新型小麦种衣剂JML对迟播冬小麦苗期生长和碳、氮代谢相关基因表达的影响
2022-07-16张小祥肖宁张新钵沈卫华季红娟李育红潘存红吉春明吴政吴云雨蔡跃刘建菊余玲陈梓春时薇张秀琴刘广青周长海姚友礼黄年生李爱宏
张小祥 肖宁 张新钵 沈卫华 季红娟 李育红 潘存红 吉春明 吴政 吴云雨 蔡跃 刘建菊 余玲 陈梓春 时薇 张秀琴 刘广青 周长海 姚友礼 黄年生 李爱宏




摘要: 為明确新型小麦种衣剂(JML)对迟播冬小麦幼苗生长及碳、氮代谢相关基因表达的影响,以扬麦23、扬麦24为试验材料,分析JML包衣处理后迟播冬小麦在种子活力、根系特征、干物质积累及碳、氮代谢相关基因表达等方面的变化特征。结果表明,JML处理后,扬麦23、扬麦24的发芽势和发芽率显著提高。茎叶与根器官中干物质积累增加,显著提高了植株氮积累量。根长、根表面积、根体积及根直径增加,与对照相比差异显著。JML处理提高了氮代谢中氮素吸收利用关键基因TaNRT1.1、TaNR、TaNIR、TaGS1、TaGS2等在幼苗植株中的表达水平,差异显著,但显著抑制了铵态氮转运蛋白基因TaAMT1.2的表达水平,促进了碳代谢中与光合作用密切相关的 TaClpP 、 TaRBC 、 TaPSB 基因的表达。因此,JML处理可以显著改善迟播冬小麦的种子活力,提高根系综合素质,增加苗期硝态氮的吸收转运、同化水平和叶片的碳同化能力,最终增加苗期植株干物质积累。
关键词: 冬小麦; 幼苗; 种子包衣剂; 氮、碳代谢; 基因表达
中图分类号: Q786 文献标识码: A 文章编号: 1000-4440(2022)03-0625-10
Response of the late sowing winter wheat seedling stage and carbon and nitrogen metabolism related gene expression to the new wheat seed coating agent JML
ZHANG Xiao-xiang 1 , XIAO Ning 1 , ZHANG Xin-bo 2 , SHEN Wei-hua 3 , JI Hong-juan 1 , LI Yu-hong 1 , PAN Cun-hong 1 , JI Chun-ming 1 , WU Zheng 1 , WU Yun-yu 1 , CAI Yue 1 , LIU Jian-ju 1 , YU Ling 1 , CHEN Zi-chun 1 , SHI Wei 1 , ZHANG Xiu-qin 1 , LIU Guang-qing 1 , ZHOU Chang-hai 1 , YAO You-li 2 , HUANG Nian-sheng 1 , LI Ai-hong 1
(1.Institute of Agricultural Sciences of the Lixiahe District in Jiangsu Province/Co-Innovation Center for Modern Production Technology of Grain Crops, Yangzhou 225007, China; 2.College of Agriculture, Yangzhou University/Jiangsu Provincial Key Laboratory of Crop Cultivation and Physiology, Yangzhou 225009, China; 3.Jiangsu Dazhong Farm Group Co., Ltd., Dafeng 224135, China)
Abstract: In order to clarify the effects of the new wheat seed coating agent (JML) on the growth of late sowing winter wheat seedlings and the expression of carbon and nitrogen metabolism-related genes, Yangmai23 and Yangmai24 were used as test materials to analyze the characteristics of seed vigor, root characteristics, dry matter accumulation, and expression of genes related to carbon and nitrogen metabolism of the late sowing winter wheat after JML treatment. The results showed that the germination vigor and germination rate of Yangmai23 and Yangmai24 were significantly improved after JML treatment. Dry matter accumulation in stems, leaves and root organs increased, which significantly increased plant nitrogen accumulation. Root length, root surface area, root volume and root diameter increased, which were significantly different compared with the control. JML treatment increased the expression levels of TaNRT1.1, TaNR, TaNIR, TaGS1, TaGS2 and other key genes for nitrogen obsorption and utilization in seedlings plants during nitrogen metabolism, and the difference was significant ( P <0.05). However, the expression of the ammonium nitrogen transporter gene TaAMT1.2 was significantly inhibited, and the expression of TaClpP , TaRBC and TaPSB genes closely related to photosynthesis in carbon metabolism was promoted. Therefore, JML treatment can significantly improve the seed vigor of late-sown winter wheat, improve the overall quality of the root system, increase the level of nitrate nitrogen absorption and assimilation at the seedling stage and the carbon assimilation capacity of the leaves, and ultimately increase the dry matter accumulation of the plant at the seedling stage.
Key words: winter wheat; seedling; seed coating agent; carbon and nitrogen metabolism; gene expression
稻麦两熟是长江中下游区域农作物的主要种植方式 [1] 。近年來,由于水稻品种生育期延迟、腾茬晚,造成稻茬小麦迟播、晚播,影响小麦适期播种,这已成为冬小麦获得稳产高产的主要障碍 [2-3] 。生产上为了保证群体质量,往往通过增加用种量及增施氮肥的手段来调节,有的地区小麦用种量已经达到350 kg/hm2 ,甚至更高 [4] ,氮肥施用量(纯N)则达到270 kg/hm2 [2] ,如此高的用种量与氮肥投入量造成生产成本过高,肥料利用率下降,农业面源污染加重 [5-6] 。另外,在不同地区由于土壤、栽培方式等因素,存在小麦种子出苗率和成苗率不高,苗弱不壮、病害重等生产问题,幼苗素质整体不高,难以达到壮苗入冬,这对小麦越冬及后期生长带来一系列不利影响,从而最终影响小麦的产量与品质。迟播冬小麦生产中,苗期如何培育健壮的个体,提高苗期群体质量,是目前稻茬冬小麦高产抗逆栽培研究的热点问题。有研究结果 [7] 表明,通过外源调节剂处理可以一定程度上缓解小麦迟播冻害问题,但对提高迟播小麦种子活力的问题还缺少有效的解决途径。为解决这一难题,除通过选择良种及增加用种量外,研发出既能提高种子出苗率和成苗率,又能增加氮素吸收利用的新型小麦种衣剂就显得极为迫切。
硝态氮与铵态氮是小麦根系吸收利用氮素的主要类型,氮同化是小麦吸收氮素的第一阶段,并直接关系到植株对氮素的转运及氨基酸等含氮化合物合成,同时氮素也作为信号调节着植株的生长与发育 [7-9] 。硝态氮在植株体内通过硝酸还原酶( NR )、亚硝酸还原酶( NiR )的作用还原为铵态盐,铵态盐继续在谷氨酸合成酶( GOGAT )、谷氨酞胺合成酶( GS )等主要功能酶作用与调控下参与各种氨基酸等含氮化合物的合成与转化,从而参与植株体内碳水化合物的形成 [10-14] 。但目前在专用种衣剂或外源生长调节剂作用下植株碳、氮代谢相关基因表达模式尚未见报道。
种衣剂是一种包裹于种子表面的肥料农药复合物,主要包括杀菌剂、杀虫剂、微量元素和植物生长调节剂等,对作物生长尤其苗期生长具有显著的正向促进效应,在水稻、玉米等作物上报道较多 [15-18] 。已有研究结果表明,尽管水稻的正向促进效应的大小随作物品种类型(基因型)不同有差异,但在不同栽培方式下均能够显著提高苗期植株的碳、氮吸收能力和植株可溶性蛋白质等含量,促进分蘖发生和干物质积累,为后期高产优质群体的构建奠定基础 [15] 。新型小麦种衣剂(JML)是江苏里下河地区农业科学研究所自主研制的小麦专用种衣剂,具有显著的壮苗、齐苗功能,但对苗期壮苗的相关影响及其内在的调节机制仍不清楚。本研究旨在分析新型小麦种衣剂(JML)在水稻秸秆全量还田下对迟播冬小麦苗期生长效应的影响,以期通过外源种衣剂来调控迟播冬小麦存在个体弱、群体质量不高等难题,对于缓解或解除小麦迟播造成的弱苗与氮素利用率低等问题具有重要意义,为迟播稻茬小麦高产、稳产与苗期抗逆栽培提供理论依据。
1 材料与方法
1.1 试验材料
试验于2017-2019年连续2年在江苏里下河地区农业科学研究所试验基地进行,前茬为水稻。试验地田块土质为沙壤土,土壤肥力中等,pH值7.51。耕作层0~ 20 cm,土壤有机质含量19.21 g/kg ,速效磷20.16 mg/kg ,速效钾62.37 mg/kg ,碱解氮89.27 mg/kg ,全氮1.56 g/kg ,全磷0.98 g/kg ,全钾12.18 g/kg 。供试品种为扬麦23(YM23,强筋型)、扬麦24(YM24,弱筋型),种衣剂为新型小麦专用种衣剂(JML),清水包衣为对照(CK)。
1.2 试验设计
该地区当年正常播期为10月25日,本试验播期为11月15日,迟播21 d,用种量127.5 kg/hm2 。以品种为主区,种衣剂处理为裂区。每品种分别进行JML包衣处理和清水包衣对照处理,每处理30 m 2 ,重复3次,随机区组排列。所有水稻秸秆全量还田,总施氮量(纯N)270 kg/hm2 ,其中基肥、追肥各50%,磷肥(P2 O5 )和钾肥(K2 O)各105 kg/hm2 ,磷肥和钾肥全部作基肥,其他管理措施同当地大田生产。包衣参照张小祥等 [15-16] 作物种子包衣的制作方法进行。
1.3 测定项目与测定方法
1.3.1 种子活力调查 参照国家标准农作物种子检验规程(GB/T3543.4-1995)的方法,并略有改进,进行小麦种子发芽试验 [19] 。在智能光温培养箱(型号:GZP250A)中用营养钵进行种子活力测试,营养钵高11.8 cm,外径14.4 cm,内径13.5 cm,内装水分饱和土壤1 200 g,每营养钵播正常小麦种子50粒,每处理播4钵,重复3次。设置常温和低温2种温度处理,常温处理:白天8∶00至 18∶00 温度控制在25 ℃,夜间18∶00至 第2 d 8∶00 温度控制在20 ℃;低温处理:白天8∶00 至18∶00 温度控制在20 ℃,夜间18∶00 至第2 d 8∶00 ,温度控制在15 ℃。测定种子发芽率( GP ),发芽势( GE )和活力指数( SVI ),其中 GP =7 d内总发芽粒数/总粒数×100%; GE =3 d内总发芽粒数/总粒数×100%; SVI = GP× S , S 为发芽结束后正常植株的干物质质量。
1.3.2 干物质质量与吸氮量测定 发芽后每隔7 d,连续28 d,每处理分别取60株长势一致的幼苗,分为地上部与根系两部分,烘箱中106 ℃杀青42 min,于85 ℃持续烘干至恒质量后分别称各部位干物质质量。植株干物质质量=地上部茎叶干物质质量+地下部根系干物质质量。
在第22 d和第30 d,分别取100株长势一致的幼苗,将植株杀青、烘干后研磨成粉末,每个样品取0.5 g置于消煮管中,使用浓硫酸定氮催化剂进行消煮,用全自动凯氏定氮仪(型号:FOSS Kijeltec 8400)测定植株茎、叶中含氮量。氮积累量=干物质质量× 含氮量。
1.3.3 根长、根平均直徑及根表面积测定 每处理在不同时期取60株长势一致的幼苗,采用全自动根系扫描分析仪进行扫描,然后用根系分析系统(北京澳作生态仪器有限公司产品)分析总根长、根体积、根系平均直径和根表面积。
1.3.4 不同器官总RNA的提取 在四叶一心期将小麦幼苗分为地上部和地下部,每个样本3次重复。在冰上快速分别取新鲜的叶片和根系,用锡箔纸在冰上进行包裹后,立即放入盛有液氮的泡沫盒中冷冻。在超净工作台采用Trizol试剂法提取叶片和根系中总RNA,通过Tiangen试剂盒[天根生化科技(北京)有限公司产品]将RNA反转录成单链cDNA。
1.3.5 引物的设计与合成 按照荧光实时定量qRT_PCR引物相关设计原则,利用Beacon designer 7.0 软件设计与目的基因相关的特异性引物。以小麦植株内 TaActin 基因的表达水平作为内参 [14] ,试验所用引物见表1。
1.3.6 实时荧光定量PCR(qRT-PCR) 参照TaKaRa SYBR Green II定量试剂盒的说明书进行qRT-PCR反应体系的配置,qRT-PCR采用CFX96 TM 实时荧光定量PCR仪(美国伯乐公司产品)进行。定量总反应体系为11 μl,其中引物为2 μl,模板cDNA为1 μl,MIX混合液为4 μl,去RNA酶水为4 μl,重复3次,样品处理重复3次。基因表达量计算方法:目的基因相对表达量=2 -△△ Ct ,其中,△△ Ct =样品基因△ Ct - 参比样品△ Ct ,样品基因△ Ct =内参基因 Ct - 目的基因 Ct ,参比样品△ Ct =参比样品内参基因 Ct - 参比样品目的基因 Ct 。
1.3.7 数据处理 2年数据变化规律一致,本研究选择2018-2019年度试验数据进行分析。试验数据采用Sigmaplot 10.0作图,采用SPSS Statistic 22软件进行方差分析,各处理的比较采用最小显著差数测验法(LSD)。
2 结果与分析
2.1 新型小麦种衣剂JML对小麦种子活力的影响
种子活力指数是种子发芽速率特征和具体生长量的综合反映,是作物种子萌发质量的重要指标。从表2中可以看出,新型小麦种衣剂JML处理后,常温下两品种间发芽趋势一致,发芽势、发芽率较空白对照(CK)平均提高11.67%和5.67%,差异显著( P < 0.05),活力指数较对照平均增幅26.98%,达显著水平。在低温处理下,两品种的发芽势、发芽率较CK平均提高5.50%和8.67%,差异显著( P < 0.05)。表明新型小麦种衣剂JML能够明显提高迟播冬小麦在常温和低温下的种子活力,增加出苗率,可减少用种量。
2.2 新型小麦种衣剂JML对小麦幼苗干物质积累的影响
苗期干物质积累的快慢在一定程度上反映了植株生长势的强弱,在小麦迟播条件下苗期干物质积累的快慢尤为重要。从图1中可以看出,两品种间变化趋势一致。在幼苗期JML处理过的幼苗茎叶及根器官干物质积累在第7 d、第14 d、第21 d及第28 d均超过同期对照,茎叶超过对照12.84%、14.43%、11.38%及12.94%,根超过对照12.75%、13.89%、15.83%和15.42%,差异均达极显著水平( P < 0.01)。表明JML有效促进了迟播小麦在幼苗阶段地上部、地下部干物质的积累,提高了植株苗期的综合素质,利于壮根壮苗。
2.3 新型小麦种衣剂JML对小麦幼苗氮素积累的影响
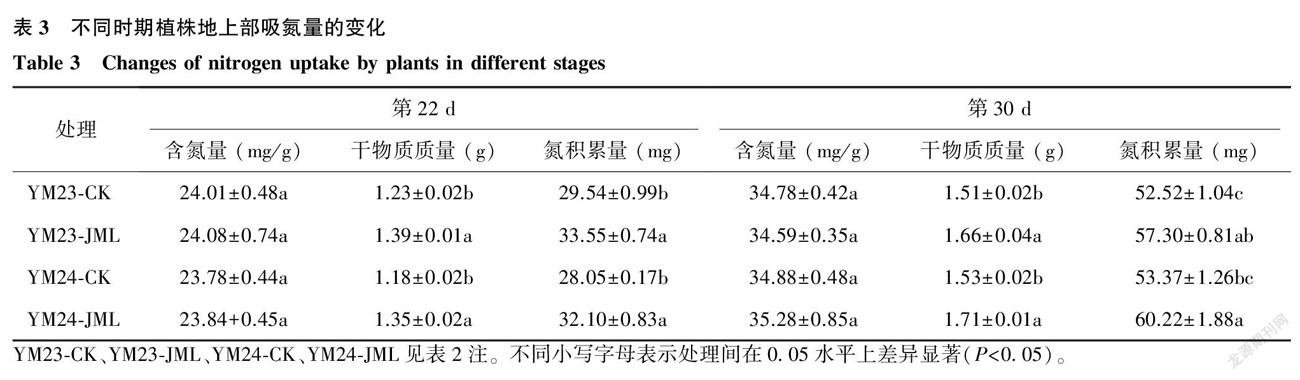
氮积累量的大小与植株生长程度密切相关。从表3中可以看出,JML处理后,两品种植株地上部吸氮量变化趋势一致,在第22 d、第30 d植株整体氮积累量较空白对照(CK)平均提高14.00%和10.96%,差异显著( P < 0.05),但植株含氮量并没有显著变化( P > 0.05)。表明JML处理能够明显提高植株的干物质质量,进而提高迟播冬小麦植株的整体氮积累量,从而利于植株生长,增强苗期抗逆性。
2.4 新型小麦种衣剂JML对小麦幼苗根系形态的影响
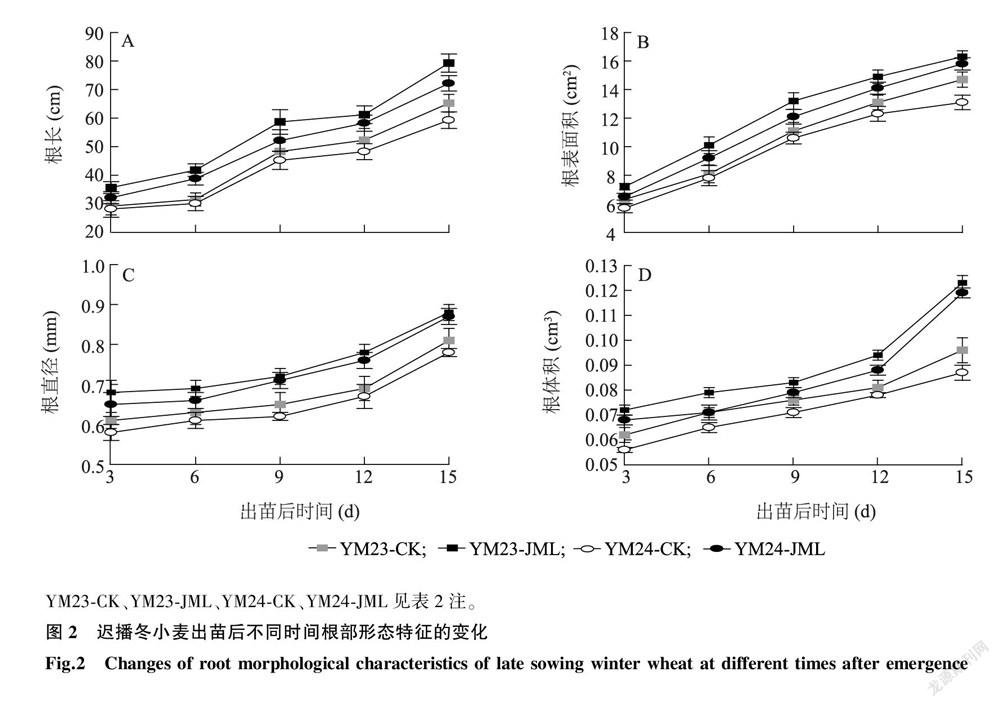
JML处理对小麦幼苗地下根系生长产生了明显的正向促进作用(图2)。从根系平均根长来看,随着生育进程的推移,两品种在第3 d、第6 d、第9 d、第12 d及第15 d的单株根系平均根长超过对照18.12%、30.72%、18.38%、18.85%及21.68%,差异达显著水平( P < 0.05)(图2A)。根系表面积平均显著增加14.16%、21.32%、16.53%、14.19%和15.75%(图2B)。根系平均直径平均超过对照11.77%、8.86%、12.64%、13.24%和10.09%(图2C),差异均达显著水平( P < 0.05)。根体积平均超过对照18.78%、10.25%、10.24%、14.43%和32.45%(图2D),差异均达显著水平( P < 0.05)。上述结果表明,JML处理显著增加了迟播小麦幼苗的根系整体素质。
2.5 新型小麦种衣剂JML对氮转运基因 TaNRT 、 TaAMT 表达的影响
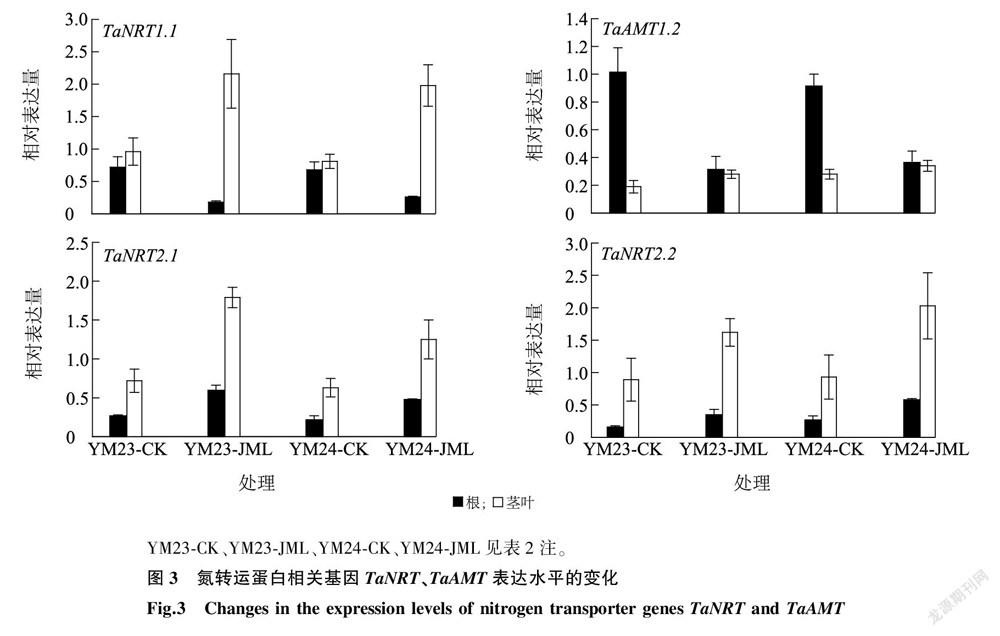
JML处理显著提高了氮转运蛋白 NRT 、 AMT 基因的表达水平(图3)。对于硝态氮(NO-3 -N)中的氮转运,JML处理后转运TaNRT1.1基因在叶片中表达水平平均提高1.35倍,差异显著( P < 0.05),而在根中表达水平与CK相比显著下降2.31倍( P < 0.05),从数值大小来看JML处理后 NRT 在根与茎的整体组合中显著超过CK(图3)。对于铵态氮(NH+4 -N)转运,JML处理后转运蛋白TaAMT1.2在根中的表达量显著低于CK,两品种平均降幅为1.89倍( P < 0.05),而在茎叶中其表达量与CK在两品种间差异均不显著( P > 0.05)。另一硝态氮转运蛋白家族基因TaNRT2.1、TaNRT2.2在两品种中变化趋势一致,JML处理后在根系与叶片中表达量均显著上调(图3)。这些差异表明JML处理后具有促进植株对不同形态氮素选择性吸收,促进了硝态氮在茎叶中的高效转运,但抑制了铵态氮在根系中的转运,这种差异应该与植株体内氮转运复杂的调节系统有关。
2.6 新型小麦种衣剂JML对氮同化过程中硝酸还原酶基因( TaNR )、亚硝酸还原酶基因( TaNIR )表达的影响
硝态氮吸收后转运输送至植株各器官进行同化作用,首先在硝酸还原酶( NR )的作用下转化为亚硝酸根,亚硝酸根接着在亚硝酸还原酶( NIR )的催化下生成铵根离子,而 NR 是这一阶段的关键限速酶。本研究发现JML处理后 TaNR 基因在植株根和叶片中均能检测到表达,JML处理与对照在根中的表达水平差异不显著( P > 0.05),但在叶片中JML处理较对照增幅平均为4.34倍,差异显著( P < 0.05)(图4)。从绝对值来看, TaNR 在植株中表达量占比优势从根中转至叶片中,暗示硝酸盐同化为亚硝酸盐过程中的高效化。另外, TaNIR 表达水平的变化与 TaNR 变化趋势一致,在根、叶片中均有表达,且两品种均在叶片中表达水平最高(图4B),表明亚硝酸盐向铵盐转化过程中的高效流畅性。
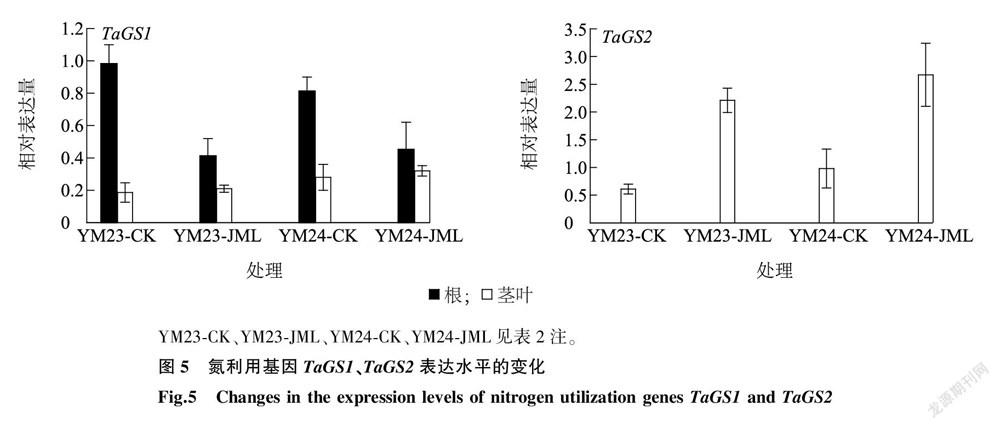
2.7 新型小麦种衣剂JML对氮利用基因TaGS1、TaGS2表达的影响
铵态氮需要在谷氨酰胺合成酶基因1(GS1)、谷氨酰胺合成酶基因2(GS2)的作用下生成谷氨酰胺,谷氨酰胺是进行其他小分子代谢的底物。从图5中可以看出,JML处理后根系中TaGS1的表达水平显著下降,差异显著( P < 0.05),但在茎叶中表达水平未下降,与CK无显著差异( P > 0.05)。
從图5中可以看出,基因TaGS2属于组成型表达,在根中表达量极低,但在叶片中高效表达。JML处理后TaGS2在两品种叶片中均显著上调,与CK均差异极显著( P < 0.01)。TaGS1、TaGS2的表达趋势不一致,推测可能与GS1主要为胞质型,而GS2为叶绿素型有关,两者受不同的氮代谢通路所调控。
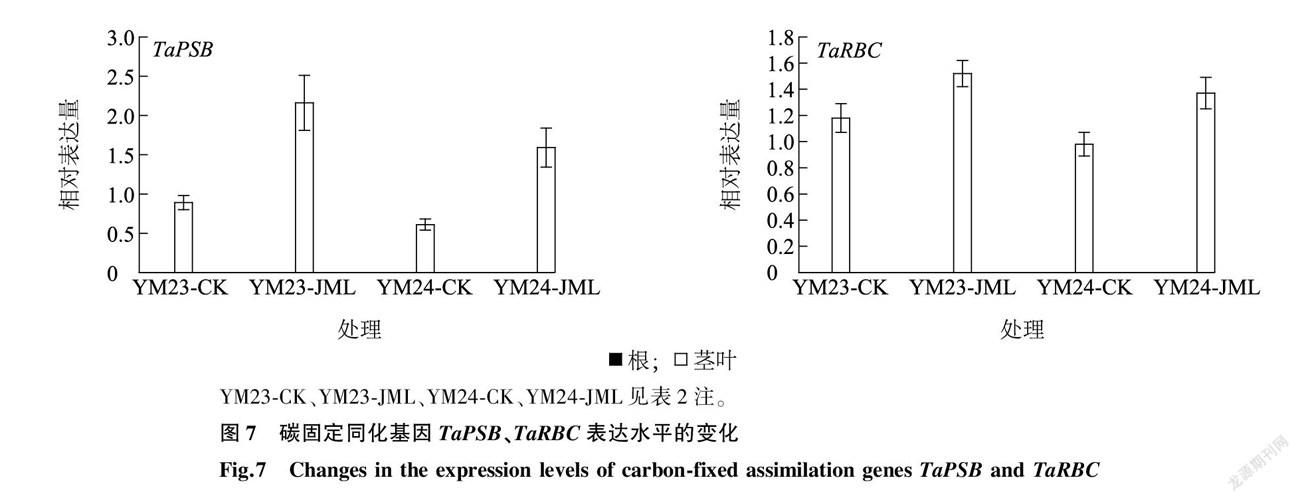
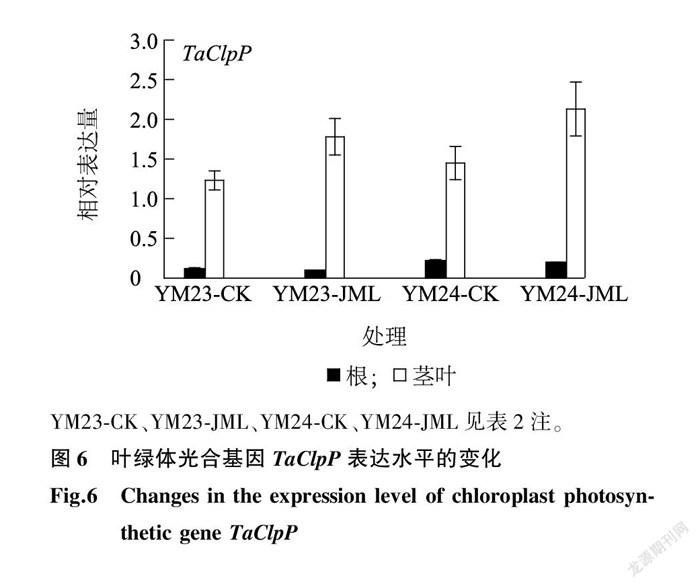
2.8 新型小麦种衣剂JML对碳代谢途径基因 TaClpP 、 TaPSB 和 TaRBC 表达的影响
光合作用是植物利用光能,同化CO2 制造碳水化合物的主要过程。光合基因 TaClpP 固定在叶绿体器官中,主要在小麦特定发育阶段表达。从图6中可以看出,JML处理后, TaClpP 在根中表达微量,品种间差异不大( P > 0.05),而在叶片中则表达水平较高,表达量与对照相比差异显著( P < 0.05)。
TaPSB (光合系统b)基因和 TaRBC (核酮糖-1,5-二磷酸羧化酶/加氧酶)基因是植物光系统反应中比较重要的2个基因,与CO2吸收与同化密切相关。从图7可以看出,在CO2 吸收与同化过程中 TaPSB 和 TaRBC 基因具有组成型表达的特点,在两品种根系中均未检测到表达,而在叶片中两者均高效表达。JML处理后小麦苗期叶片表达水平高于对照,差异显著( P < 0.05)。 TaRBC 是光合作用C3反应中重要的羧化酶, TaRBC 基因表达上调表明JML处理后植株叶片对无机态碳即CO2 的吸收更加高效,加速碳水化合物的形成。
3 讨 论
迟播冬小麦在苗期常伴随低温、冻害等不利因素,严重影响苗期的小麦群体质量 [3,20] 。合理的苗期群体结构是保障冬小麦实现稳产高产的前期物质基础,播种期延迟易引起出苗率降低、幼苗素质弱和群体干物质积累下降等问题 [3] 。前人通过加大播种量和增施氮肥等常规解决途径进行调控,但均不易形成冬前壮苗 [3,21] 。赵红梅等 [22] 的研究结果表明,旱地小麦逆境栽培通过膜覆土或膜际条播的方式获得增温效应使得小麦出苗时间提前,从而增加群体分蘖,促进冬前壮苗。另有研究发现,迟播小麦通过增加用种量能保证产量,但增加了茎秆基部节间长度,影响了株型和群体质量,增加小麦后期倒伏风险 [2] 。本研究结果表明,JML处理小麦种子后显著提高植株干物质质量与植株氮积累量,培育了健壮个体,能够增加迟播冬小麦的低温抗逆能力。种子活力指数是种子群体发芽速率和成苗率特征的综合反映,是种子能否正常使用的直接体现。沈奇等 [23] 研究发现种衣剂应用在油料作物紫苏中可以显著提高紫苏种子活力,提高出苗率,增加幼苗生长速度。本研究结果表明,JML处理小麦种子后可以显著提高冬小麦发芽势、发芽率及种子活力指数等各项关键生理指标,增强迟播冬小麦的出苗率和成苗率,促进幼苗生长,减少大田用种量。
氮素是影响前期植物群体质量和后期产量形成的关键元素,它参与植物体内的重要生理代谢 [24] 。小麦吸收的氮素以硝态氮(NO-3 -N)为主,硝态氮是小麦根系直接吸收与同化的主要氮素形态 [10,25] 。适量的氮肥增施能促进作物地下根系的健壮发育,增加根长、根体积和根质量,促进植株干物质的积累 [24,26] 。本研究结果表明,JML处理有效增大了根长、根直径、根体积和根表面积,促进了小麦根系的生长,增加对土壤水分与养分的吸收,利于培育壮根。已有研究结果显示,在籼稻和粳稻苗期转录组分析中发现,调控壮苗有关氮素吸收的关键 Hub 基因表达水平不同,可能与植株吸收氮素的形态差异有关 [27] 。迟播冬小麦尽管通过增施氮肥可以一定程度上增强幼苗素质,但却增加田间氮素的损失并降低氮素吸收利用率 [28] 。有研究结果表明,土壤中硝态氮的淋溶深度、淋失量与施氮量密切相关,在施氮量超过160 kg/hm2 的阈值后,硝态氮的淋失程度显著上升,而氮素吸收利用率显著下降 [29] 。 NR 是氮同化阶段的关键限速酶和诱导酶基因,其表达量大小与作物的氮素吸收利用效率高低密切相关 [30-31] 。本研究结果表明,JML处理促进了迟播冬小麦苗期对土壤表层硝态氮的吸收与利用,提高了植株氮积累量,显著提高了氮代谢中氮素吸收利用关键基因TaNRT1.1、 TaNR 、 TaNIR 、TaGS1、TaGS2等在植株中的表达水平,一定程度上降低了苗期氮素的损失,提高了氮素利用率。同时本研究还发现,JML处理抑制了迟播冬小麦根系对土壤表层铵态氮(NH+4 -N)的吸收与利用,铵态氮转运蛋白TaAMT1.2基因的表达水平在扬麦23、扬麦24品种中显著降低,但在叶片中的表达水平未有明显变化。这表明JML处理有助于小麦叶片对硝态氮的选择利用而降低了铵态氮利用,其内在机制需要进一步深入研究。JML处理促进小麦生长,应该与其含有内生真菌次生代谢产物有关的生物刺激素有关,而生物刺激素属于植物免疫诱抗剂类型。有研究结果表明,生物刺激素在促进植物生长和增强抗病性方面表现出较高活性 [32-35] ,激发植物体内水杨酸代谢通路,可诱导植株体内活性氧(ROS)积累及相关基因的表达,同时有效增加根尖生长素的含量,调节植株对氮素的吸收,以此促进植物生长 [36] 。
碳代谢主要表现为二氧化碳的同化,在细胞中以氨基酸作为主要底物合成各种蛋白质,参与植株体内各种代谢活动 [26,37-38] 。 PSB 和 RBC 基因是植物光合系统反应中比较重要的2个基因,是光合作用C3反应中重要的羧化酶基因,与CO2吸收与同化密切相关。本研究结果表明,JML处理后植株叶片 TaPSB 和 TaRBC 表达量显著上调,从碳代谢相关调控途径来看,JML处理后叶片对无机态碳即CO2 的吸收更加高效,从而加速碳水化合物的形成,利于迟播小麦植株生长。植物体内氮代谢与碳代谢互相影响。从提高迟播冬小麦幼苗质量考虑,JML处理可提高植株碳、氮途径上相关基因的表达水平,从而调控下游相关蛋白质的表达量,最终增强迟播小麦幼苗的抗逆能力和群体质量。
参考文献:
[1] 刘红江,郭 智,孙国峰,等.不同播种方式对小麦产量形成及氮素利用效率的影响[J].江苏农业学报,2019,35(5):1075-1081.
[2] 张明伟,马 泉,丁锦峰,等.密度与肥料运筹对迟播小麦产量和茎秆抗倒能力的影响[J].麦类作物学报,2018,38(5):584-592.
[3] 高德荣,张 晓,康建鹏,等.长江中下游麦区小麦迟播的不利影響及育种对策[J].麦类作物学报,2014,34(2):279-283.
[4] 吴金芝,黄 明,王志敏,等.极端晚播对小麦籽粒产量、氮素吸收利用和籽粒蛋白质含量的影响[J].应用生态学报,2018,29(1):185-192.
[5] 朱兆良. 农田中氮肥的损失与对策[J].土壤与环境,2000,9(1):1-6.
[6] 张福锁,崔振岭,王激清,等.中国土壤和植物养分管理现状与改进策略[J].植物学通报,2007,24(6):687-694.
[7] 顾大路,杨文飞,文廷刚,等. 冻害胁迫下防冻剂处理对小麦生理特征和产量的影响[J].江苏农业学报,2016,32(3):490-496.
[8] DIER M, MEINEN R, ERBS M, et al. Effects of free air carbon dioxide enrichment (FACE) on nitrogen assimilation and growth of winter wheat under nitrate and ammonium fertilization[J]. Global Change Biology,2018,24(1):40-54.
[9] WROBLEWITZ S, HUTHER L, MANDERSCHEID R, et al. The effect of free air carbon dioxide enrichment and nitrogen fertilisation on the chemical composition and nutritional value of wheat and barley grain[J]. Archives of Animal Nutrition,2013,67(4):263-278.
[10] ZHOU B, SERRET M D, ELAZAB A, et al. Wheat ear carbon assimilation and nitrogen remobilization contribute significantly to grain yield[J]. Journal of Integrative Plant Biology,2016,58(11):914-926.
[11] ALJAZAIRI S, ARIAS C, NOGUES S. Carbon and nitrogen allocation and partitioning in traditional and modern wheat genotypes under pre-industrial and future CO2 conditions[J]. Plant Biology (Stuttgart, Germany),2015,17(3):647-659.
[12] BERAUD J, FINE P, YERMIYAHU U, et al. Modeling carbon and nitrogen transformations for adjustment of compost application with nitrogen uptake by wheat[J]. Journal of Environmental Quality,2005, 34(2):664-675.
[13] PREY L, KIPP S, HU Y, et al. Nitrogen use efficiency and carbon traits of high-yielding european hybrid vs. line winter wheat cultivars: potentials and limitations[J]. Frontiers in Plant Science,2018, 9:1988.
[14] 赵君霞. 氮素及干旱胁迫对冬小麦幼苗生长和氮代谢相关基因表达的影响[D].郑州:河南农业大学,2015.
[15] 张小祥,李爱宏,蒋 敏, 等.新型水稻机插秧专用拌种剂的应用效果研究[J].江苏农业科学,2016,44(6):120-122
[16] 李星星,严青青,李春艳,等.种衣剂对棉花幼苗耐寒性的观测及耐寒种衣剂筛选[J].中国农业大学学报,2018,23(3):38-47.
[17] 张小祥,戴正元,孙成韬,等.新型玉米超微粉种衣剂的理化性质及田间应用效应[J].江苏农业科学,2014,42(7):92-94.
[18] 佟莉蓉,王 娟,张亚妮,等.不同种衣剂配方对达乌里胡枝子幼苗生长和生理特性的影响[J].草地学报,2020,28(3):844-851.
[19] 许 晨,王文静,曹 珊,等.花后DA-6处理调控小麦种子活力的机理[J].中国农业科学,2021,54(9):1821-1834.
[20] 文廷刚,钱新民,王伟中,等.不同生长调节剂拌种对小麦种子发芽及幼苗素质的影响[J].江苏农业科学,2017,45(1):59-61.
[21] 张金宝,秦 霞,孙佩贤,等.黄淮麦区种植密度对晚播冬小麦花后氮素代谢和利用率的影响[J].西北农林科技大学学报(自然科学版),2010,38(12):112-116,122.
[22] 赵红梅,高志强,孙 敏,等.休闲期耕作对旱地小麦土壤水分、花后脯氨酸积累及籽粒蛋白质积累的影响[J].中国农业科学,2012,45(22):4574-4586.
[23] 沈 奇,杨 森,徐 静,等. 种衣剂对紫苏发芽率及产量品质性状的影响[J].中国农学通报,2018,34(28):21-25.
[24] TIAN H, YUAN X, DUAN J, et al. Influence of nutrient signals and carbon allocation on the expression of phosphate and nitrogen transporter genes in winter wheat ( Triticum aestivum L.) roots colonized by arbuscular mycorrhizal fungi [J]. PLoS One, 2017, 12(2):e0172154.
[25] 蒋会利,温晓霞,廖允成.施氮量对冬小麦产量的影响及土壤硝态氮运转特性[J].植物营养与肥料学报,2010,16(1):237-241.
[26] BLOOM A J, SMART D R, NGUYEN D T, et al. Nitrogen assimilation and growth of wheat under elevated carbon dioxide [J]. Proc Natl Acad Sci USA, 2002, 99(3):1730-1735.
[27] ZHANG X X, ZHOU J, HUANG N S, et al. Transcriptomic and co-expression network profiling of shoot apical meristem reveal contrasting response to nitrogen rate between indica and japonica rice subspecies [J]. International Journal of Molecular Sciences, 2019, 20(23): 5922.
[28] 张 嫚,周苏玫,杨习文,等.减氮适墒对冬小麦土壤硝态氮分布和氮素吸收利用的影响[J].中国农业科学,2017,50(20):3885-3897.
[29] 戴 健,王朝辉,李 强,等.氮肥用量对旱地冬小麦产量及夏闲期土壤硝态氮变化的影响[J].土壤学报,2013,50(5):956-965.
[30] YIN X M, LUO W, WANG S W,et al. Effect of nitrogen starvation on the responses of two rice cultivars to nitrate uptake and utilization[J]. Pedosphere,2014,24(5):690-698.
[31] GUO T C, XUAN H M, YANG Y Y, et al. Transcription analysis of genes encoding the wheat root transporter NRT1 and NRT2 families during nitrogen starvation[J]. Journal of Plant Growth Regulation,2014,33(4):837-848.
[32] 龙 伟,姚小华, 吕乐燕,等. 油茶种子性状及浸种后内源激素含量分析[J].南京林业大学学报(自然科学版),2020,44(5):148-156.
[33] 戴 毅,田龙果,潘贞志,等. 激素和非生物逆境胁迫调控植物硝酸盐转运蛋白功能的研究进展[J].江苏农业学报,2020,36(6):1595-1604.
[34] 刘 果,高丽琼,彭 彦,等. 外源激素对低温胁迫下南美油藤幼苗叶片生理生化指标的影响[J].南方农业学报,2020,51(9):2181-2190.
[35] 王 鹏,郭文英,王文静. 苦皮藤愈伤组织诱导体系建立[J].江苏农业科学,2019,47(23):92-95.
[36] LU C, LIU H, JIANG D, et al. Paecilomyces variotii extracts (ZNC) enhance plant immunity and promote plant growth[J]. Plant Soil, 2019,441:383-397.
[37] MILLER Z J, LEHNHOFF E A, MENALLED F D, et al. Effects of soil nitrogen and AJMLospheric carbon dioxide on wheat streak mosaic virus and its vector ( Aceria tosichella Kiefer) [J]. Plant Disease, 2015, 99(12):1803-1807.
[38] SINHA S K, RANI M, BANSAL N, et al. Nitrate starvation induced changes in root system architecture, carbon:nitrogen metabolism, and miRNA expression in nitrogen-responsive wheat genotypes [J]. Applied Biochemistry and Biotechnology, 2015, 177(6):1299-1312.
(責任编辑:陈海霞)
收稿日期:2021-09-10
基金项目:国家现代农业产业技术体系建设专项(CARS-01-60);江苏省重点研发计划项目(BE2018362);江苏省农业科技自主创新基金项目[CX(18)1002, CX(19)3060];扬州市社会发展项目(YZ2019050)
作者简介:张小祥(1980-),男,江苏东台人,博士,副研究员,主要从事作物栽培与壮苗生理调控。(E-mail)zhngyz@126.com
通讯作者:姚友礼,(E-mail)youyl@yzu.edu.cn;黄年生,(E-mail)jsyzhns@163.com
