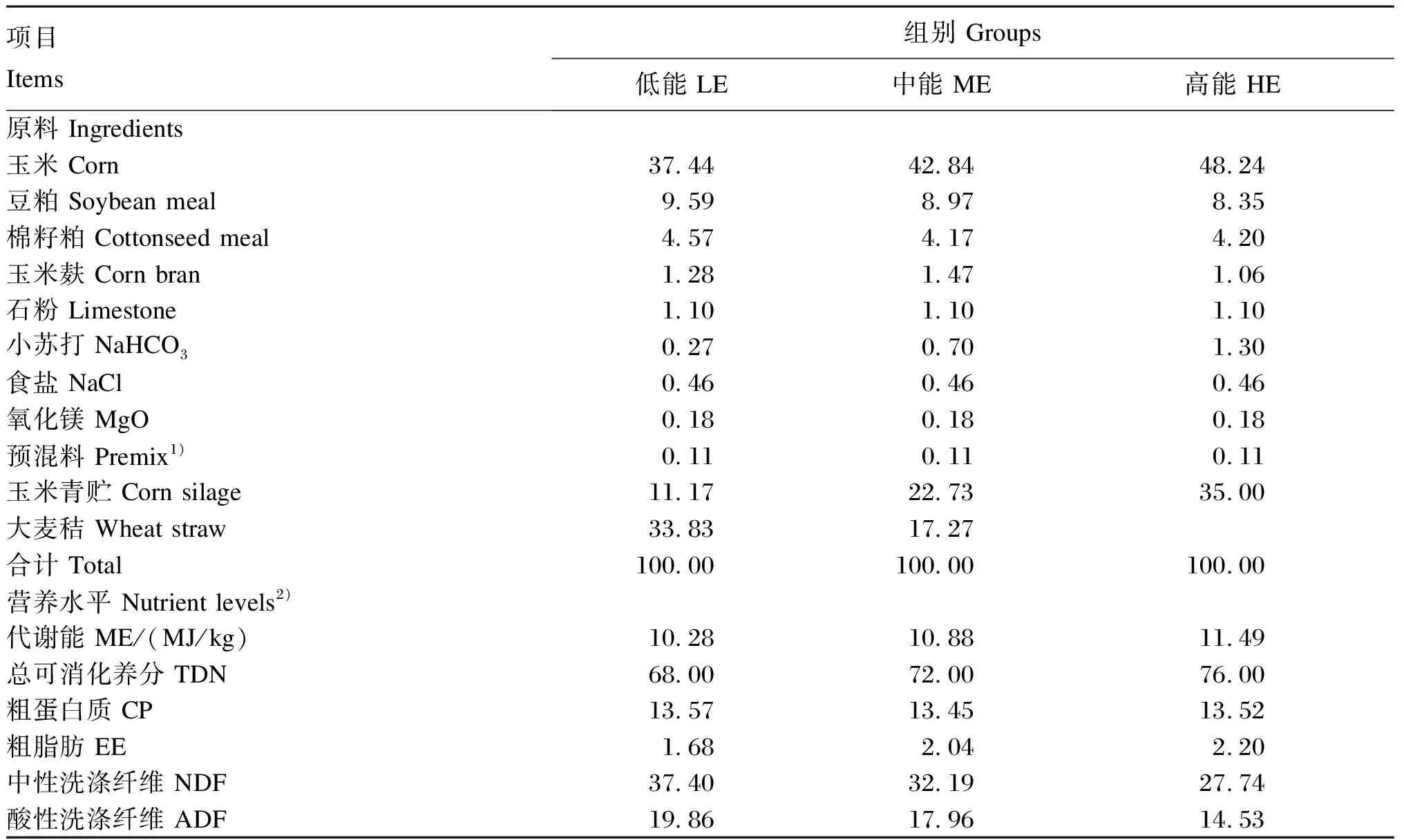
表1 饲粮组成及营养水平(干物质基础)
试验在内蒙古自治区乌兰察布市商都县六台滩肉牛养殖合作社进行。试验牛采用围栏散养,每天08:00和17:00各饲喂1次全混合日粮,自由采食和饮水。试验牛按照牛场常规管理进行免疫、驱虫等。
1.2 样品采集和指标测定
1.2.1 采食量和生长性能
试验期每天记录饲喂量和剩料量,用于计算DMI(kg/d)。正试期第1天和第120天称重,并计算ADG(kg/d)和料重比。
1.2.2 粪样采集和养分表观消化率测定
每月采集饲粮样本,于65 ℃箱内烘干至恒重,粉碎后过1 mm筛。根据AOAC所描述的方法[8]测定饲粮中干物质(DM)、CP、粗脂肪(EE)和粗灰分(Ash)含量,参照Van Soest等[9]所示的方法测定饲粮中中性洗涤纤维(NDF)和酸性洗涤纤维(ADF)含量。
粪样每2个月采集1次,每次连续3 d,每天3次。早、中、晚采集部分粪样,混合均匀后取样,并加入1/4粪重的10%(质量体积分数)酒石酸混匀,于65 ℃烘干,粉碎后过1 mm筛,保存待测。粪样中常规养分含量测定方法与饲粮样品相同。根据Van Keulen等[10]的方法,采用酸不溶性灰分法测定养分表观消化率。计算公式如下:
养分表观消化率=100×[1-(Ad×Nf)/(Af×Nd)]。
式中:Ad(g/kg)和Af(g/kg)分别为饲粮和粪样中酸不溶性灰分含量;Nd(g/kg)和Nf(g/kg)分别为饲粮和粪样中某养分含量。
1.2.3 瘤胃液采集和指标测定
每2个月采集1次瘤胃液,晨饲后2 h使用瘤胃导管收集瘤胃液200 mL左右,立即测定pH(205 Testo Instrument Co., Ltd.,德国),之后用4层纱布过滤。滤液分装于2个10 mL离心管中,并置于-20 ℃冷冻保存。其中一管用分光光度计,参考苯酚-次氯酸钠法[11]测定氨态氮(NH3-N)浓度;另一管用25%偏磷酸(4∶1,体积比)酸化后,根据Wang等[12]的方法采用气相色谱(GC-2014 Shimadzu Corporation,Kyoto,日本)测定瘤胃VFA浓度。
1.2.4 血液采集和指标测定
每2个月采集1次血液,晨饲前通过尾根静脉采血。将真空采血管(不含有抗凝剂)倾斜静置2 h后,在3 000 r/min转速下离心15 min,移取上层血清分装于1.5 mL离心管中,于-80 ℃冰箱中冷冻保存。血清葡萄糖(GLU)、总胆固醇(TC)、甘油三酯(TG)、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)、尿素氮(UREA)、β-羟丁酸(BHB)、总蛋白(TP)、白蛋白(ALB)和肌酐(CREA)含量以及谷丙转氨酶(ALT)、谷草转氨酶(AST)和碱性磷酸酶(ALP)活性等血清生化指标均采用全自动生化分析仪(HITACHI 7020,HITACHI Co.,Tokyo,日本),根据试剂盒(北京九强生物技术股份有限公司)的说明进行检测。血清活性氧(ROS)含量采用酶标仪(ST-360),根据试剂盒(上海科华生物工程股份有限公司)的说明进行检测。血清谷胱甘肽过氧化物酶(GSH-Px)、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活性和总抗氧化能力(T-AOC)以及丙二醛(MDA)含量等抗氧化指标均采用全自动生化分析仪(ZY KHB-1280,上海科华生物工程股份有限公司),根据试剂盒(南京建成生物工程研究所)的说明进行检测。血清氧化应激指数(OSI)根据以下公式计算:
OSI=ROS(U/mL)/T-AOC(U/mL)。
1.3 数据统计分析
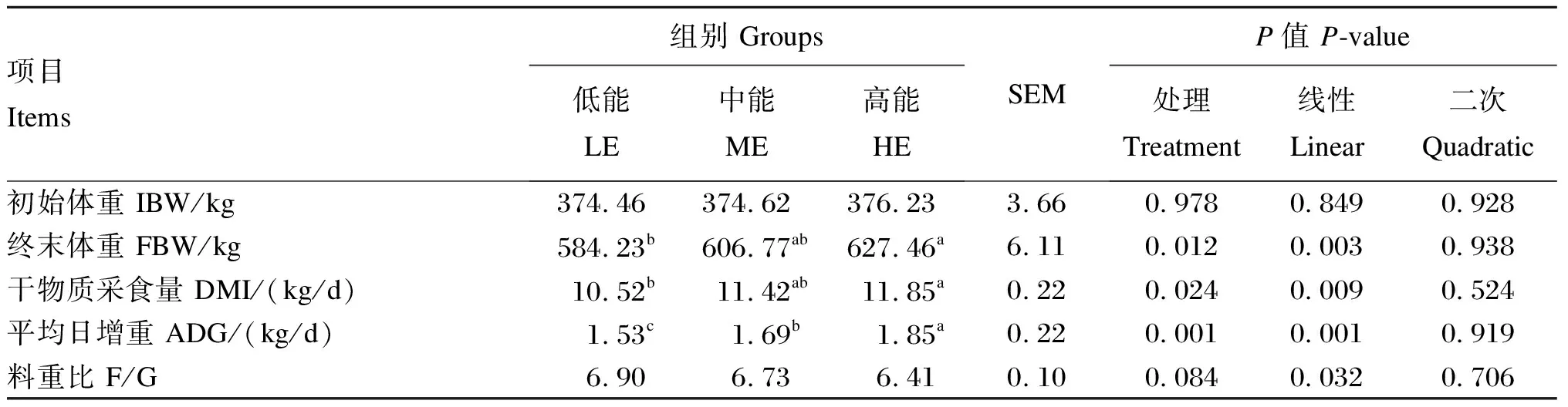
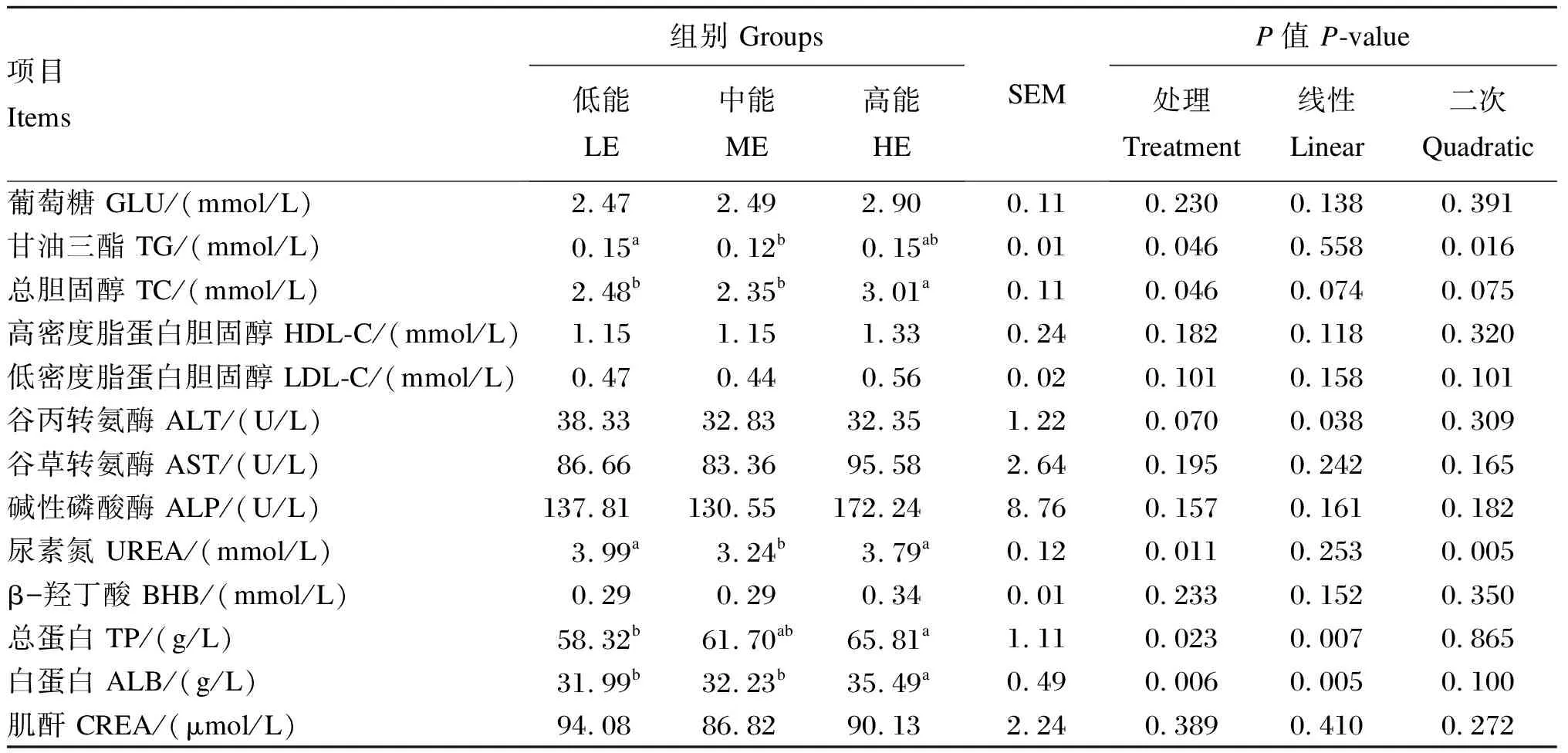
本试验所有数据均首先利用Excel 2016进行初步整理和分析,然后利用SPSS 22.0统计软件进行单因素方差分析(one-way ANOVA),差异显著时,用LSD法进行多重比较,以P<0.05表示差异显著,0.05 由表2可知,随着饲粮能量水平的提高,安格斯公牛终末体重、DMI和ADG均呈线性提高(P<0.05),料重比呈线性降低(P<0.05)。高能组安格斯公牛终末体重和DMI显著高于低能组(P<0.05)。 由表3可知,随着饲粮能量水平的提高,安格斯公牛对饲粮DM、有机物(OM)和ADF的表观消化率线性提高(P<0.05),对饲粮NDF的表观消化率呈线性提高的趋势(P=0.068),对饲粮EE的表观消化率呈二次变化(P<0.05)。其中,高能组DM表观消化率显著高于低能组(P<0.05),ADF表观消化率显著高于其他2组(P<0.05);中能组EE表观消化率显著高于其他2组(P<0.05)。 表2 饲粮能量水平对安格斯公牛生长性能的影响 表3 饲粮能量水平对安格斯公牛养分表观消化率的影响 由表4可知,随着饲粮能量水平的提高,安格斯公牛瘤胃液总挥发性脂肪酸(TVFA)、乙酸、丙酸、丁酸、戊酸和异戊酸浓度呈线性提高(P<0.05),瘤胃液乙酸/丙酸值和pH呈线性降低(P<0.05)。其中,高能组瘤胃液总挥发性脂肪酸(TVFA)、乙酸、丙酸、丁酸、戊酸和异戊酸浓度显著高于其他2组(P<0.05);低能组瘤胃液乙酸/丙酸值和pH显著高于其他2组(P<0.05)。 由表5可知,随着饲粮能量水平的提高,安格斯公牛血清TG和UREA含量呈二次变化(P<0.05),血清TP和ALB含量呈线性提高(P<0.05),血清ALT活性呈线性降低(P<0.05)。高能组血清TC和ALB含量显著高于其他2组(P<0.05)。 由表6可知,饲粮能量水平对安格斯公牛血清OSI和抗氧化指标均无显著影响(P>0.05)。 本试验结果发现,随着饲粮能量水平的提高,试验牛ADG和DMI显著提高,料重比线性降低,这与钟港等[13]使用湘南黄牛的研究结果相近(高能组和低能组精料ME分别为16.42和14.93 MJ/kg)。本试验中,安格斯公牛DMI升高的原因可能是随着饲粮能量水平的升高,NDF逐渐下降,这与Arelovich等[14]研究中饲粮NDF含量下降,而DMI提高的结果一致。随着DMI和饲粮能量水平的提高,肉牛摄入总养分增多,用于肌肉生长和脂肪沉积养分相对增加,用于维持消耗的养分相对减少,最终使肉牛ADG提高。 表4 饲粮能量水平对安格斯公牛瘤胃发酵参数的影响 表5 饲粮能量水平对安格斯公牛血清生化指标的影响 本试验中,随着饲粮能量水平的提高,料重比线性下降。这可能因为高能饲粮以丙酸发酵为主,甲烷能损失较少,低能饲粮以乙酸发酵为主,甲烷能损失较大[15]。此外,随着饲粮能量水平的提高,饲粮DM、OM和ADF表观消化率随之提高,这也与料重比降低有关。能蛋比是用来评价饲粮能量和蛋白质平衡的指标,影响动物生长潜能的发挥程度[16]。本试验饲粮CP水平一致,随着饲粮能量水平的提高,能蛋比提高更利于肉牛生长。 本试验中,随着饲粮能量水平的提高,饲粮DM和OM表观消化率线性提高,可能是由于高能组饲粮中非结构碳水化合物(NSC)含量较高,反刍动物更容易消化。本试验中,EE表观消化率随饲粮能量水平的提高先上升后下降,这与白大洋[17]研究中西门塔尔牛随着饲粮能量水平的提高,EE表观消化率先升高后下降的结果一致(前期、中期和后期综合净能分别为6.38、6.53和6.80 MJ/kg),具体原因有待进一步研究。 表6 饲粮能量水平对安格斯公牛血清OSI和抗氧化指标的影响 本试验中,随着饲粮能量水平的提高,饲粮ADF表观消化率线性提高,饲粮NDF表观消化率有线性提高趋势。原因可能是低能组结构性碳水化合物(SC)较多,难以进行消化,同时能量供给不足,用来消化降解养分的瘤胃微生物生长繁殖缓慢。此外,养分表观消化率还和饲粮能氮释放速率同步性有关。Chumpawadee等[18]研究发现,随着饲粮能氮释放速率同步指数的升高(0.39、0.50、0.62和0.74),婆罗门-泰国本土杂交阉牛的DM表观消化率呈提高趋势,OM和ADF表观消化率呈线性提高,而CP和NDF表观消化率差异不显著,这与本试验对应养分表观消化率的结果类似(1.0表示全天氮和能量供应的完全同步,而<1.0则表示异步程度)。这说明本试验中随着饲粮能量水平的提高,饲粮能氮释放速率同步性也随之增加。 瘤胃液的pH是瘤胃发酵过程的综合反映指标,受饲粮组成和能量水平影响较大。柏峻[19]报道,育肥前期锦江阉牛随饲粮能量水平的提高(综合净能分别为6.02、6.38、6.74、7.10和7.46 MJ/kg),瘤胃液pH呈逐渐降低的趋势。这可能是由于随着饲粮能量水平的提高,发酵迅速的可溶性碳水化合物含量更高,乳酸和VFA产生速率大于降解和吸收速率,同时NDF含量降低,反刍次数减少,唾液分泌量降低,综合导致pH下降。能量充足会促进瘤胃微生物对NH3-N的吸收利用,以合成瘤胃微生物蛋白(MCP),降低NH3-N浓度。相反,能量受限则会抑制微生物对NH3-N的摄取。本试验中,瘤胃液NH3-N浓度在适合瘤胃微生物生长的范围(6~30 mg/dL)之内[20],说明试验饲粮能量水平设置利于瘤胃微生物的发酵。 饲粮碳水化合物在瘤胃中发酵产生的VFA(乙酸、丙酸和丁酸等),是反刍动物主要能量来源,为瘤胃微生物提供生长繁殖的能量和碳架。由于瘤胃微生物的作用,SC发酵产生乙酸,NSC发酵产生丙酸。本试验中,随着饲粮能量水平的提高,瘤胃液乙酸和丙酸浓度线性提高,乙酸/丙酸值线性降低,这与DM、OM和ADF表观消化率随饲粮能量水平提高而提高的结果相对应。猜测是由于能量水平提高,饲粮NSC占比增加,导致丙酸产量上升,饲粮ADF含量减少,同时ADF消化率提高,综合造成乙酸浓度稍有增加,最终导致乙酸/丙酸值随饲粮能量水平提高而显著降低,也说明瘤胃发酵类型由乙酸发酵过渡到丙酸发酵。赵广永[21]认为谷物型饲粮在瘤胃中更容易被发酵,TVFA增多。本试验中,瘤胃液TVFA浓度随饲粮能量水平的提高而提高,可能是高能饲粮谷物占比更高导致。这与张娟利等[22]饲喂新疆褐牛杂交公牛高能饲粮组TVFA浓度显著高于低能量饲粮组的结果一致(高能组和对照组ME分别为13.1和10.0 MJ/kg)。本试验中,随着饲粮能量水平的提高,瘤胃液异戊酸和戊酸浓度显著升高,可能原因是瘤胃液中戊酸和异戊酸浓度会随着非纤维性碳水化合物/NDF的提高而显著提高[23]。异丁酸和异戊酸是支链脂肪酸,支链脂肪酸是瘤胃内纤维分解菌的生长因子,可提高纤维分解菌的数量促进纤维的消化[24]。因此,随着饲粮能量水平的提高,异戊酸浓度升高可能是ADF表观消化率提高的另一原因。 UREA是反映动物机体蛋白质代谢水平和饲粮氨基酸平衡状况的直观指标,其含量与机体内氮沉积有密切的关系。本试验中,中能组血清UREA含量显著低于低能组,可能是因为肉牛生长需要大量蛋白质,通过减少UREA的产生而增加氮的保留,从而有利于机体生长发育[25]。此外,本试验中,高能组血清UREA含量显著高于中能组,可能是因为高能组CP摄入量多,瘤胃降解蛋白超过了微生物的利用能力,从而造成瘤胃中NH3-N浓度以及血清中UREA含量升高。本试验中,瘤胃中NH3-N浓度与血清UREA含量变化趋势一致(低能组>高能组>中能组),更证实了我们的推测。 血清中TP主要包括ALB和球蛋白(GLB)2类。血清中TP含量能反映出饲粮中蛋白质的营养水平和动物对蛋白质的消化吸收程度,当体内蛋白质的合成作用增强时,血清中TP和ALB含量较高。本试验中,高能组血清TP含量显著高于低能组,血清ALB含量显著高于其他2组,原因可能是高能组饲粮蛋白质摄入量高且合成作用强,在生长性能上表现为高能组ADG显著高于其他组。与本试验结果不同,钟港等[13]认为高能组和低能组湘南黄牛血清TP和ALB含量差异不显著,可能原因是蛋白质摄入量无差异。ALT和AST活性是评价肝脏功能的重要指标,当动物肝细胞受到胁迫或损伤时活性会提高[25]。本试验中,饲粮能量水平提高对安格斯公牛血清AST和ALT活性无显著影响,说明适当提高饲粮能量水平不会对肉牛肝脏产生负面影响。 TC和TG是反映脂类代谢和吸收的综合指标,是血脂的重要组成成分,与摄入能量相关。本试验中,中能组血清TG含量显著低于低能组,有低于高能组的趋势,可能原因是高能组能量较高,代谢产物TG和TC较多。低能组能量较低,对血管中TG利用的能力较弱,所以血清TG含量较高[26]。本试验中,饲粮能量水平发生变化,而血清LDL-C、HDL-C、GLU和BHB含量没有变化,这说明适当提高饲粮能量水平并未造成机体能量代谢紊乱。 ROS是机体正常代谢产物,应激状态会导致ROS产生增多,导致脂质过氧化[27]。MDA是脂质过氧化反应的产物,能反映动物体内脂质过氧化程度和细胞损伤的程度,是氧化应激的标志[28]。机体的抗氧化体系由酶促和非酶促两大系统组成,非酶类抗氧化剂主要有维生素E、维生素C、谷胱甘肽及β-胡萝卜素等;酶类抗氧化剂有SOD、CAT及GSH-Px等[29]。T-AOC是反映机体抗氧化能力的综合指标,相比于单个抗氧化指标,能更好地反映机体抗氧化状态的整体水平[30]。氧化应激是由氧化剂和抗氧化剂之间的失衡所定义的,不仅要分析它们的水平,还要通过比例来分析它们间的关系[31]。因此,OSI常被作为反映氧化应激的综合指标[32]。本试验中,各组血清OSI和抗氧化指标均没有显著差异,表明饲粮能量水平提高并不会对安格斯公牛造成氧化应激。可能原因是安格斯牛能够快速适应高精料饲喂[33],饲粮营养水平合适和纤维充足。但是钟港等[13]发现,随着饲粮能量水平的提高,湘南黄牛血清中MDA含量和ALT活性显著上升,GSH-Px活性显著下降。这可能是由于长期饲喂高能饲料导致湘南黄牛体内沉积的TC含量较高,而肝脏代谢能力有限,造成肝脏损伤,引起氧化应激。 综上所述,饲喂高能量水平饲粮能提高安格斯公牛生长性能和养分表观消化率,降低料重比,促使瘤胃发酵向丙酸型转变,不会造成氧化应激;生产实践中,13~17月龄安格斯公牛推荐饲喂CP水平为13.5%、ME为11.49 MJ/kg的高能量水平饲粮。2 结 果
2.1 饲粮能量水平对安格斯公牛生长性能的影响
2.2 饲粮能量水平对安格斯公牛养分表观消化率的影响

2.3 饲粮能量水平对安格斯公牛瘤胃发酵参数的影响
2.4 饲粮能量水平对安格斯公牛血清生化指标的影响
2.5 饲粮能量水平对安格斯公牛血清OSI和抗氧化指标的影响
3 讨 论
3.1 饲粮能量水平对安格斯公牛生长性能的影响

3.2 饲粮能量水平对安格斯公牛养分表观消化率的影响

3.3 饲粮能量水平对安格斯公牛瘤胃发酵的影响
3.4 饲粮能量水平对安格斯公牛血清生化指标的影响
3.5 饲粮能量水平对安格斯公牛氧化应激和抗氧化能力的影响
4 结 论
