椴树红松林掘根倒木及其微立地对凋落叶分解的影响
2022-07-15段文标朱帅威陈立新
段文标,朱帅威,陈 佳,陈立新
(东北林业大学 林学院,黑龙江 哈尔滨 150040)
风是森林生态系统的主要干扰源[1]。自上个世纪以来,风引起的森林损害大幅度增加[2]。树木连根拔起(倒下的树木称为“掘根倒木”)是森林生态系统中风干扰的主要形式之一[3]。它将埋藏的土壤养分和植物碎屑等暴露在大气和地面中,不仅重新分配了森林生态系统中的土壤、生物量和养分[4],更重要的是创造了独特的丘-坑微立地[5]。从生态学的角度来看,丘-坑微立地与未受干扰的微立地有很大区别[6],它们为树木更新提供了特定的生态位[7-8],增加了陆地生态系统的生物、生态多样性[9-11]和森林管理决策的复杂性[6],对土壤形成、土壤形态沉积物移动、排水模式、养分循环、森林结构及其生态功能等特征有潜在影响[12]。掘根倒木及其形成的丘-坑微立地是许多森林底部的重要组成部分[6]。例如在温带原始森林中,大约30%的树木因掘根倒木而死亡[13];在未管理的森林中,有23%~42%的树木因掘根倒木而死,而不是干中折断或立枯死亡[14]。此外,确定掘根倒木及其形成的丘-坑微立地的年龄,对于理解它们在自然生态系统中的动态变化也至关重要。所以,在生态功能衰弱甚至退化的森林中,开展掘根倒木及其丘-坑微立地的相关研究十分必要。
分解是生态系统恢复的重要考虑因素[15]。森林凋落物的分解是一个重要的生物地球化学过程[16],是全球碳收支和养分周转的重要组成部分[17],也是陆地森林生态系统可持续性的必要条件[18],控制着土壤有机质的形成、养分循环、能量流动和植物生长[19]。数据显示,相比于枝、干、皮、根,凋落叶的贡献率最高[20]。凋落叶混合分解是自然界常见的现象[20]。混合分解机制通常不同于单一组分凋落叶,其分解特点是在森林底层内,非生物和生物因素共同控制而发生的相互作用[21]。作为连接地上和地下物质循环之间的生态界面[17,22],它将植物有机物和营养元素转化为土壤有机和无机物质,并由微生物群落驱动[23]。研究表明,分解过程中的质量变化和凋落物层的微环境也会影响微生物群落的结构和功能[24],从而影响凋落叶的分解速率。众所周知,分解率已被用来表示凋落叶的分解动态,单位时间内其值越大表示凋落叶分解越快[25-27],而凋落叶养分元素的含量在很大程度上也影响分解速率[28]。因此,更好地理解凋落叶分解影响因素的相对贡献对于预测生态系统过程和功能非常重要[29]。自20世纪70年代初以来,研究者们对凋落叶的分解过程及其影响因素进行了广泛的探索[30-31],但缺乏凋落叶与微环境之间的研究,尤其是缺乏与掘根倒木及其形成的丘-坑微立地之间的研究。从研究区域看,在温带小兴安岭地区的相关研究也不充分。所以,在掘根倒木频繁出现的小兴安岭地区,深入研究掘根倒木及其形成的丘-坑微立地对凋落叶分解的影响非常紧迫。
目前,小兴安岭地区因风干扰而导致掘根倒木现象明显增多[32],使得该区受风干扰的部分生态系统的稳定性有所降低。所以,本研究选取黑龙江省伊春市带岭区凉水国家级自然保护区具有典型丘-坑微立地的代表性林型——椴树红松林(Tilia amurensis-Pinus koraiensis forest)内的红松Pinus koraiensis、紫椴Tilia amurensis和色木槭Acer mono这3 种优势树种的混合凋落叶为研究对象,分析凋落叶在掘根倒木微立地条件下的质量和养分变化动态,进一步探究掘根倒木的腐烂等级及其形成的不同类型微立地对凋落叶分解的影响,以期更好地确定掘根倒木及其丘-坑微立地对椴树红松林生态系统的作用,从而为提升其稳定性和生产力等提供科学依据。
1 研究区与研究方法
1.1 研究区概况
研究区设在黑龙江省伊春市带岭区凉水国家级自然保护区,位于小兴安岭山脉的东南段—达里带岭支脉的东坡,地理坐标为47°06′49″~47°16′10″N,128°47′08″~128°57′19″E。该区属于温带大陆性季风气候,冬季气候严寒且干燥;降水多集中在6—8月,雨热同期。年平均气温只有-0.3℃,年均降水量为676 mm。地带性土壤为暗棕壤。主要的森林类型为以红松为主的针阔混交林,所选样地内的主要组成树种为红松、紫椴、臭冷杉Abies nephrolepis、红皮云杉Picea koraiensis、色木槭Acer mono、花楷槭Acer ukurunduense和春榆Ulmus davidiana。

表1 椴树红松林样地种群特征Table 1 Species characteristics of the sample plots in Tilia amurensis-Pinus koraiensis forests
1.2 试验设计
通过踏查筛选出由掘根倒木形成的具有典型丘-坑微立地的代表性林型—椴树红松林,在该林型内设置1.2 hm2(100 m×120 m)的矩形样地。根据Rouvinen 等[33]倒木腐烂等级五级划分法,选择具有完整坑底、丘面微立地的Ⅱ级、Ⅲ级和Ⅳ级共3 个腐烂等级的掘根倒木18 株,并将掘根倒木(Uprooted treefalls,UT)、坑底(Pit bottom,PB)和丘面(Mound face,MF)微立地设置为样方,每一种微立地选取3 个样方作为重复;同时将掘根倒木形成的林隙内未受掘根干扰的微立地称为林隙微立地(以下简称林隙,Forest gap,FG),将倒木形成的林隙外未受掘根干扰的完整的临近的郁闭林分下的微立地称为郁闭林分微立地(以下简称林分,Intact stand,IS),同时将林隙及其郁闭林分设置为对照组。

图1 掘根倒木丘-坑微立地示意图Fig.1 Diagram of uprooted treefalls and their pit-mound microsites
1.3 凋落物的收集及处理
2019年6月,收集上一年自然凋落的、上层较为新鲜(凋落叶保持原有状态,颜色变化不明显,外表无明显分解痕迹)的半分解状态的红松、紫椴和色木槭凋落叶,带回实验室用流动的水流冲洗上面的泥土,自然风干后装入分解袋内,分解袋内3 种叶片的比例依据3 种优势树种在IS 中所占的自然比例(2∶1∶1)而定,每袋称取10 g,其中红松5.0 g,紫椴2.5 g,色木槭2.5 g,误差不超过0.01 g。尼龙分解袋规格为15 cm×10 cm,网眼大小为1 mm;总计数量为165 袋,其中每个样方多放2 袋凋落叶为备用,以免意外情况的发生;在放置时凋落叶袋之间不重叠,紧贴地表,用少量表层土壤覆盖,使其接近自然状态与土壤充分接触。留下一部分自然风干后未装入分解袋的凋落叶用于初始值(背景值)的测定。
2019年7月初放置凋落叶分解袋,于2019年8—10月每隔30 d 各取样一次,共取样3 次,每次取33 袋凋落叶(11 个取样点×3 次重复),带回实验室。将样品放在0.149 mm 网筛上小心冲洗,洗去沉积物。清洗凋落叶上的泥沙和杂物后,分别封入纸袋,置于鼓风干燥箱,80℃下烘干至恒质量后称取其干质量,后将凋落叶粉碎并过0.25 mm 筛、保存备测。
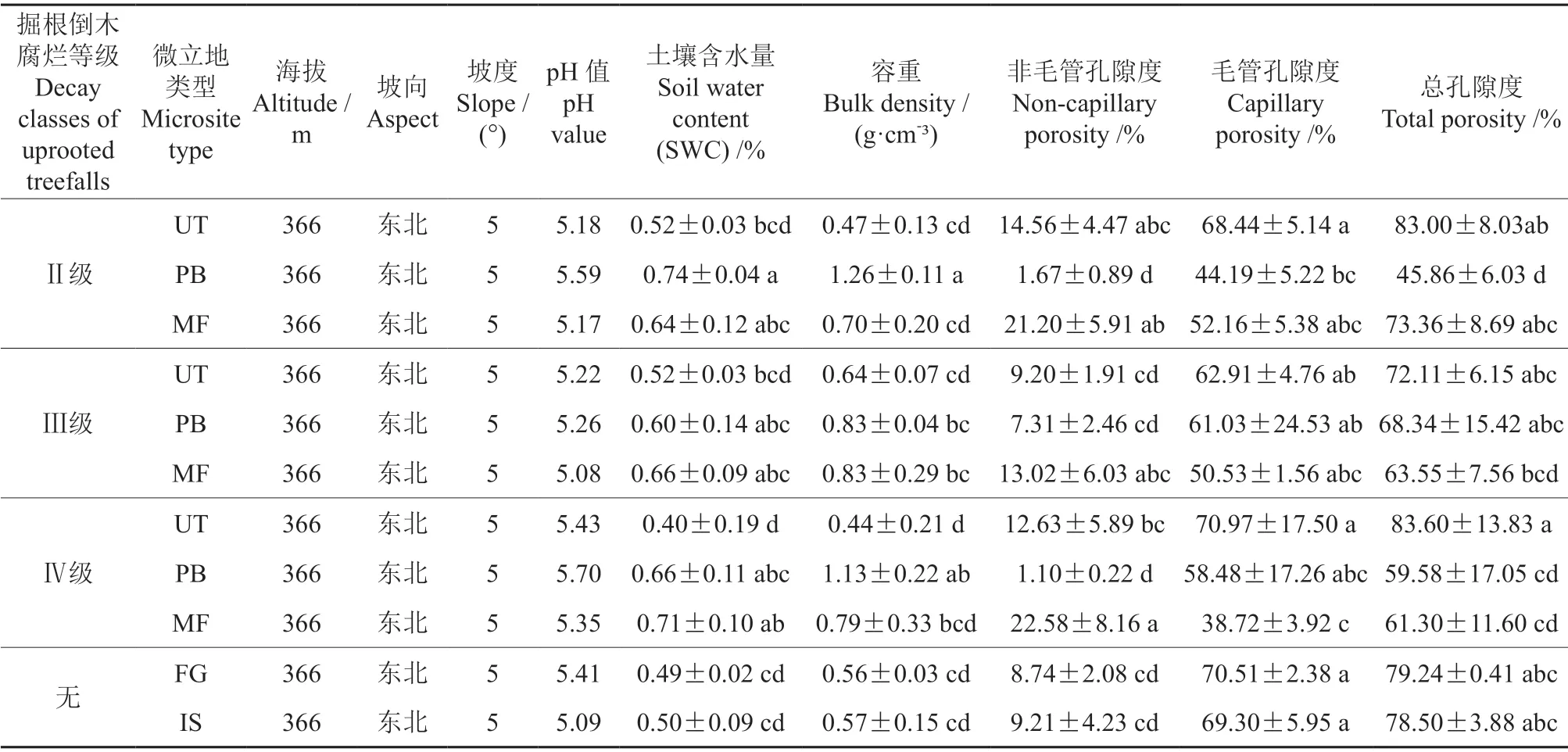
表2 椴树红松林样地概况及土壤性质†Table 2 General situations and soil properties in the sample plots in Tilia amurensis-Pinus koraiensis forests
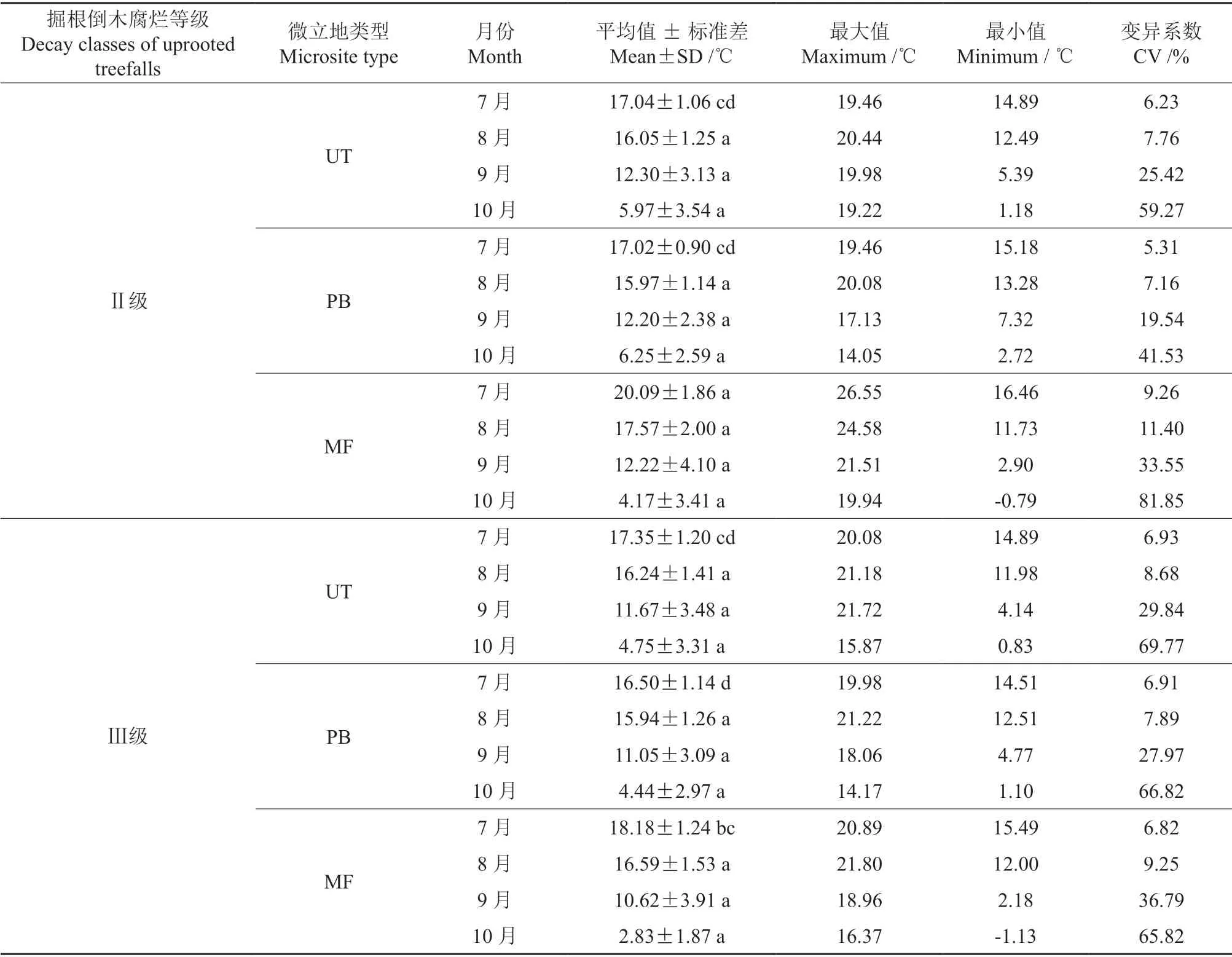
表3 椴树红松林内各微立地不同月份的土壤温度统计特征Table 3 Statistical characteristics of soil temperature in different months in each microsite in Tilia amurensis-Pinus koraiensis forests
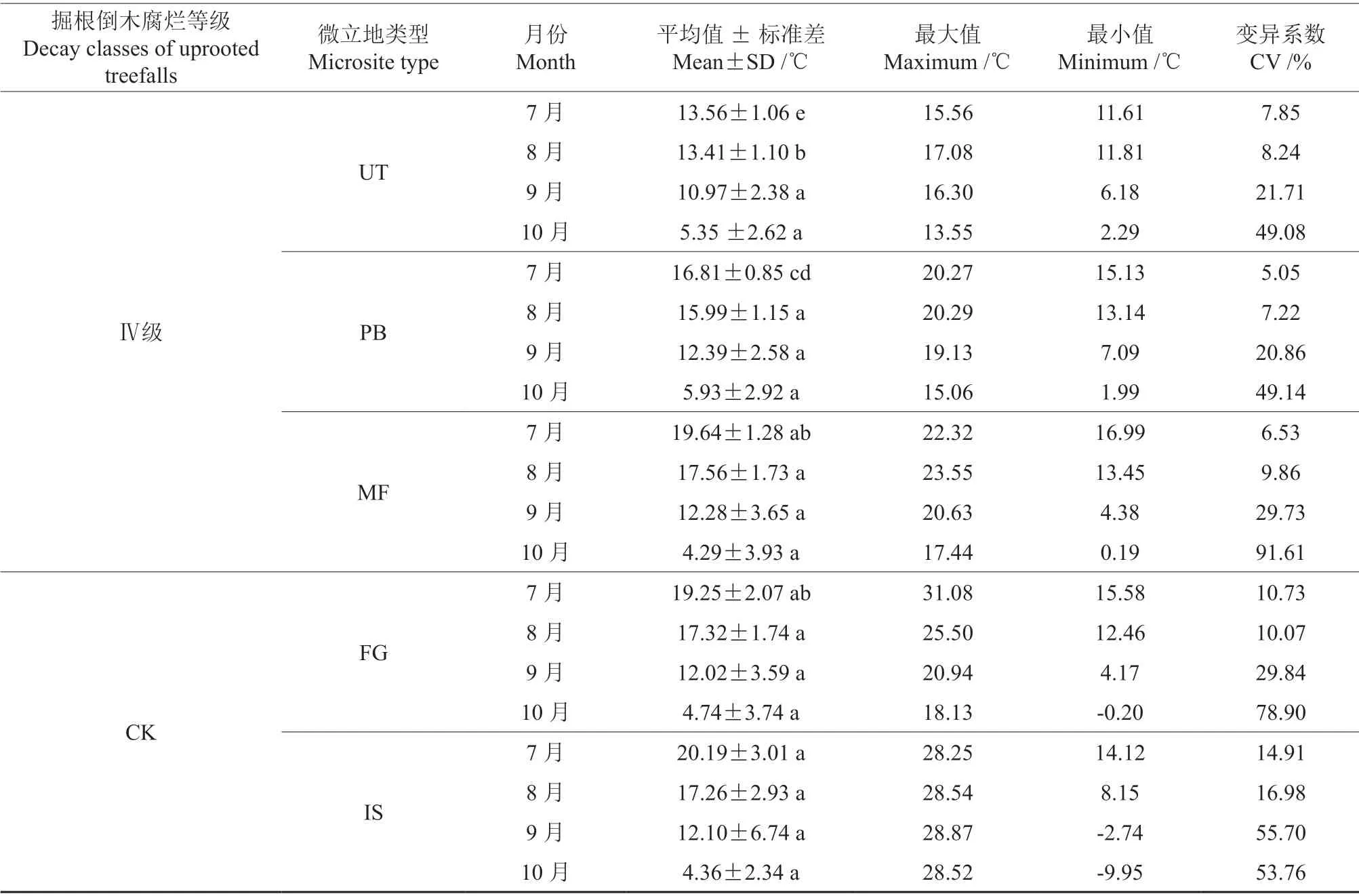
续表3Continuation of table 3
1.4 凋落叶组分指标的测定
凋落叶组分测定指标包括全碳、全氮、全磷含量。全碳含量利用总有机碳氮分析仪(TOC)测定;全氮含量采用浓硫酸-高氯酸消煮,凯氏定氮法测定;全磷含量采用浓硫酸-高氯酸消煮,钼锑抗显色法测定。

表4 凋落叶的初始养分含量及其比值Table 4 Initial nutrient content and ratio of litter
1.5 数据处理
凋落叶质量分解率:
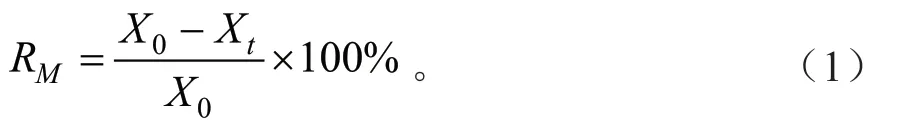
式中:X0为凋落叶初始质量(g);Xt为凋落叶分解t时间后剩余质量(g)。
分解速率k用修正后Olson 指数衰减模型[34]:

式中:y为凋落叶质量残留率(%);a为拟合参数;k为分解系数(g·g-1·a-1);t为时间(a)。
凋落叶分解50%(T50%)和95%(T95%)所需时间为[34]:


凋落物养分释放率[35]:
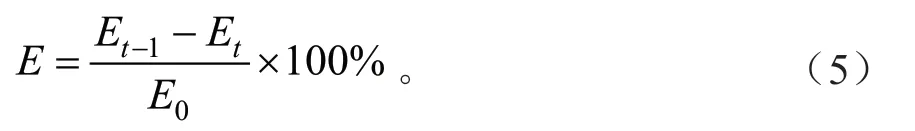
式中:E为第t次采样时凋落物的养分释放率(%);Et-1为第t-1 次采样时凋落物养分残留量(g/kg);Et为第t次采样时凋落物养分残留量(g/kg);E0为凋落物养分初始含量(g/kg)。
使用Excel 2016 软件计算各项指标的平均值和标准偏差,并完成Olson 衰减指数模型的拟合。利用软件SPSS 22.0 软件完成统计分析,使用Origin 2018 软件作图。采用最小显著差异法(LSD)检验掘根倒木腐烂等级和微立地类型对凋落叶养分释放的影响。采用三因素方差分析法(Three-way ANOVA)探究掘根倒木腐烂等级、微立地类型和分解时间对质量分解率、养分释放率及其养分元素比的影响。采用Pearson 相关性分析法探讨不同腐烂等级掘根倒木和微立地类型与凋落叶质量分解率、碳、氮、磷剩余含量及其比值之间的相关性。
2 结果与分析
2.1 掘根倒木腐烂等级和微立地类型对凋落叶质量分解率的影响
掘根倒木腐烂等级、微立地类型和分解时间均极显著影响凋落叶的质量分解率(P<0.01),但他们的交互作用对其影响不显著(P>0.05,表5)。
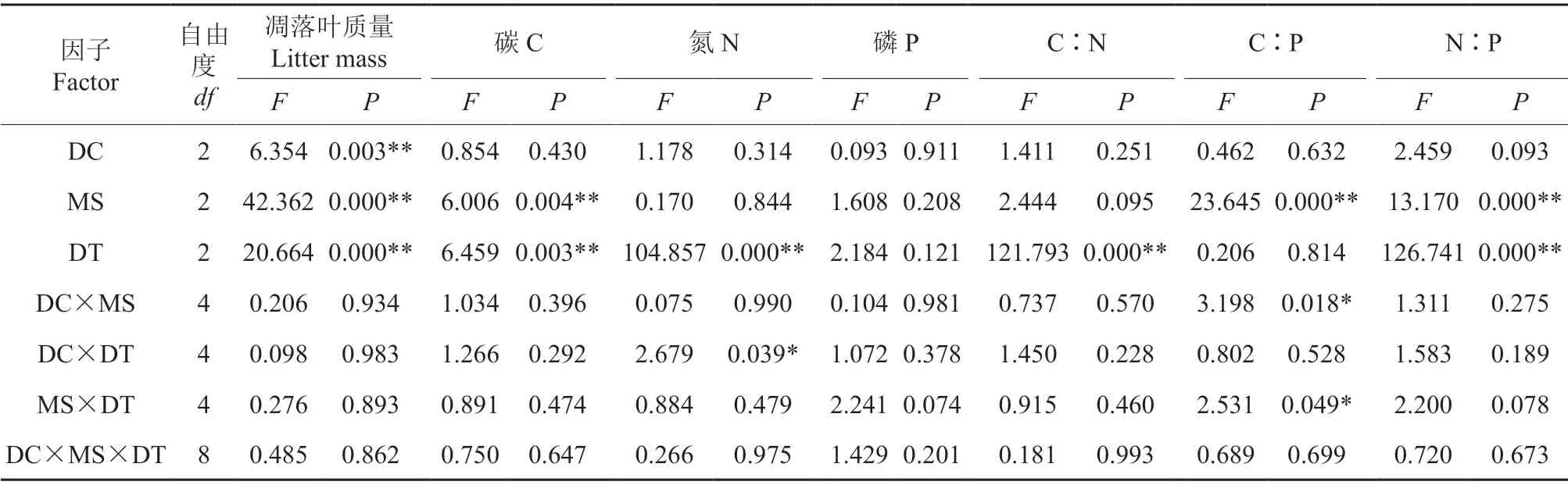
表5 掘根倒木腐烂等级、微立地类型和分解时间对凋落叶质量和养分元素的分解率及元素比影响的三因素方差分析†Table 5 Three-way ANOVA analysis of the effect of decay classes of uprooted treefalls,microsite types and decomposition time on the decomposition rate of litter mass,nutrient element and element ratios
凋落叶分解90 d 后,对照组FG 和IS 的凋落叶分解率分别为17.73%和19.10%,各掘根倒木腐烂等级下UT、PB 和MF 微立地的凋落叶分解率为10.10%~22.30%(图2)。其中,在Ⅱ级掘根倒木腐烂等级中,3 种微立地类型下的凋落叶分解率为10.10%~21.13%,依次为UT>IS>FG>MF>PB;在Ⅲ级掘根倒木腐烂等级中,3 种微立地类型下的凋落叶分解率为15.03%~22.30%,依次为MF>UT>IS>FG>PB;在Ⅳ级掘根倒木腐烂等级中,3 种微立地类型下的凋落叶分解率为13.27%~20.70%,依次为UT>IS>MF>FG>PB。可知,PB 微立地的凋落叶分解速率最慢,MF微立地次之,UT 微立地最快。另外,在同一微立地类型条件下,各掘根倒木腐烂等级的凋落叶分解率依次为Ⅲ级(19.65%)>Ⅳ级(17.27%)>Ⅱ级(15.39%)。可见凋落叶的质量分解程度与掘根倒木腐烂等级和微立地类型之间均存在一定的关系。
2.2 掘根倒木腐烂等级和微立地类型对凋落叶分解常数k 的影响
为了说明掘根倒木腐烂等级和微立地类型与凋落叶分解速率常数k之间的关系,利用Olson 提出的指数衰减模型,对不同微立地下的质量残留率(y)与时间(t)进行拟合,得到的衰减模型的相关系数大多数达到显著水平(表6),说明能够较好地模拟凋落叶的分解动态(R2>0.75,P<0.01)。从表6可以看出,UT、PB 和MF 这3 个微立地类型下,Ⅱ级掘根倒木腐烂等级的凋落叶分解速率分别为1.19、0.37 和0.90;Ⅲ级掘根倒木腐烂等级的凋落叶分解速率分别为1.47、0.72 和1.23;Ⅳ级掘根倒木腐烂等级的凋落叶分解速率分别为1.42、0.45 和1.06;对照组IS 和FG 的凋落叶分解速率常数分别为1.23 和1.17。一般以95%凋落叶被分解所需的时间来表示凋落叶的周转期,UT、PB 和MF 这3 个微立地类型下,Ⅱ级掘根倒木腐烂等级的凋落叶周转期分别为2.51、8.21 和3.33 a;Ⅲ级掘根倒木腐烂等级的凋落叶周转期分别为2.04、4.17 和2.44 a;Ⅳ级掘根倒木腐烂等级的凋落叶周转期分别为2.10、6.66 和2.83 a;对照组IS和FG 的凋落叶周转期分别为2.57 和2.44 a。由于分解速率常数k值是表征凋落叶分解速率的常用指标,k值越大,凋落叶分解越快。
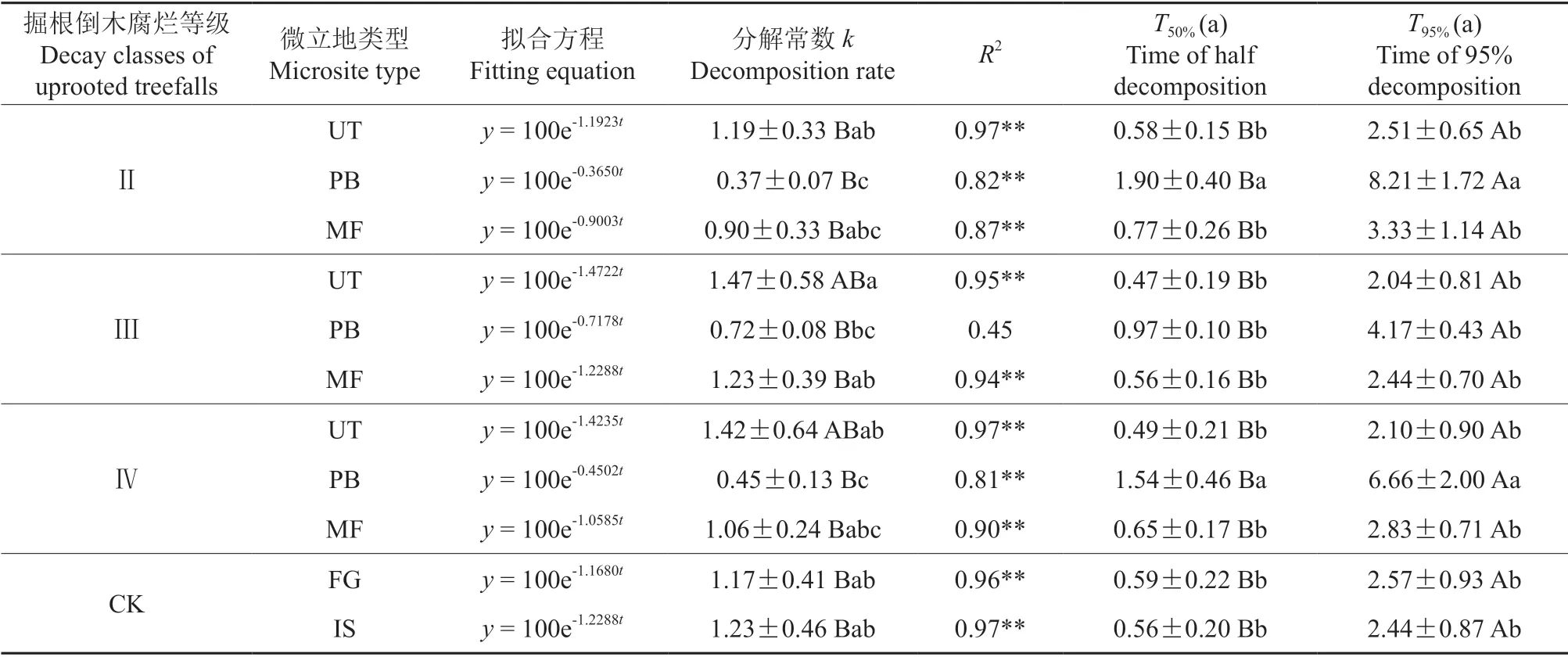
表6 不同腐烂等级掘根倒木下各微立地上的凋落叶残留率随时间的指数回归方程†Table 6 Exponential regression equation of the residual rate of litter on each microsite under different decay classes of uprooted treefalls with the development of time
在同一微立地类型、不同掘根倒木腐烂等级条件下,凋落叶表现出如下特点:Ⅱ级、Ⅲ级和Ⅳ级PB 的凋落叶完成95%分解所需时间分别比IS 对照推迟了5.77、1.73 和4.22 a;MF 的凋落叶推迟了0.89、0 和0.39 a;UT 的凋落叶提前了-0.07、0.40和0.34 a。Ⅱ级、Ⅲ级和Ⅳ级PB 的凋落叶完成95%分解所需时间分别比FG 推迟了5.64、1.60 和4.09 a;MF 的凋落叶推迟了0.76、-0.13 和0.26 a;UT 的凋落叶提前了0.06、0.53 和0.47 a。由上述可知,Ⅲ级、Ⅳ级掘根倒木腐烂等级的凋落叶分解速率大于Ⅱ级掘根倒木腐烂等级的凋落叶分解速率;在同一掘根倒木腐烂等级下,UT 和MF 微立地类型下的凋落叶分解速率大于PB 微立地类型下的凋落叶分解速率。
2.3 掘根倒木腐烂等级和微立地类型对凋落叶养分释放的影响
微立地类型和分解时间均极显著影响凋落叶的C 元素释放率(P<0.01,表5)。由图3可知,凋落叶分解至第30 天时,各个微立地类型下的凋落叶C 表现为释放;分解至第60 天时,Ⅱ级、Ⅲ级UT、MF 及Ⅳ级MF 的凋落叶C 表现为富集,其他微立地的凋落叶C 表现为释放;分解至第90天时,Ⅱ级、Ⅲ级UT 和Ⅱ级PB 的凋落叶C 表现为释放,其他微立地的凋落叶C 表现为富集。分解至第30 天和第60 天时,Ⅱ级、Ⅲ级和Ⅳ级的凋落叶C 释放率以PB 微立地最大,分解至第90天时,各个掘根倒木腐烂等级的凋落叶C 释放率以MF 微立地最大。整个分解期间,各个掘根倒木腐烂等级各微立地的凋落叶表现为明显的C 释放现象,PB 微立地的释放程度最高。
分解时间以及分解时间与掘根倒木腐烂等级的交互作用均显著影响凋落叶的N 元素释放率(P<0.05,表5)。由图3可知,凋落叶分解至第30 天时,FG、IS 和Ⅱ级MF 微立地的凋落叶N表现为富集,各掘根倒木腐烂等级下UT 和PB 微立地的凋落叶N 元素表现为释放;分解至第60 天时,Ⅱ级、Ⅳ级UT 和Ⅱ级PB 的凋落叶N 表现为富集,其他微立地的凋落叶N 表现为释放;分解至第90 天时,对照组以及各个微立地的凋落叶N元素均表现为不同程度的释放。分解至第30 天时,凋落叶N 富集率以FG、IS 对照组最大,分解至第60 天和第90 天时,凋落叶N 释放率以FG、IS对照组最大,其中分解到第90 天时,Ⅱ级UT、PB、MF 微立地的凋落叶N 释放率大于Ⅲ级、Ⅳ级的UT、PB、MF 微立地的凋落叶N 释放率。整个分解期间,各个掘根倒木腐烂等级的凋落叶表现为明显的N 释放现象,各丘坑微立地的N 释放程度相近。
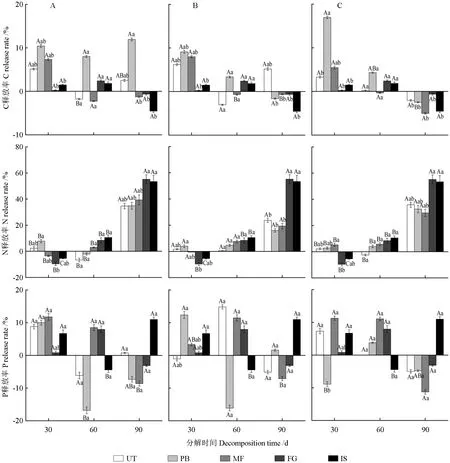
图3 不同腐烂等级掘根倒木下各微立地凋落叶C、N、P 元素的释放进程Fig.3 Release process of C,N and P elements in litter at different microsites under different decay classes of uprooted treefalls
各掘根倒木腐烂等级和微立地类型对凋落叶的P 元素释放率的影响均不存在显著性关系(P>0.05,表5)。由图3可知,凋落叶分解至第30 天时,Ⅲ级UT、Ⅳ级PB 的凋落叶P 表现为富集,其他微立地的凋落叶P 表现为释放;分解至第60 天时,Ⅱ级UT、Ⅱ级PB、Ⅲ级PB 和IS 的凋落叶P 表现为富集,其他微立地的凋落叶P 表现为释放;分解至第90 天时,Ⅱ级UT、Ⅲ级PB 和IS 的凋落叶P 表现为释放,其他微立地的凋落叶P 表现为富集。分解至第30 天时,各个掘根倒木腐烂等级的凋落叶P 释放率以Ⅲ级PB 微立地最大;分解至第60 天时,各个掘根倒木腐烂等级的凋落叶P富集率以Ⅱ级PB 微立地最大;分解至第90 天时,各个掘根倒木腐烂等级的凋落叶P 富集率以Ⅳ级MF 微立地最大。整个分解期间,各个掘根倒木腐烂等级的凋落叶表现为明显的P 释放现象,Ⅲ级UT 微立地的释放程度最高。
2.4 凋落叶质量分解率与剩余养分元素指标的关系
由表7可知,经过90 d 分解,对照组IS 的凋落叶质量分解率与N 剩余含量、N∶P 显著负相关,与C∶N 显著正相关,而与C、P 剩余含量和C∶P无显著相关关系。对照组FG 的凋落叶质量分解率与各养分剩余含量及其比值均无显著相关关系。Ⅱ级UT 微立地的凋落叶质量分解率与N 剩余含量显著负相关,与C∶N 显著正相关。Ⅱ级PB 微立地的凋落叶质量分解率与C、N、C∶P 显著负相关。Ⅲ级PB 微立地的凋落叶质量分解率与N剩余含量极显著负相关,与C∶N 极显著正相关。各级掘根倒木腐烂等级,MF 微立地的凋落叶质量分解率与各养分剩余含量及其比值均无显著相关关系。而N∶P 与Ⅱ级、Ⅳ级掘根倒木腐烂等级下的UT 和PB 微立地的凋落叶质量分解率显著、极显著负相关。

表7 不同掘根倒木腐烂等级和微立地类型下凋落叶质量分解率与碳、氮、磷剩余含量及其比值的相关性†Table 7 The correlation between mass loss rates and contents and the ratios of residual total carbon,nitrogen and phosphorus in different decay classes of uprooted treefalls and microsites
3 讨 论
3.1 掘根倒木腐烂等级和微立地类型以及分解时间对凋落叶分解的影响
凋落叶分解是陆地生态系统物质循环和能量流动的一个关键过程[36-37]。作为连接植物和土壤的重要生物因子,土壤微生物的活动、群落结构和多样性都会影响凋落物分解[38-39]。研究凋落叶的分解过程有助于加强对凋落叶质量衰减过程中C、N、P 营养物质行为的理解[40-41]。本研究中凋落叶在分解90 d 后,不同腐烂等级的凋落叶分解率依次为Ⅲ级(19.65%)>Ⅳ级(17.27%)>Ⅱ级(15.39%);不同微立地的凋落叶分解率依次为UT>MF>PB。
各腐烂等级各微立地的凋落叶分解动态基本符合Olson 衰减指数模型,分解常数为0.37~1.47,分解50%和95%所需时间分别为0.47~1.90 a 和2.04~8.21 a。其中UT、PB 和MF 这3 种微立地的凋落叶分解常数分别为1.19~1.47、0.37~0.72和0.90~1.23;对照组IS 和FG 的分解常数分别为1.23 和1.17。PB 微立地的凋落叶分解常数与长白山次生针阔混交林[42]的凋落叶分解常数(0.27~0.76)相接近,MF 和UT 微立地的凋落叶分解常数与海南岛尖峰岭热带针叶林[43]的凋落叶分解常数(0.84~1.58)相接近。吉林蛟河天然红松阔叶林中,3 个树种的分解常数k值分别为:红松(0.53±1.03)、紫椴(1.60±0.52)、色木槭(1.50±1.81)[44],本试验中3 种凋落物混合,k值大部分介于其间,极端低值出现于PB 微立地。3 种微立地的凋落叶分解常数相差较大,这可能是与微立地之间的气候条件、土壤物理性质和土壤养分状况等微环境有关。已有研究表明[45],在这3 种微立地中,UT 微立地的土壤物理性质最好,MF 微立地次之,PB 微立地最差;UT 和MF 微立地的凋落叶分解较快,可能是因为这两处微立地的土壤较肥沃,土壤环境较稳定。而PB 微立地在生长季经历了暴晒和水涝等情况,使得此微立地的微环境失衡,导致微生物活动受限,从而减缓了凋落叶的分解速度。本研究的分解常数较北方针阔混交林[46]的凋落叶分解常数(0.25~1.08)略有偏高。虽然凋落物分解速率与纬度呈极显著负相关(P<0.001)[47],但纬度通过影响水热条件间接影响凋落物分解,充沛的水热条件直接增强淋溶作用并增加微生物活性,促进凋落物分解(P<0.01)[48]。本试验起止时间为7月初至10月末,是小兴安岭地区温度和湿度较高的时间段,因此本试验结果总体上符合凋落叶分解速率的地域性变化趋势。
3.2 掘根倒木腐烂等级和微立地类型以及分解时间对凋落叶养分释放的影响
森林凋落叶分解过程中的养分动态模式有富集-释放和持续释放等模式[49],但不同森林类型和不同地理区域可能有很大差异。本研究中,在Ⅱ、Ⅲ、Ⅳ级掘根倒木腐烂等级条件下,UT、PB、MF微立地的凋落叶C 含量为500.88~728.41 g/kg,整体表现为持续释放到缓慢富集的过程;UT、PB、MF 微立地的凋落叶N 含量为5.76~15.24 g/kg,整体表现为前期缓慢富集而后期持续释放的过程;UT、PB、MF 微立地的凋落叶P 含量为0.77~1.13 g/kg,整体表现为先释放后富集的变化模式,与云南松林、华山松林和常绿阔叶林中凋落物分解过程N、P 变化过程相似[50],与全球植物凋落叶N、P 含量(10.9、0.85 g/kg)[51]相接近。
N、P 作为决定植物生长和微生物矿化有机物质的限制因子,其释放模式受到C∶N 和N∶P 的影响[52]。本试验中UT、PB、MF、FG 和IS 微立地的凋落叶C∶N 分别为57.69、54.03、60.69、69.07 和76.30;N∶P 分别为13.55、11.76、14.19、12.72、和12.11,均高于中国森林生态系统凋落物的C∶N(44.30),均低于中国森林生态系统凋落物的N∶P(25.00)。有研究发现[53-58],在凋落物C∶N>30 和N∶P<14 时,N 对微生物的活动具有限制性;在14<N∶P<16 时,凋落物分解受N、P 共同限制;在N∶P>16 时,凋落物分解受P 的限制性强。数据表明研究区各微立地凋落叶的分解明显受到N养分元素的限制,N 含量不足以满足微生物的活动需求,微生物必须固定外源N,此时需考虑到土壤中的N 的有效性对凋落物分解的影响[59]。
本研究中凋落叶分解90 d 后C∶N 为43.89~127.49,显著高于长白山次生针阔混交林凋落叶的C∶N 值(29.88~69.60)[42],极端高值为FG、IS微立地,其N 释放率均显著大于UT、PB 和MF微立地,且FG>IS,说明在林隙和郁闭林分中椴树红松林凋落叶的N 循环较好,对林木生长可能有一定的促进作用,也反映出林隙干扰对森林生态系统的物质循环具有独特的作用。这种趋势的出现可能是由于倒木形成的坑丘微立地土壤C、P 养分相对丰富,N 含量低于对照,这与张玉双[60]的研究结果一致。也可能是因为凋落物分解需要较长的时间,本试验周期较短且混合凋落叶中50%为红松凋落叶,难分解有机物比例较高,使得研究期间凋落叶质量损失速度与C、N、P 元素的释放速度不同步导致剩余C 的比例较高。本研究中凋落叶的C∶P、N∶P 分别为445.44~902.50、6.73~17.83,与长白山次生针阔混交林[42]凋落叶的C∶P(381.28~867.32)和N∶P(8.62~20.00)相接近。方差分析结果表明微立地类型对凋落叶P 元素释放均产生不同程度的影响,其中MF 微立地凋落叶分解受P 限制,这可能是因为凋落物P元素影响了微生物活动,当P 元素越丰富,分解力强的细菌越多,越有利于凋落物分解[61-62]。
树木风倒掘根干扰为森林生态系统演变提供了重要动力[63],倒木坑丘微立地的形成使森林微生境异质性发生改变,从而造成了凋落物分解速率和养分释放模式等方面的差异,为物种更新创造了条件,可能会丰富森林结构,而倒木坑丘微立地能否提升森林生产力还有待进一步研究。本试验中,随着时间的推移,凋落叶在不同倒木腐烂等级不同微立地的质量分解率、养分动态模式及释放量、养分化学计量比都在随之改变,因此在未来的研究中应采用统一的方法开展更长周期的凋落物分解试验。
3.3 掘根倒木腐烂等级和微立地类型以及分解时间的交互作用
三因素方差分析结果表明,掘根倒木腐烂等级与微立地类型之间的交互作用对凋落叶C 元素释放产生显著影响(P<0.05);其他交互作用对凋落叶分解和养分释放均不产生显著影响(P>0.05),这可能是因为掘根倒木腐烂等级、微立地类型和分解时间之间的交互作用包含了许多生物生理反应[64-65]。凋落叶的分解过程受凋落叶自身性质、环境因素等多重影响[66-67],而这些因素先后或多或少地作用于凋落物分解,所以对凋落叶质量损失、养分释放的作用机理还有待进一步研究。
4 结 论
1)凋落叶分解90 d 后,各掘根倒木腐烂等级的凋落叶分解率依次为Ⅲ级(19.65%)>Ⅳ级(17.27%)>Ⅱ级(15.39%);与对照组相比,UT 微立地的凋落叶分解最快,MF 次之,PB 最慢。
2)Olson 衰减指数模型能够较好地模拟凋落叶的分解动态。比较UT、PB、MF 这3 个微立地类型对分解常数的影响可知,Ⅱ级掘根倒木腐烂等级的分解常数分别为1.19、0.37 和0.90;Ⅲ级掘根倒木腐烂等级的分解常数分别为1.47、0.72 和1.23;Ⅳ级掘根倒木腐烂等级的分解常数分别为1.42、0.45 和1.06;对照组IS 和FG 的分解常数分别为1.23 和1.17;分解50%和95%所需时间分别为0.47~1.90 a 和2.04~8.21 a。
3)各个掘根倒木腐烂等级的凋落叶表现出明显的C、N、P 释放现象,PB 微立地C 释放程度最高,各微立地的N 释放程度相近,Ⅲ级UT 微立地P 释放程度最高。
4)Ⅱ级掘根倒木腐烂等级下UT 微立地的凋落叶质量分解率与N 剩余含量呈显著负相关,与C∶N 呈显著正相关。Ⅱ级掘根倒木腐烂等级下PB微立地与C、N、C∶P 呈显著负相关。Ⅲ级掘根倒木腐烂等级下PB 微立地与N 剩余含量呈极显著负相关,与C∶N 呈极显著正相关。各掘根倒木腐烂等级的MF 微立地的凋落叶质量分解率与各养分剩余含量及其比值均无显著相关关系。而N∶P 与Ⅱ级、Ⅳ级掘根倒木腐烂等级下的UT 和PB 微立地的凋落叶质量分解率呈显著和极显著负相关。
