基于RNA-Seq鉴定黑羽番鸭肉质风味差异的候选基因
2022-07-16段修军孙国波张蕾陆艳凤王丽华田培余刘嘉晟杨孟广
段修军 孙国波 张蕾 陆艳凤 王丽华 田培余 刘嘉晟 杨孟广




摘要: 為挖掘黑羽番鸭肉质风味相关候选基因,通过对黑羽番鸭胸肌、腿肌中挥发性风味物质进行测定,并通过Illumina HiSeq2500高通量测序进行转录组对比分析,结合参考基因组对所获得的序列进行序列比对、基因注释和差异表达等分析,筛选出差异表达基因并进行GO富集分析。通过荧光定量 PCR(Real-time PCR,qRT-PCR)方法检测4个差异候选基因表达水平,验证测序结果的可靠性。结果表明,黑羽番鸭胸肌中挥发性风味物质辛醛、2,3-辛二酮含量显著高于腿肌,3-羟基-2-丁酮含量显著低于腿肌( P < 0.05)。通过参考基因组比对和差异表达分析,初步获得614个差异表达基因,171个差异基因在黑羽番鸭腿肌组织上调表达,443个差异基因在黑羽番鸭腿肌组织下调表达。结合GO分析和KEGG富集分析,最终获得了20个候选功能基因,它们分别参与机体氨基酸形成、糖代谢以及脂肪代谢等生物过程,其中10个差异候选基因在黑羽番鸭胸肌中参与肌内脂肪代谢过程,这些基因可能通过形成肌内挥发性风味物质进而影响肌肉风味。qRT-PCR 验证结果表明,筛选出的差异候选基因表达趋势与转录组测序中表达的趋势相似,说明测序结果可靠。在黑羽番鸭胸肌、腿肌中挥发性风味物质对比基础上,结合RNA-Seq技术对黑羽番鸭肉质风味前体物质特异性形成机制进行综合分析,筛选出ACOT11、APOA1、FABP3等脂肪代谢相关的差异基因可能与黑羽番鸭胸、腿肌肉质风味形成相关。
关键词: 黑羽番鸭; 挥发性风味物质; 脂肪代谢; 转录组测序
中图分类号: S834+.89 文献标识码: A 文章编号: 1000-4440(2022)03-0739-09
Identification of candidate genes related to meat flavor in black Muscovy duck based on RNA-Seq
DUAN Xiu-jun, SUN Guo-bo, ZHANG Lei, LU Yan-feng, WANG Li-hua, TIAN Pei-yu, LIU Jia-sheng, YANG Meng-guang
(Jiangsu Agri-animal Husbandry Vocational College, Taizhou 225300, China)
Abstract: To explore the candidate genes related to meat flavor in black Muscovy duck, volatile flavor compounds in breast and leg muscles of black Muscovy duck were determined, and transcriptome analysis was performed by Illumina HiSeq2500 high-throughput sequencing. Based on reference genome, the obtained sequences were analyzed by sequence alignment, gene annotation and differential expression analysis, and differentially expressed genes were screened out and GO enrichment analysis was performed. Moreover, to verify the sequencing results, qRT-PCR was used to detect the expression levels of four differential candidate genes. Results showed that the contents of octanal and 2,3-octanedione in breast muscle were significantly higher than those in leg muscle, and the content of 3-hydroxy-2-butanone in breast muscle was significantly lower than that in leg muscle ( P < 0.05). Through genome comparison and differential expression analysis, 614 differentially expressed genes were preliminarily obtained, 171 differentially expressed genes were up-regulated and 443 differentially expressed genes were down-regulated in leg muscles of black Muscovy duck. By GO and KEGG enrichment analysis, 20 functional candidate genes were finally obtained. These candidate genes were involved in biological processes, such as amino acid formation, glucose metabolism and fat metabolism. Among them, ten differentially expressed candidate genes were involved in fat metabolism in breast muscle of black Muscovy duck, and these genes may affect muscle flavor by forming intramuscular volatile flavor compounds. Results of qRT-PCR verification showed that the expression trends of screened differentially expressed candidate genes were similar to those in transcriptome sequencing, indicating that the sequencing results were reliable. Based on the comparation of volatile flavor compounds in breast and leg muscles of black Muscovy duck, the specific formation mechanism of meat flavor precursors in black Muscovy duck was comprehensively analyzed, differentially expressed genes related to fat metabolism such as ACOT11, APOA1, FABP3 may be related to the formation of meat flavor in breast and leg muscles of black Muscovy duck.
Key words: black Muscovy duck; volatile flavor compounds; fat metabolism; transcriptome sequencing
番鸭( Cairna moschata )发源于南美洲和中美洲的热带,因其鸭嘴的基部和眼圈周围具有黑色或红色的肉瘤被称为瘤头鸭,又因其鸭肉瘦肉含量高,肉质鲜香,具有野禽风味的特点被称为麝香鸭 [1] 。番鸭按照羽色可以分为白羽、黑羽和花羽3种类群,其中黑羽番鸭因羽色和肉质特异性深受消费者喜爱,但是其体型较小,生长速度较慢 [2-3] 。为进一步提高黑羽番鸭的生产性能,江苏农牧科技职业学院自2000年开始,利用法国番鸭和福建黑羽番鸭进行选育,构建了羽色、胫、爪全黑的优质黑羽番鸭类群,其具有生长速度快、体型适中、脂肪含量低、肉质鲜美等优点 [4-5] 。
孙国波等 [3] 对公、母黑羽番鸭不同阶段胸、腿肌肉质进行对比分析,结果表明黑羽番鸭胆固醇含量较低,氨基酸丰富且含量高,公母鸭胸、腿肌营养成分存在一定差异性。王锦锋等 [6] 研究结果表明,黑羽番鸭胸肌的水分比腿肌高,腿肌粗蛋白质含量比胸肌高。吉文林等 [7] 对黑羽番鸭屠宰性能和肉质的研究结果表明,黑羽番鸭公鸭胸、腿肌的剪切力显著高于母鸭,公鸭肌肉中(胸、腿肌)pH 值显著高于母鸭(胸、腿肌)。林谦等 [8] 对不同羽色(黑羽、白羽以及黑白花羽)的公母番鸭进行屠宰性能测定对比分析,发现雄性黑羽番鸭的屠宰性能较好。钱建中等 [4] 研究结果表明,黑羽番鸭脂肪含量低于北京鸭、樱桃谷鸭,且黑羽番鸭公鸭胸肌剪切力和腿肌失水率显著高于母鸭。上述关于黑羽番鸭肉用性能的相关研究仅局限于个体肉质分析水平,对于黑羽番鸭胸、腿肌肉质性状的形成机制仍不清楚。
鸭肉的风味主要由鸭肉中的挥发性風味物质,即鸭肉中能产生挥发性肉香味的化合物成分,经加热后形成的特殊风味,是消费者对鸭肉食品产生的一系列生理反应和感官评定,是影响鸭肉食用口感的一个重要因素 [3,9] 。从基因组宏观层面对黑羽番鸭肉质性状的调控机制进行深度挖掘,可以挖掘黑羽番鸭肉质风味形成的关键基因。本研究以黑羽番鸭为研究对象,通过对黑羽番鸭胸肌、腿肌中挥发性风味物质进行测定,结合RNA-Seq转录组测序,对黑羽番鸭肉质风味前体物质特异性沉积机制进行系统、全面的综合分析,为培育优质肉用黑羽番鸭提供参考。
1 材料与方法
1.1 试验材料
本试验所用的黑羽番鸭由江苏农牧科技职业学院培育,选择具有相同遗传背景、同批孵化、同批出雏的黑羽番鸭,所用个体均在同一环境下饲养至300日龄。
1.2 样品采集
在黑羽番鸭大群体中随机抽样6只(公鸭3只,母鸭3只),按照国家实验动物处理行为准则进行屠宰,分别采取同一部位的胸肌、腿肌组织样本。将同一样本分为2组:一组迅速放入-80 ℃ 冰柜中速冻,以供测定分析挥发性风味物质含量;另一组放入-80 ℃ 冰箱保存,以供进行转录组测序。
1.3 试验方法
1.3.1 挥发性风味物质测定 参考王卫等 [10] 方法,利用气-质联用仪(Trace DSQ GC-MS)进行黑羽番鸭胸肌、腿肌中挥发性风味物质测定,并获得各成分相对含量。
1.3.2 RNA提取及转录组测序 通过Trizol法提取各肌肉样本组织总RNA,经RNA质检后进行转录组测序 [11] 。
1.3.3 测序数据处理及分析 结合测序获得的数据,对比参考基因组,获得黑羽番鸭胸、腿肌肉组织样本的特异转录组序列信息,完成基因组定位分析。在上述比对结果的基础上,使用htseq-count(v0.6.0)软件 [12] 统计每个转录本的表达水平,而后对所有基因进行对比分析,选取FDR(False discovery rate,错误发现率)小于0.05且差异倍数(FC)大于等于2的基因作为候选差异基因。最后通过GO(Gene ontology,基因本体)进行差异基因功能注释分析,以及KEGG(Kyoto encyclopedia of genes and genomes, 京都基因与基因组百科全书)进行通路富集分析 [13] 。
1.3.4 qRT-PCR检测 随机抽取方法1.3.2中差异表达基因4个,以反转录得到的黑羽番鸭胸、腿肌肉组织(样本号1~ 6)的cDNA为模板进行qRT-PCR(表1)检测,检验上述基因在黑羽番鸭胸、腿肌肉组织的表达规律与测序分析结果是否一致。
1.4 数据分析
qRT-PCR结果分析采用 2 -△△ Ct 方法,SPASS 20.0软件对试验所有数据进行 t 检验,以 P< 0.05视为差异显著, P< 0.01视为差异极显著 [14] 。
2 结果与分析
2.1 黑羽番鸭胸肌、腿肌中挥发性风味物质含量
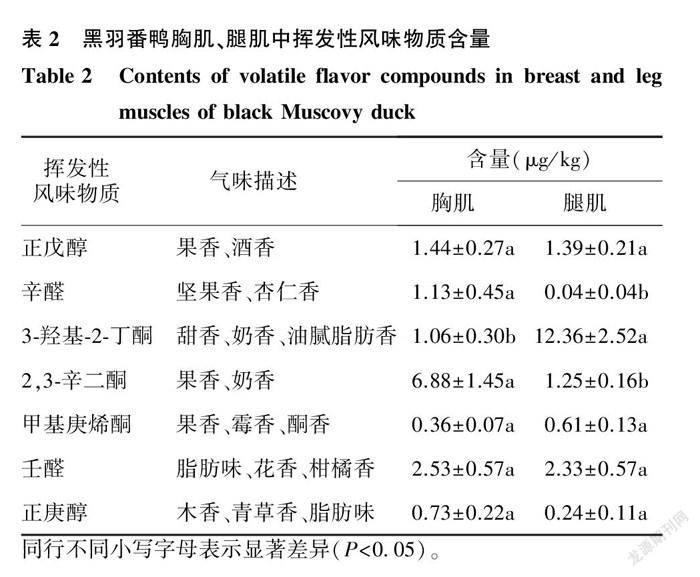
对黑羽番鸭胸肌、腿肌中的挥发性风味物质进行分离鉴定,通过NIST数据库进行检索,而后结合参考文献[15],筛选出可能与黑羽番鸭肉质风味相关的7种挥发性风味物质。由表2可知,黑羽番鸭胸肌中辛醛、2,3-辛二酮含量显著高于腿肌,3-羟基-2-丁酮含量显著低于腿肌,而正戊醇、甲基庚烯酮、壬醛以及正庚醇在胸、腿肌中含量无显著差异。
2.2 黑羽番鸭胸肌、腿肌转录组测序分析
共构建300日龄公、母黑羽番鸭的胸肌、腿肌各3个组织样品的转录组文库,对测序初始文库进行质量控制筛选,去除带接头、无法确定碱基信息以及低质量的测序数据(reads),获得的筛选数据结果如表3所示。300日龄公、母黑羽番鸭的胸肌、腿肌12个样品测序后获得39 527 132~ 54 977 176 条初始数据,而后经去除接头以及低质量数据筛选,获得37 824 878~ 52 542 526 条有效数据。过滤后有效数据占原始数据的比例大于94%,表明本试验构建的黑羽番鸭胸肌、腿肌12个样品文库质量较好。另外,过滤后有效数据中准确率在99.9%以上的碱基总数占93%以上,进一步说明本试验测序结果质量较好。综上所述,本试验构建的公、母黑羽番鸭的胸肌、腿肌12个组织转录组文库质量较好,可用于后续差异基因功能分析,保证了研究结果的可靠性。
2.3 黑羽番鸭腿肌与胸肌差异基因筛选及功能分析
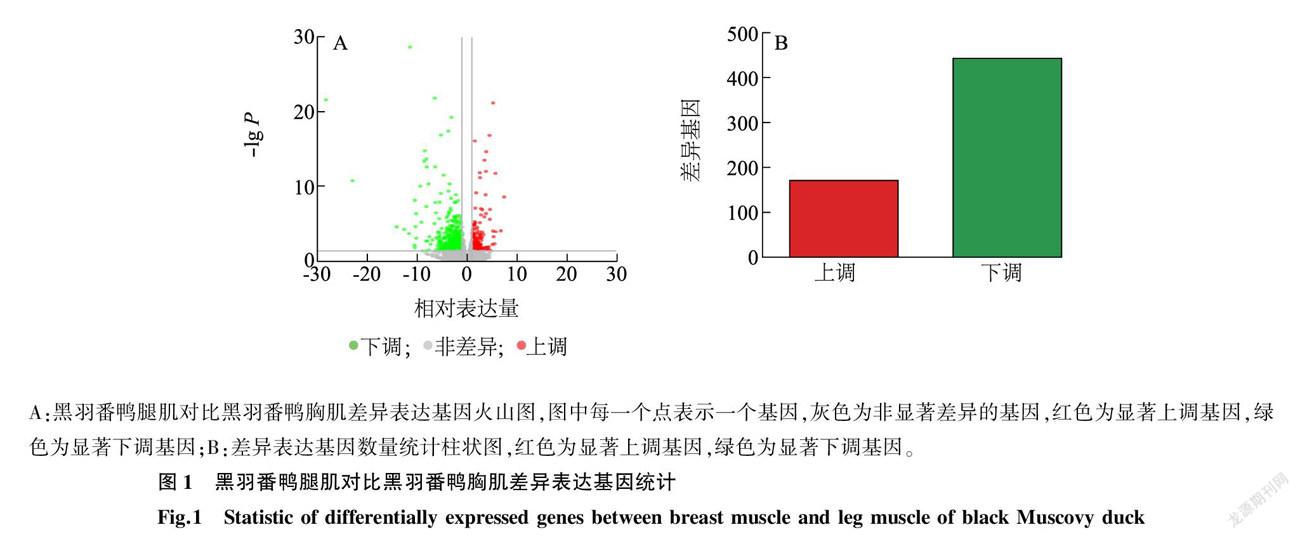
通过FPKM值 [9] 计算转录组测序所捕获的基因参考表达值,以黑羽番鸭腿肌组织为对照,黑羽番鸭胸肌组织为试验组,计算所捕获基因的差异倍数(Fold change值)。为聚焦关键基因,筛选黑羽番鸭腿肌对比黑羽番鸭胸肌基因表达差异值|log2 Fold change |≥1( P < 0.05)的差异基因作为黑羽番鸭胸肌、腿肌组织的候选差异基因,而后根据差异基因的 Fold change 对数值以及统计学显著程度绘制候选差异基因的火山图(图1),以标示黑羽番鸭胸肌、腿肌组织差异基因的分布情况。最终筛选出614个差异基因,其中171个差异基因上调表达(黑羽番鸭腿肌对比黑羽番鸭胸肌),443个差异基因下调表达(黑羽番鸭腿肌对比黑羽番鸭胸肌)。
2.4 黑羽番鸭腿肌与胸肌差异基因GO功能富集分析
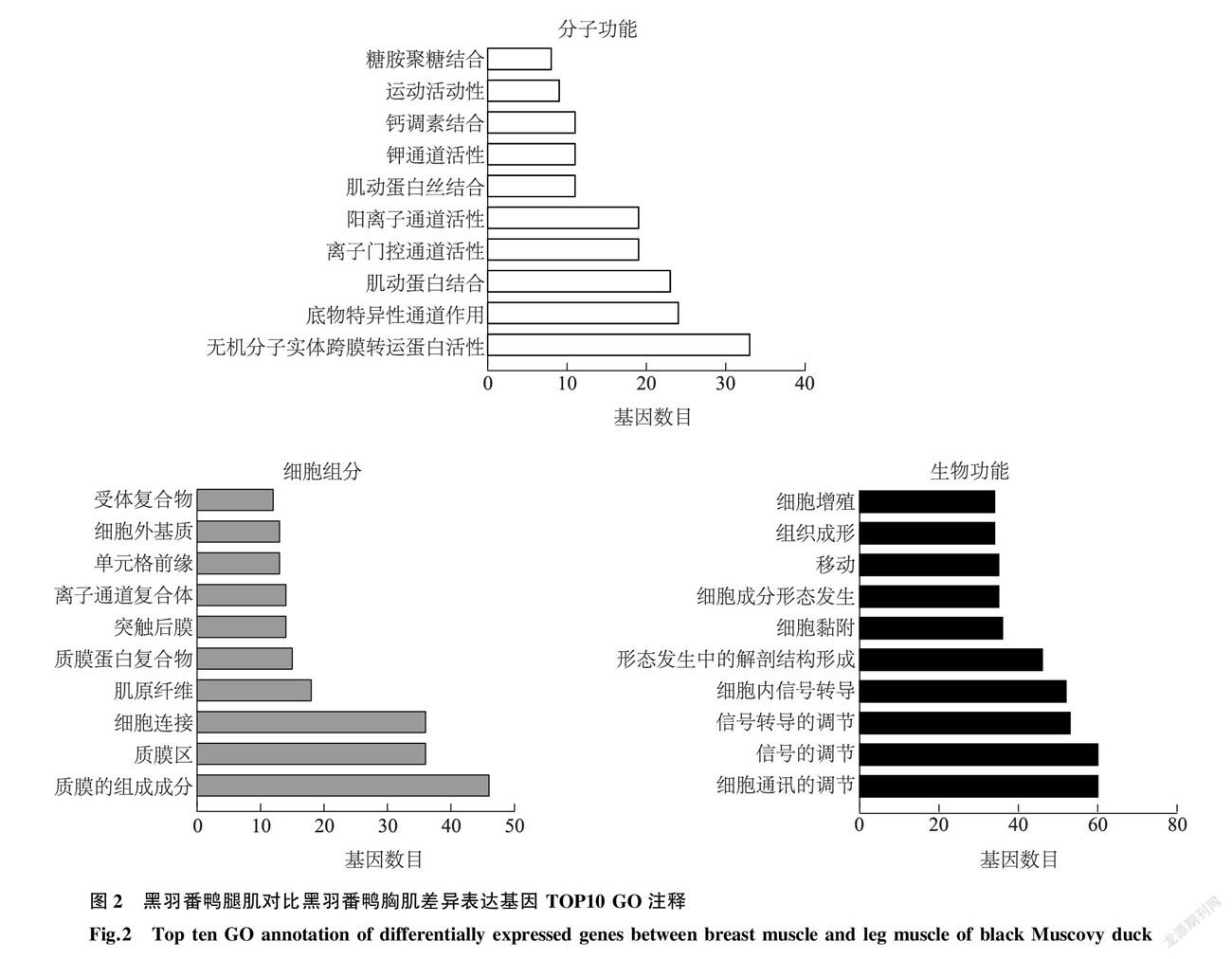
为探究黑羽番鸭腿肌与黑羽番鸭胸肌差异基因的功能,结合GO数据库对上述筛选获得的614个显著差异表达基因(|log2 Fold change |≥1( P < 0.05)进行基因功能富集分类,差异基因分别富集至分子功能(Molecular function,MF)、细胞组分(Cellular component,CC)以及生物过程(Biological process,BP)3个方面。为进一步聚焦614个显著差异基因在黑羽番鸭胸、腿肌肉组织风味物质形成中的作用,筛选在上述3类分组中前10的功能亚类(图2)。在MF功能中,黑羽番鸭胸、腿肌肉组织的差异表达基因在无机分子实体跨膜转运蛋白活性(Inorganic molecular entity transmembrane transporter activity)与底物特异性通道活性(Substrate-specific channel activity)中所占比例最大,分别聚集到33和24个差异表达基因。在BP功能中,细胞通讯的调控(Regulation of cell communication)与信号调节(Regulation of signaling)富集到的差异基因数目最多,各有60个差异基因。在CC功能中,差异基因在质膜组成成分(Integral component of plasma membrane)中所占的比例最高(46个差异基因),信号调节(Regulation of signaling)次之(36个差异基因)。
2.5 黑羽番鸭腿肌与胸肌差异基因KEGG通路注释
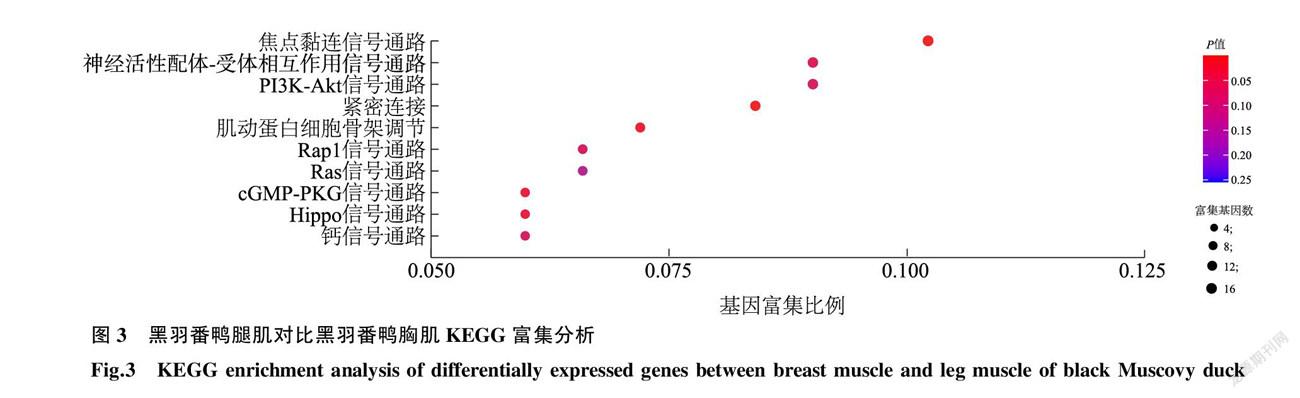
为深入解析差异基因在黑羽番鸭胸肌、腿肌风味物质形成中的作用,通过KEGG信号通路数据库对上述筛选获得的614个显著差异表达基因进行通路注释,共富集到109条差异表达的信号通路。以差异表达基因在此Pathway全部基因中所占的比例(GeneRatio)和 p 值进行筛选。GeneRatio越大,表示富集的程度越大; p 值越接近于0,表示富集越显著。由此筛选出富集最显著的10条差异信号通路(图3),包括焦点黏连(Focal adhesion)信号通路、PI3K-Akt(PI3K-Akt signaling pathway)信号通路以及Hippo(Hippo signaling pathway)信号通路等。
2.6 黑羽番鸭风味物质相关的差异表达基因筛选
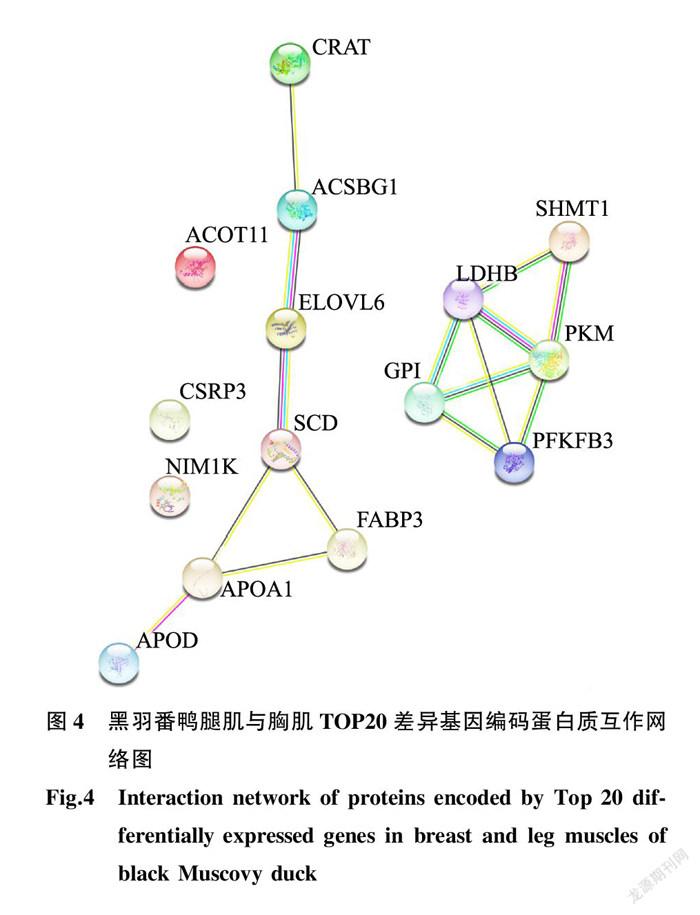
结合上述测序结果、差异基因筛选及功能分析,获得多个与风味物质形成有关的基因存在显著差异,表4列举出黑羽番鸭胸肌、腿肌转录组中与风味物质相关的差异表达前20个基因(TOP20)及其功能描述。根据GO分析以及KEGG功能注释,其中基因LOC101793529、LOC101794721、NIM1K和SHMT1与氨基酸形成有关,基因 GPI 、 LDHB 、PFKFB3、 PKM 参与糖代谢生物过程,剩余大部分差异基因与机体脂肪、脂肪酸代谢相关,例如基因ACOT11、ACSBG1、FABP3、ELOVL6和LOC101790670等。
为了进一步研究这些差异基因之间的互作关系,使用STRING(v.11.5)数据库构建蛋白质互作网络分析图。在黑羽番鸭胸肌、腿肌组织风味物质相关差异基因TOP20中,共鉴定出15种已知编码蛋白质,其中13种蛋白质被发现参与2组蛋白质相互作用(图4)。
2.7 黑羽番鸭腿肌与胸肌差异表达基因荧光定量验证
为验证黑羽番鸭胸、腿肌转录组差异基因测序结果,分别选择表4中黑羽番鸭腿肌高表达基因LOC101790670、PFKFB3和胸肌高表达基因LOC101792626、FABP3以 GADPH 为内参基因进行qRT-PCR 验证。qRT-PCR结果(图5)显示:上述4个差异基因在黑羽番鸭胸肌和腿肌组织中的上、下调表达规律与本试验通过黑羽番鸭胸肌和腿肌组织转录组测序获得的上、下调表达规律一致,证实本试验转录组测序获得的结果较为准确,所筛选出的差异基因可信,可用于后续的功能验证分析。
3 讨 论
黑羽番鸭生长速度快、体型适中,其鸭肉中瘦肉含量高、肉质鲜美,又因其具有独特的野禽风味而深受广大消费者喜爱 [1-2] 。本课题组前期研究结果表明,黑羽番鸭腿肌中胆固醇含量以及肌苷酸含量均低于胸肌 [3] 。而有关畜禽肉中风味物质研究结果表明,胆固醇、肌苷酸和挥发性风味物质等是形成其肉质风味的主要来源 [15] 。基于前期研究结果,本研究进一步对黑羽番鸭胸肌、腿肌中的挥发性风味物质进行对比分析,并结合RNA-Seq技术挖掘黑羽番鸭肉质风味的调控基因,为进一步培育优质肉用黑羽番鸭以及探究肉质风味调控机制提供参考。
禽肉中醛类、醇类及酮类含量不同可能是造成禽肉风味有差异的原因 [16] 。本试验对黑羽番鸭胸肌、腿肌中挥发性风味物质进行分离鉴定,筛选出可能与黑羽番鸭肉质风味相关的7种挥发性风味物质,其中辛醛、2,3-辛二酮含量在黑羽番鸭胸肌中显著高于腿肌,3-羟基-2-丁酮含量显著低于腿肌( P < 0.05),而正戊醇、甲基庚烯酮、壬醛以及正庚醇在胸肌和腿肌中含量无显著差异。辛醛由油酸氧化而形成,具有坚果香、杏仁香味,对肉香味的贡献较大,在黑羽番鸭胸肌中含量(1.13 μg/kg )显著高于腿肌(0.04 μg/kg )。2,3-辛二酮是脂质氧化的另一重要产物,在含量低时有果香、奶香味,黑羽番鸭胸肌中含量(6.88 μg/kg )显著高于腿肌(1.25 μg/kg ),说明胸肌中酮类物质含量较高,酮类物质又可结合脂质产生焦糖味 [17] ,从而有益于胸肌肉质的风味提升。3-羟基-2-丁酮是某些风味物质形成過程中的中间产物,具有甜香、奶香和油腻脂肪香味,黑羽番鸭胸肌中含量(1.06 μg/kg )显著低于腿肌(12.36 μg/kg ),但是由于其香味阈值高,需要在高含量下才能产生嗅觉反应 [15] ,因此对黑羽番鸭肉质风味的整体贡献不高。黑羽番鸭胸肌与腿肌挥发性风味物质对比后,筛选出辛醛、2,3-辛二酮以及3-羟基-2-丁酮这3类含量存在显著差异的物质,这可能与黑羽番鸭胸肌与腿肌肉质风味区别相关。
进一步对黑羽番鸭胸肌、腿肌进行转录组对比分析,筛选出614个差异基因,其中上调差异表达基因171个,下调差异表达基因443个。结合GO基因功能分析以及KEGG富集分析,筛选出黑羽番鸭胸肌、腿肌转录组中与风味物质相关的TOP20显著性差异基因。TOP20显著性差异基因参与机体脂肪代谢相关的差异基因占绝大多数:与胸肌相比,黑羽番鸭腿肌中大多数基因ACOT11、ACSBG1、APOA1、APOD、ELOVL6、FABP3和 SCD 等均呈现下调表达,少数基因 CRAT 、LOC101790670和LOC101792317等呈上調表达。酰基辅酶A硫酯酶11(ACOT11)作为酰基辅酶A硫酯酶( ACOT )家族成员之一,是一类催化脂肪酰基辅酶A水解形成游离脂肪酸和辅酶A的酶,在脂类的合成与分解过程中发挥着巨大的作用,能减少能量消耗同时保存体内热量 [18] 。ACOT11基因能促进小鼠体内游离脂肪酸浓度升高 [19] ,将该基因敲除后,小鼠脂肪酸氧化效率增加,脂质沉积减少 [20] ,可以抵抗饮食导致的肥胖以及肝脏脂肪变性 [21] 。脂肪酸结合蛋白3(Fatty acid-binding protein 3,FABP3)作为脂肪酸结合蛋白家族中的重要成员,可以通过长链脂肪酸的摄取、转运等调节体内脂肪和葡萄糖代谢过程,从而影响肌内脂肪沉积 [22-23] 。相关研究结果表明,FABP3的表达水平与30~ 90日龄的哈萨克绵羊肌肉组织中的脂肪含量呈显著正相关 [24] ,与拜城油鸡肌肉组织中的脂肪含量相关 [25] ,与6周龄北京鸭的腹脂率呈显著负相关关系 [26] ;在前体脂肪细胞中过表达可以有效促进脂肪合成 [27-28] 。
上调表达基因表达的蛋白质中,GPI(Glucose phosphate isomerase, 葡萄糖-6-磷酸异构酶)是一种多功能蛋白酶, 主要参与糖酵解和糖异生这2个生物过程,能够催化葡萄糖-6-磷酸和果糖-6-磷酸之间的可逆反应 [29] ,目前研究多集中于医学方面,GPI与非球型血红细胞贫血症、类风湿关节炎等疾病密切相关 [30-31] 。PFKFB3(6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3,6-磷酸果糖2-激酶/果糖-2,6-二磷酸酶3)参与生物体糖酵解反应,调节糖代谢通路和能量代谢过程 [32] ,研究证实,PFKFB3与肿瘤、血管钙化、糖尿病等疾病的发生相关 [33-36] 。其余上调表达基因如LOC101790670、LOC101792317等,从GO功能注释中可发现与脂肪代谢相关。但是,上述这些基因尚未有与肉质风味相关的研究报道。
肉质风味形成的主要原因之一是肌肉组织中含有大量的脂质,其风味的差异与肌内脂肪有着一定的关系。肌内脂肪作为风味物质的主要前体之一,在体内合成与分解的过程中形成大量的挥发性化合物,如酮类、醇类、酸类等,是挥发性风味物质的主要来源 [24] 。本试验发现黑羽番鸭胸肌中挥发性风味物质较腿肌中高,且通过转录组筛选获得大量参与机体脂肪代谢相关的差异基因。通过蛋白质网络互作图分析发现,这些差异基因的表达蛋白ACSBG1、APOA1、APOD、ELOVL6、FABP3、SCD和CRAT呈现出清晰的互作效应,这些基因可能共同作用于黑羽番鸭胸肌和腿肌的脂肪沉积过程,从而影响黑羽番鸭肉质风味,其具体的调控机制有待结合黑羽番鸭胸肌和腿肌的脂肪含量及各类脂肪酸比例进行深入研究。
4 结 论
本研究在黑羽番鸭胸肌、腿肌中挥发性风味物质对比基础上,结合RNA-Seq技术对黑羽番鸭肉质风味前体物质特异性沉积机制进行了系统、全面的综合分析,筛选出的ACOT11、APOA1、FABP3等脂肪代谢相关差异基因可能与黑羽番鸭肉质风味形成相关。本研究结果为深入探讨黑羽番鸭肉质风味形成机理以及优质肉用黑羽番鸭的选育提供了参考依据。
参考文献:
[1] 唐现文,张响英,陆艳凤. MC4R 在黑羽番鸭肌肉组织中的表达研究[J].中国家禽,2020,42(7):116-120.
[2] 孙国波,王 健,陆艳凤,等. 番鸭羽色与产肉性状的观测研究[J].江苏农业科学,2019,47(2):173-176.
[3] 孙国波,王日君,董 飚,等. 黑羽番鸭及继代选育后代的胸腿肌营养成分测定[J].贵州农业科学,2015,43(1):104-107.
[4] 钱建中,段修军,卞友庆,等. 不同性别黑羽番鸭屠宰性能、常规肉品质分析[J].江苏农业科学,2014,42(2):164-166.
[5] 董 飚,段修军,孙国波,等. GH 基因对黑羽番鸭体重和屠宰性能的遗传效应分析[J].浙江农业学报,2015,27(1):22-27.
[6] 王锦锋,包小成. 樱桃谷鸭、白羽番鸭、黑羽番鸭和苏牧鸭肉品质的比较研究[J].畜牧与兽医,2010,42(2):40-42.
[7] 吉文林,段修军,董 飚,等. 黑羽番鸭屠宰性能及肉品质的研究[J].西南农业学报,2013,26(2):795-797.
[8] 林 谦,吴买生,蒋桂韬,等. 不同羽色和性别番鸭屠宰性能及肌肉成分比较研究[J].家畜生态学报,2014,35(1):30-34.
[9] 章琳俐,李 丽,朱志明,等. 基于RNA-seq鉴定连城白鸭肉质风味相关候选基因[J].农业生物技术学报,2021,29(4):711-722.
[10] 王 卫,刘文龙,覃 瑶,等. 无抗放养模式有利于固始鸡挥发性风味物质的形成[J].湖南农业科学,2017(6):70-73.
[11] 张 蕾,章敬旗,章玮月,等. 泰州鹅开产前后肝脏组织转录组对比分析[J].福建农业学报,2020,35(9):987-996.
[12] ROBINSON M D, MCCARTHY D J, SMYTH G K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data[J]. Bioinformatics, 2010,26(1): 139-140.
[13] YU G C, WANG L G, HAN Y Y, et al. clusterProfiler: an R package for comparing biological themes among gene clusters[J]. Omics: a Journal of Integrative Biology,2012,16(5):284-287.
[14] 唐现文,张 蕾,章敬旗,等.高邮鸭卵巢中双黄蛋性状相关lncRNA表达差异分析[J].甘肃农业大学学报,2021,56(2):40-49,60.
[15] 孙旺斌,付 琪,薛瑞林,等. 不同枣粉水平对陕北白绒山羊肉挥发性风味物质的影响[J].动物营养学报,2021,33(10):1-13.
[16] 崔小燕,苟钟勇,蒋守群,等. 鸡肉风味的形成机制与调控研究进展[J].动物营养学报,2019,31(2):500-508.
[17] 李柯呈,徐宝才,姚 忠,等.干腌盐量对南京盐水鸭特征风味成分的影响[J].食品与发酵工业,2019,45(17):98-104.
[18] 张 丽,强 俊,徐 跑,等. 酰基辅酶A硫酯酶11基因(ACOT11)及其家族的研究进展[J].江苏农业科学,2021,49(2):12-18.
[19] ZHANG Y Z, LI Y X, MICHELE W N, et al. Targeted deletion of thioesterase superfamily member 1 promotes energy expenditure and protects against obesity and insulin resistance[J].Proceedings of the National Academy of Sciences of the United States of America,2012,109(14): 5417-5422.
[20] TAKAYUKI O, KANAKO I, RYOTA M, et al. Chronic high-fat feeding impairs adaptive induction of mitochondrial fatty acid combustion-associated proteins in brown adipose tissue of mice[J]. Biochemistry and Biophysics Reports,2017,10:32-38.
[21] 胡晓维,靳聪飞,刘新峰,等. 牛ACOT11基因克隆及其编码蛋白生物信息学分析[J].天津农学院学报,2015,22(3):1-6.
[22] 艾锦新,龙安炬,罗卫星,等. 黔北麻羊FABP3基因多态性及其与生长性状的相关性研究[J].畜牧与兽医,2021,53(1):13-18.
[23] BI Y L, YUAN X Y, ZHU P F, et al. A novel long noncoding RNA, ENSGALG00000021686, regulates the intracellular transport of fatty acids by targeting the FABP3 gene in chicken[J]. Biochemical and Biophysical Research Communications,2020,528(4):706-712.
[24] HUANG Z G, XIONG L, LIU Z S, et al. The develop-mental changes and effect on IMF content of H-FABP and PPAR-PPAR-γ mRNA expression on sheep muscle[J].Asia Cenetica Sinica,2006(6):507-514.
[25] 沙尔山别克·阿不地力大,舒 婷,吴旭升,等. 拜城油鸡 A-FABP、H-FABP和PPAR-PPAR-γ 基因mRNA表达的发育性变化及其与肌内脂肪含量的相关性研究[J].中国家禽,2019,41(14):5-11.
[26] 荣光辉,徐 琪,徐铁山,等. 脂肪型和瘦肉型北京鸭 A-FABP 基因组织表达特征[J].中国畜牧杂志,2015,51(1):5-9.
[27] YI BAO, WANG J G, WANG S, et al. Overexpression of Banna mini-pig inbred line fatty acid binding protein 3 promotes adipogenesis in 3T3-L1 preadipocytes.[J]. Cell Biology International, 2014, 38(8):918-923.
[28] 葛 凯. 脂肪细胞大小及脂肪代谢调控巢湖鸭肌内脂肪的分子机理[D].安徽农业大学,2019.
[29] 韩 龙,杜翠红.葡萄糖-6-磷酸异构酶研究进展[J].药物生物技术,2012,19(6):561-564.
[30] 白小娟,張丽卿.葡萄糖-6-磷酸异构酶在类风湿关节炎中的研究进展[J].山西医药杂志,2018,47(6):668-670.
[31] 黄 鹏,唐 黎,汪爱平,等. GPI 基因突变致葡萄糖磷酸异构酶缺乏症1例报告并文献复习[J].临床儿科杂志,2020,38(10):785-788.
[32] GRIESEL B A, MATSUZAKI S, BATUSHANSKY A, et al. PFKFB3-dependent glucose metabolism regulates 3T3-L1 adipocyte development[J]. The FASEB Journal, 2021, 35(7): 1-19.
[33] 李 骏,张 倩,江东根,等. PFKFB3基因在前列腺癌中的表达及其对前列腺癌细胞糖酵解及生长的影响[J].中华腔镜泌尿外科杂志(电子版),2020,14(6):460-465.
[34] 饶利栋,邓雪强,易 轩,等.PFKFB3通过调控PI3K/AKT信号通路促进骨肉瘤细胞的增殖及转移[J].肿瘤,2021,41(2):77-90.
[35] 牛 津,张 敏,王巧娟,等.PFKFB3抑制剂减轻高磷诱导的大鼠血管平滑肌细胞钙化[J].心脏杂志,2021,33(2):117-122.
[36] 袁 姿. 血清TLR4和PFKFB3水平与2型糖尿病及肥胖的相关性研究[D].苏州:苏州大学,2019.
(责任编辑:张震林)
收稿日期:2021-09-02
基金项目:泰州市科技支撑项目(TN201915);江苏省农业重大新品种创制项目(PZCZ201737)
作者简介:段修军(1972-),男,湖南南县人,硕士,教授,研究方向为动物遗传育种与繁殖。(E-mail)sgb1981@126.com
