桂西北红椿(Toona ciliata)群落结构与更新特征1)
2022-08-04冯立新买凯乐杨善云苏付保伊洪伟李荣珍兰健花
冯立新 买凯乐 杨善云 苏付保 伊洪伟 李荣珍 兰健花
(广西生态工程职业技术学院,广西柳州,545004)
陈荣 朱昌叁
(玉林师范学院) (广西林业科学研究院)
群落的结构是指群落中各物种在时空上的配置状况,反映不同种群个体在群落中的组配情况及更新机制[1-4]。物种的自然更新能力是影响种群稳定的重要因素[5]。因此,人工经营、保护物种的策略[6-7],尤其对濒危种群落结构及种群更新特征的研究,可为区域濒危种群保护对策的制定提供理论依据[8-12]。
红椿(Toonaciliata)为国家二级保护濒危物种,为楝科香椿属,红椿材质有“中国桃花心木”之称,深受群众喜爱。我国天然红椿在云南、广西、贵州等地分布较为集中。红椿种群濒危的重要原因之一是天然更新困难。当前对红椿的研究主要集中在红椿的年龄结构及空间格局[13],红椿天然林种子和林分天然更新等方面[14],而少有对红椿群落结构和更新特征的研究。本文以广西西北部红椿(Toonaciliata)主要集中地群落为研究对象,分析红椿天然群落结构及天然更新方式,目的在于发现红椿群落的结构特征和红椿群落的更新特征以及更新过程中存在的问题,为红椿天然林群落的保护和更新提供技术支持。
1 研究地概况
选取桂西北具代表性的隆林、西林和田林县的红椿亚热带集中分布区为研究地。广西隆林县位于云贵高原东南,地形以中山为主,地貌结构有土山区(非溶岩)和石山区(溶岩)两大类,乔木主要有红椿(Toonaciliata)、银合欢(Leucaenaleucocephala)、粗糠柴(Mallotusphilippensis)、栓皮栎(Quercusvariabilis)、麻栎(Quercusacutissima)、枫香(Liquidambarformosana)、苦楝(Meliaazedarach)等,灌木主要有灰毛浆果楝(Cipadessacinerascens)、波叶山蚂蝗(Desmodiumsequax)、杜茎山(Maesajaponica)、马莲鞍(Streptocaulonjuventas)、毛鸡矢藤(Paederiafoetida)等,草本植物主要有野牡丹(Melastomacandidum)、千斤拔(Flemingiaprostrata)、散穗弓果黍(Cyrtococcumpatensvar.latifolium)、华南毛蕨(Cyclosorusparasiticus)、地桃花(Urenalobata)、蔓生莠竹(Microstegiumfasciculatum)、艾蒿(Artemisiaargyi)、碎米莎草(Cyperusiria)等;年均气温为19.1 ℃,年降水量为1144.6 mm。西林县位于桂西,地处为广西、云南、贵州三省(区)结合处,地形地貌主要以土山为主,乔木以红椿、华瓜木(Alangiumchinense)、三年桐(Verniciafordii)、灰毛浆果楝、苦楝、朴树(Celtissinensis)、阴香(Cinnamomumburmannii)、西南桦(Betulaalnoides)、木荷(Schimasuperba)、桉树(Eucalyptusrobusta)等为主,灌木主要有杜茎山、油茶(Camelliaoleifera)、盐肤木(Rhuschinensis)、红荷(Schimawallichii)、美丽胡枝子(Lespedezathunbergiisubsp.formosa)、野牡丹等,草本植被主要有蔓生秀竹、磨芋(魔芋)(Amorphophalluskonjac)、土淮山(Dioscoreajaponica)、毛鸡矢藤、紫茎泽兰(Ageratinaadenophora)、蕨菜(Pteridiumaquilinumvar.latiusculum)、华南毛蕨、地桃花等;年均气温为19.9 ℃,年降水量为743.5 mm。田林县位于桂西北,地处中亚热带地区,地形地貌以山地为主,由土山(砂岩与页岩)和石山两类组成;乔木以红椿、构树(Broussonetiapapyrifera)、华瓜木、板栗(Castaneamollissima)、杉木、八角(Illiciumverum)、秋枫(Bischofiajavanica)、泡桐(Paulowniafortunei)、大果榕(Ficusauriculata)、栓皮栎(Quercusvariabilis)等为主,灌木主要有白背叶(Mallotusapelta)、盐肤木、枇杷(Eriobotryajaponica)、华瓜木、美丽胡枝子、番石榴(Psidiumguajava)等,草本以地桃花、紫茎泽兰、蕨菜、土三七(Gynurajaponica)、东风菜(Asterscaber)、六棱菊(Laggeraalata)、蔓生秀竹、玉叶金花(Mussaendapubescens)、龙牙草(Agrimoniapilosa)、千斤拔等为主;年均温度17.0 ℃,年年降水量1 300.0 mm。
2 研究方法
2.1 样地设置与调查
在红椿主要分布地点设置调查样地。样地面积为20 m×20 m,每个县设置3个重复。每个样地分割成16个5 m×5 m的调查小样方,对小样方内植株高度大于1.3 m(H>1.3 m)的全部乔木、灌木和藤本进行每木调查,主要记录植物的名称、高度(藤本为长度)、胸径(或者地径);同时,在每个小样方中心设置面积为1 m×1 m的红椿幼苗及草本调查样方,调查红椿幼苗及草本,记录红椿幼苗的苗高和地径、草本的物种名称、高度、盖度、多度等信息。
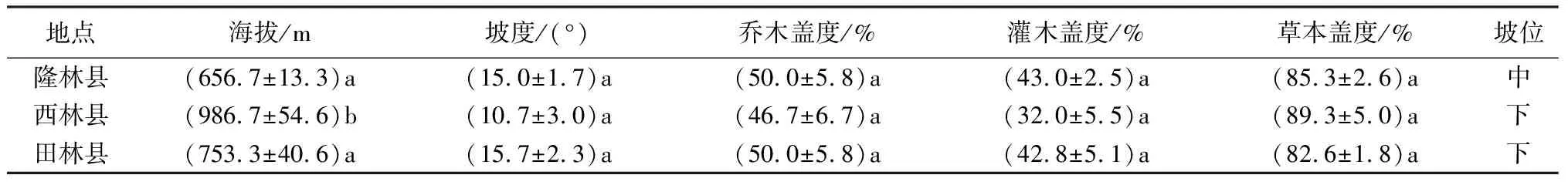
表1 研究地生境特征
2.2 数据统计分析
在野外调查数据的基础上,按乔、灌、藤,科、属、种等观测不同样地红椿群落组成情况,计算物种多度、丰富度、相对频度和相对显著度;以相对多度、相对频度和相对显著度的平均值计算物种重要值;以物种出现的样方数计算相对频度;以物种胸径计算相对显著度。对样地内的乔、灌、藤等以科、属、种的物种丰富度分别统计,并计算多样性指数(Simpson指数、Shannon-Wiener指数)。
根据调查观测,按上限排外法,径阶结构共划分为6级(D为胸高直径):I级(D<1 cm),II级(1 cm≤D<5 cm),III级(5 cm≤D<10 cm),IV级(10 cm≤D<20 cm),V级(20 cm≤D<40 cm);VI级(D≥40 cm)。按照径阶结构的6个等级,统计样地中不同径阶物种的丰富度及个体多度。
根据外业调查观测,按上限排外法,高度级结构共划分为4级(H为株高):I级(H<5 m),II级(5 m≤H<10 m),III级(10 m≤H<20 m),IV级(H≥20 m)。按照高度级结构的4个等级,统计样地内不同高度级物种的个体数量值(物种数量、丰富度、多度等)。
天然红椿群落一般以实生苗方式进行更新。根据对野外红椿样地中实生苗的调查数据,对3个样地点的红椿幼苗的多度、平均胸径、平均高度等进行统计。
利用SPSS 20.0对数据进行统计分析,利用单因素方差分析比较不同县之间物种多样性差异,采用Duncan多重比较检验显著性(P<0.05)。
3 结果与分析
3.1 红椿群落物种组成
在广西红椿群落中,共调查到物种99种,分属61科88属。其中乔木有42种,分属26科37属;灌木有11种,分属6科10属;藤本有5种,分属5科5属;草本有41种,分属24科36属。
隆林县红椿群落中,共发现物种52种,分属32科39属,其中,乔木有9种,灌木有16种,藤本有5种,草本有22种;西林县红椿群落中,共发现物种56种,分属41科50属,其中乔木有17种,灌木有17种,藤本有1种,草本有21种;田林县红椿群落中,共发现物种74种,分属50科67属,其中乔木有20种,灌木有22种,藤本有2种,草本有30种。
由表2可知,红椿群落3个调查地中,红椿的物种重要值均为最大值。隆林县物种重要值前3位的物种分别是红椿(0.627)、枫香(0.390)、散穗弓果黍(0.381);西林县则为红椿(0.605)、紫茎泽兰(0.478)、蔓生莠竹(0.406);田林为红椿(0.734)、紫茎泽兰(0.293)、千斤拔(0.285)。
3.2 红椿群落物种多样性特征
由表3可知,隆林县、西林县和田林县的科、属、物种、乔木、藤本、草本的物种丰富度无显著差异;隆林县和田林县的灌木的丰富度均高于西林县,且差异显著(P<0.05);3个县红椿群落的Shannon-Wiener多样性指数、Simpson多样性指数均无显著差异。
由图1可知,在物种相对多度曲线中,隆林县和田林县有明显的优势物种,隆林县明显优势物种为红椿和枫香;田林县明显优势物种为红椿、簕仔树;西林县则表现为相对优势物种为红椿、野牡丹和美丽胡枝子。3个地点群落内物种数量由多到少依次为:田林县、西林县、隆林县。3个地点红椿群落中均有大量低密度物种。
3.3 红椿群落的径阶结构
由表4可知,不同径阶物种丰富度上,西林县的红椿群落中的物种丰富度随着径阶的增大而逐渐降低;隆林县和田林县的红椿群落的物种丰富度在第II径阶最高,第II径阶之后逐渐降低;隆林县、西林县和田林县3个地点的物种丰富度在6个径阶的横向比较中均无显著差异。

表2 红椿群落优势种的重要值
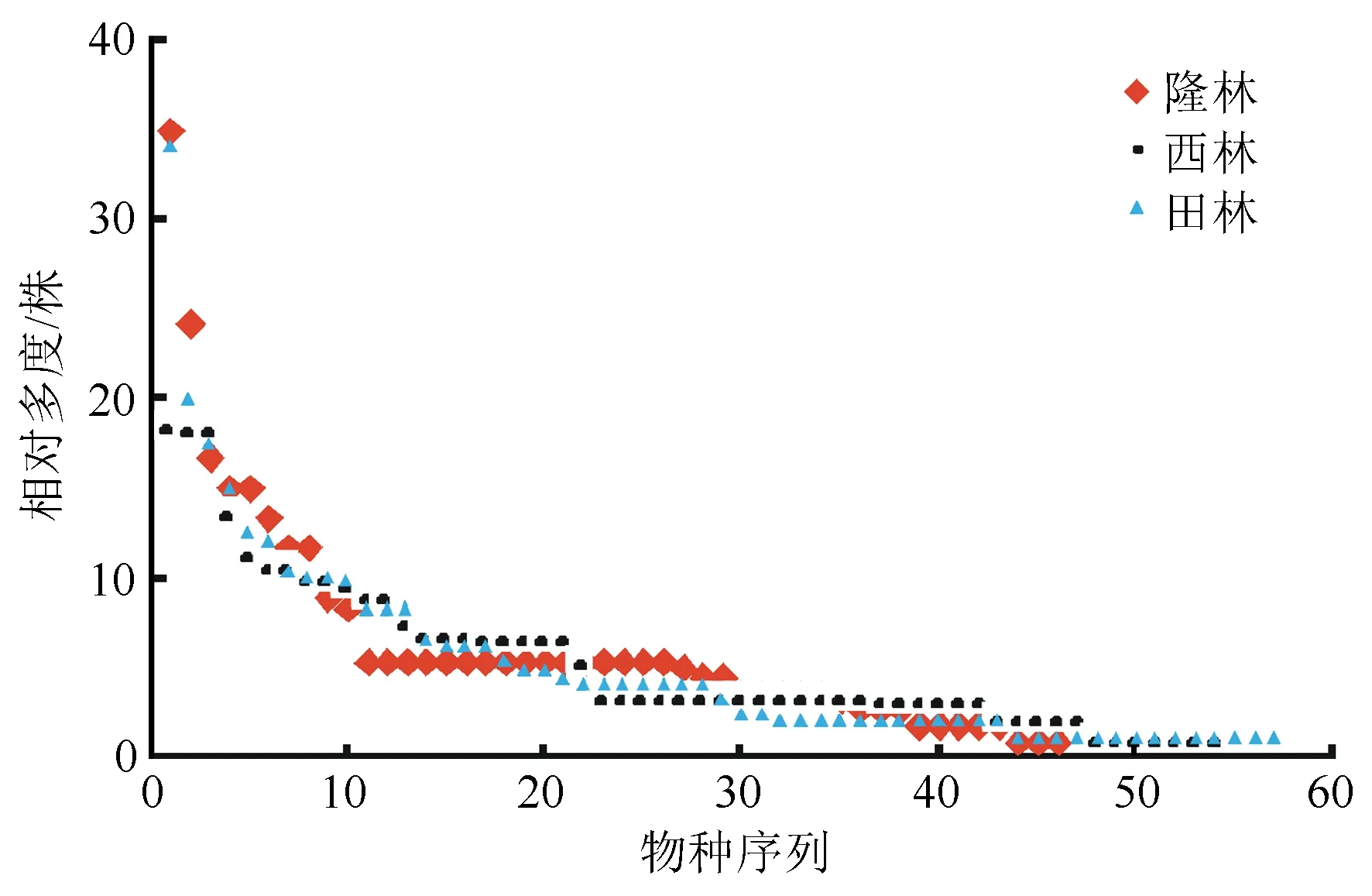
图1 红椿群落物种的相对多度
不同径阶多度上,隆林县、西林县和田林县在第II径阶物种多度最高;西林县红椿群落表现出随着径阶的增大多度逐渐降低的趋势,但隆林县和田林县从第II径阶多度逐渐降低,到第V径阶物种多度回升,到VI径阶物种多度又降低,为6个径阶中多度的最小值。在相同径阶中,第V径阶隆林县个体多度值最高,第VI径阶西林县物种个体多度值最高。

表3 红椿群落物种多样性

表4 红椿群落不同径阶的丰富度与多度

调查地红椿群落不同径阶的多度径阶Ⅰ径阶Ⅱ径阶Ⅲ径阶Ⅳ径阶Ⅴ径阶Ⅵ隆林县(37.33±7.42)a(68.00±27.23)a(25.33±17.49)a(13.33±1.33)a(53.33±17.94)b(1.33±1.33)a西林县(34.67±5.81)a(44.00±26.63)a(25.33±7.06)a(17.33±8.11)a(8.00±2.31)a(14.67±3.53)b田林县(45.33±22.43)a(78.67±47.12)a(13.33±2.67)a(6.67±2.67)a(34.67±7.06)ab(5.33±3.53)ab

表5 红椿群落不同高度级的丰富度与多度
由表5可知,不同高度级物种丰富度上,在桂西北调查的3个样地,红椿群落均表现出随着高度级的增大而物种丰富度逐渐降低的趋势。在相同高度级的比较中,3个地点物种丰富度均无显著差异。
不同高度级个体多度上,隆林县、西林县和田林县红椿群落均表现出随着高度级的增大而个体多度逐渐降低的趋势;在相同高度级的比较中,第IV高度级中隆林县物种个体多度最高,而在其他高度级中3个地点个体多度无显著差异。
3.4 种群更新特征
由表6可知,在红椿群落调查过程中,隆林县的3个样地未发现红椿幼苗。在西林县和田林县的个别样地发现红椿幼苗,但数量极少。田林县的3个样地的幼苗数量在3个调查点中最多,平均每个样地为3株幼苗,平均胸径和树高分别为1.3和0.73 m。西林县仅有1株幼苗,平均胸径为0.13 cm,平均树高为0.10 m。

表6 红椿实生幼苗特征
4 结论与讨论
4.1 红椿天然群落物种组成
本研究中桂西北红椿天然群落的主要特点为:群落的乔木上层为红椿所占据,不同地点红椿的伴生树种不同,以枫香、苦楝为主,个别地点伴生树种为银合欢、粗糠柴等,而灌木层的优势种主要为杜茎山、马莲鞍等。红椿群落3个样地的伴生乔木树种和灌木优势种均不相同,这与以往对红椿资源调查时的结果是相一致的(红椿与上层乔木的树种没有显著性关联)[15-16]。
天然植物群落中物种的组成受多种环境因素(例如温度、水分、养分等)的影响,而海拔又影响其他因子。在红椿群落中,物种组成与其所处海拔相关密切。我国西南地区红椿天然群落分布较多,在低山缓坡谷地或“四旁”生长较多,红椿天然分布的海拔范围为300~2 600 m[17]。本调查样地的海拔为650~1 000 m,属于云贵高原南麓,喀斯特峰丛洼地区。该地区土壤侵蚀退化严重,人地关系矛盾尖锐[18-19]。红椿经济价值高、人为因素对红椿的干扰以及自然环境变化等多种因素致使其天然更新困难。
调查地属于亚热带常绿落叶阔叶林(属于次生林),该地区植被恢复中被划分为比较高级的阶段,这也与以往对红椿资源调查的结果是一致的[20-21];桂西北的红椿群落具有乔、灌、藤、草等植物层次,与毛红椿的群落层次特征类似[15];桂西北调查地点的红椿群落中物种种类较多,多样性指数平均值为0.91。
4.2 红椿数量特征
红椿为群落的优势树种,重要值最大,为上层物种。红椿属于强阳喜光性树种,对生境要求较高。红椿幼苗多生长在光照好、地势相对平坦的地方,在红椿林冠下虽有少量幼苗,但不见幼树的情况较多[20]。本次调查中,红椿平均树高为18 m,平均胸径为28 cm,而幼苗数量很少,多数样地中幼树为0~3株,平均苗高为70 cm。红椿整体群落较小,为母树性质的红椿大树,红椿实生林地,林下苗木更新困难且分布不均匀。
红椿群落中第I高度级(树高小于5 m)的物种丰富度和多度具有最大值。红椿群落中第II径阶的物种丰富度和多度值最大,其次为I径阶。3个调查点红椿群落物种丰富,草本植物的平均盖度均大于82%,灌木的平均盖度为39%,且物种高度不超过5 m。而人工培育的1年生红椿苗平均苗高为40~120 cm[22-24],5年生红椿苗高平均为5.8 m[25],而红椿天然林下的幼苗苗高远小于人工培育的同龄幼苗。可见,红椿在幼苗和幼树阶段与群落其他物种及红椿物种间的竞争激烈,红椿幼苗稍耐阴,幼树及大树极喜阳,因此,红椿幼苗在幼苗阶段的成活及幼苗成长为幼树的环境竞争激烈,且在该地区的红椿群落中,速生物种多,不利于红椿成苗成长为幼树和大树。
4.3 红椿天然林保护
适宜的生境可促进森林更新[26],而一些地带性植被的建群种,如栎类林下“只见幼苗不见幼树”即是自然更新不良的体现[27]。本次调查到的红椿群落所在坡度均在10~15°的缓坡上,这样的缓坡能留存一定的土壤和水分,但抗外界人为干扰能力弱,影响群落的稳定和保护。其他因素(如光照、水分、林分郁闭度等)对红椿幼苗的分布和成活也有重要影响,但人为破坏是对桂西北红椿群落的主要影响因素,主要因为红椿的经济价值高,当地群众喜欢用红椿木材做家具和建房。
林地生境对红椿种子的萌发和生长具有重要影响,实生苗数量和母树的种子产量有关联性,光照对红椿结实有较大的影响,种子生长的生境对种子萌发、生长及开花结实也有重要的影响。红椿种子没有休眠期,种子散落后遇适当环境即萌发,未萌发的种子,其活力在3个月后就会失活,并且,遮阴和光照不足会降低种子的萌发率。红椿幼苗在林缘和四旁地等光照较好的地段具有较好的萌发率和生长趋势,但容易被误认为是杂灌草除去[20]。当年春季降水情况、林下枯枝落叶层厚度、结实情况、种群规模、人为干扰、林下光照量等因素直接影响红椿林下幼苗数量。
本次调查样地的红椿群落,林下红椿幼苗稀少,表现出更新难的特征。在调查地的红椿群落中,胸径小于5 cm的树种(径阶I和径阶II)占6个径阶中树种数量的50%以上。而胸径小于5 cm的红椿株数,在3个调查地点的样地中,仅有3株,并且分布不均匀。在调查的红椿群落中,高度小于5 m(第I高度级)的物种占群落总物种数量的65%~79%,而小于5 m(第I高度级)的红椿在调查地仅有2株。高龄级的红椿(通常具有较大的树高及胸径)呈聚集分布[13]。
此外,随着整个喀斯特地区脆弱生态系统压力的增大,人们对桂西北红椿资源过度采伐和开发,使红椿天然资源的生境遭到严重破坏,进而影响到红椿的种子产量,使红椿种子的萌发和生长更加困难。国家对濒危植物的保护措施虽然阻止了天然红椿小生境的恶化,但需要加强人工措施来促进红椿天然种群的天然更新。建议对红椿天然种群采取相应的就地保护措施[13],适当清除林下地被物,为红椿种子的萌发和生长提供空间和光照条件[14],并及时对飘落的种子进行覆土。在喀斯特脆弱生态系统中,应该人为保护或者创造林隙以促进林下幼苗更新,也可以在天然红椿林下补植红椿苗木,逐渐扩展红椿群落。同时,积极提倡和大力发展红椿人工林。
