陆地棉GhCRE1影响种子萌发初探
2022-07-11徐敏慧JoseeOrnellaMusaniwabo韩少凡刘一涵杜丽桦刘爱玉汪启明屠小菊
徐敏慧,Josee Ornella Musaniwabo ,韩少凡,刘一涵,杜丽桦,刘爱玉,汪启明,屠小菊
(1.湖南农业大学 生物科学技术学院,湖南 长沙 410128;2.湖南农业大学 农学院,湖南 长沙 410128)
种子萌发关系着物种的生存和繁衍,细胞分裂素(Cytokinin,CK)等植物激素作为一种小分子信号物质,在浓度极小时仍对种子萌发具有举足轻重的作用[1]。细胞分裂素的作用涉及包括细胞分裂与分化及种子萌发和果实发育等许多方面[2-4],它可通过拮抗ABA信号加快种子萌发[5-7],且这种拮抗作用依赖于细胞分裂素受体CRE1以及下游组氨酸磷酸转移蛋白等[7]。在种子发育的早期阶段,添加一定浓度的细胞分裂素能加速萌发[8],这一时期,细胞分裂素水平暂时升高,还能最终决定种子大小[2,9]。
CRE1可能是表达最强的细胞分裂素受体,它是细胞程序性死亡(Programmed cell death,PCD)所必需的[10-11],大多数已知的CRE1基因突变不会导致较强的植物形态表型[9,12],但会改变生理参数,如磷酸盐饥饿反应和硫酸盐获取的调节[12-13]。Tran等[14]研究发现,拟南芥(Arabidopsisthaliana)CRE1的T-DNA插入突变体种子萌发势稍低于野生型,但是未见陆地棉(GossypiumhirsutumLinn)CRE1基因调控种子萌发的报道。
CRISPR/Cas9技术提供了一种直接的方法,可以精确设计基因组中所需的突变,且可筛选到不含外源基因的突变体,已成为基因功能研究的重要工具[15]。Wang等[16]利用CRISPR/Cas9技术对西瓜(Citrulluslanatus)Clbg1基因进行了靶向突变,发现Clbg1功能缺失型突变体萌发率显著提高。Zhou等[17]利用CRISPR/Cas9技术构建水稻(Oryzasativa)Os05circ02465基因功能缺失型突变体,发现其种子萌发后生长速度加快。Shi等[18]利用CRISPR/Cas9技术敲除烟草(Nicotianatabacum)NtCIPK23基因,发现敲除突变体显著延迟了种子萌发时间。
本研究以陆地棉细胞分裂素受体基因GhCRE1为研究对象,通过克隆该基因,构建棉花GhCRE1过表达载体,转化拟南芥获得过表达转基因株系。采用最新的CRISPR/Cas9 基因编辑技术获得拟南芥CRE1功能缺失突变体,通过对过表达株系及功能缺失株系的表型观察,解析GhCRE1基因在调控植物生理过程中的功能,旨在为进一步研究陆地棉GhCRE1基因的功能提供植物材料,为明确植物萌发的分子机制奠定基础。
1 材料和方法
1.1 试验材料
供试棉花品种为陆地棉矮化突变体陆矮1号(LA-1),由湖南农业大学棉花研究所提供。大肠杆菌菌株DH5α和农杆菌菌株GV3101均购自长沙擎科生物技术有限公司。载体pYLCRISPR/Cas9Pubi-B、pYLsgRNA-AtU6-1、pYLsgRNA-AtU3d-LacZ、pCUbi1390由华南农业大学刘耀光院士实验室赠送,拟南芥野生型Col-0由湖南农业大学道地药用植物规范化栽培与利用湖南省工程实验室赠送。
1.2 试验方法
1.2.1 陆地棉GhCRE1基因的克隆 使用TRIzol提取法获得纯度较高的陆地棉总RNA,反转录获得全cDNA。利用拟南芥中的CRE1基因序列,在NCBI数据库(www.ncbi.nlm.nih.gov)及COTTONGEN数据库(www.cottongen.org)中进行同源比对,选取2个数据库中相同序列信息,使用Primer Premier 5.0设计并在NCBI中验证获取特异性引物序列GhCRE1-F/R。以cDNA为模板,以GhCRE1-F/R为引物进行PCR反应扩增GhCRE1全长,引物序列为GhCRE1-F:5′-TACCCAGAATTCAACGTTGTCTCG-3′,GhCRE1-R:5′-CTATGAGTCCGAGATGGGTTTCG-3′,反应产物通过1%琼脂糖凝胶进行电泳检测;产物回收纯化后,连接T clone载体转化大肠杆菌进行菌落PCR,选择阳性克隆测序,确定获得GhCRE1基因cDNA序列具有完整的开放阅读框。
1.2.2 重组过表达载体的构建 将pCUbi1390载体用PstⅠ酶切,以含有目的基因重组T clone载体质粒为模板进行PCR扩增,PCR产物纯化后与线性表达载体进行无缝克隆(图1)。将无缝克隆后的表达载体转化到大肠杆菌,挑选单克隆菌落进行PCR,选择阳性单克隆提取质粒,采用酶切及测序的方法进行验证。验证引物序列为:GhCRE1-CDS-F:5′-GCTTTGGGTAATGGTGATG-3′,GhCRE1-CDS-R:5′-CAATAAGCAGATCACGAAGAATAAG-3′。
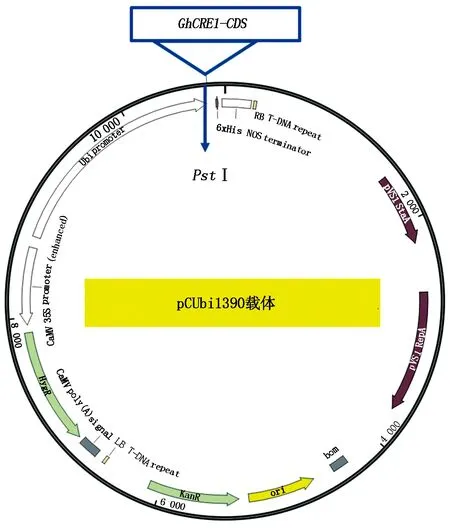
图1 重组GhCRE1过表达载体构建方法示意Fig.1 Schematic diagram of construction method of recombinant GhCRE1 overexpression vector
1.2.3 拟南芥GhCRE1基因过表达株系的筛选 构建的过表达载体用冻融法转化农杆菌GV3101,随后用浸花法侵染花蕾期的拟南芥Col-0,收获T0种子。T1潮霉素筛选具有抗性的植株,利用验证引物GhCRE1-CDS-F/R进行PCR检测,得到阳性植株,同样的方法筛选至纯合子。以Actin11为内参,设计qPCR引物GhCRE1-qF/R,检测GhCRE1基因的表达量,筛选表达量高的株系进行后续试验,引物序列如表1。

表1 拟南芥GhCRE1过表达材料的筛选与鉴定引物Tab.1 Primers used in screening and identification of Arabidopsis GhCRE1 overexpression plants
1.2.4 拟南芥CRE1靶向敲除载体构建 sgRNA靶向确定和引物设计:对拟南芥CRE1基因的ORF的5′区及功能区均设计一个靶位点,以提高突变效率。利用软件CRISPR-P设计拟南芥CRE1靶位点分别为:靶位点1:ATCCTCTCACAACTCATTAC位于5′UTR区,GC含量40%,选用AtU3d-LacZ作为启动子;靶位点2:GCTGCTGCGTTTGAAAGAAA位于外显子区域,GC含量45%,选用 AtU6-1作为启动子。采用Overlapping PCR的方法分别将2个靶位点序列引入PCR正向及反向引物中,并引入依次排列的BsaⅠ酶切位点至引物5′端,获得所需 sgRNA。相应的引物如表2。

表2 sgRNA表达盒引物Tab.2 Primers used in construction of sgRNA expression cassette
CRISPR/Cas9基因敲除载体的构建:使用限制性内切酶BsaⅠ酶切pYLCRISPR/Cas9质粒,酶切产物经电泳后切胶回收。采用Golden Gate cloning(NEB)的方法,利用T4连接酶将pYLCRISPR/Cas9线性化载体分别与CRE1的2个sgRNA表达盒片段组装。将重组载体转入大肠杆菌DH5α,使用含相应抗生素的LB培养基培养,菌落PCR筛选阳性菌。利用各靶点引物特异性,对阳性菌进行靶位点测序,采用的引物为SP-L与SP-R,序列如下:SP-L:GCGGTGTCATCTATGTTACTAG,SP-R:CCCGACATAGATGCAATAACTTC。
1.2.5 拟南芥CRE1基因编辑植株的获得及筛选 将构建的敲除载体利用冻融法转化农杆菌GV3101,用浸花法侵染花蕾期的野生型拟南芥Col-0。收集得到T0种子。使用引物CRE1T1-F/R和CRE1T2-F/R对T0植株分别进行PCR扩增,获得的扩增片段切胶回收后测序检测,测序结果显示,在靶位点处为套峰的,对其相应的植株进行单株收种并检测编辑效果。逐代筛选后,使用CRE1功能缺失型突变体纯合子进行后续试验,测序引物见表3。
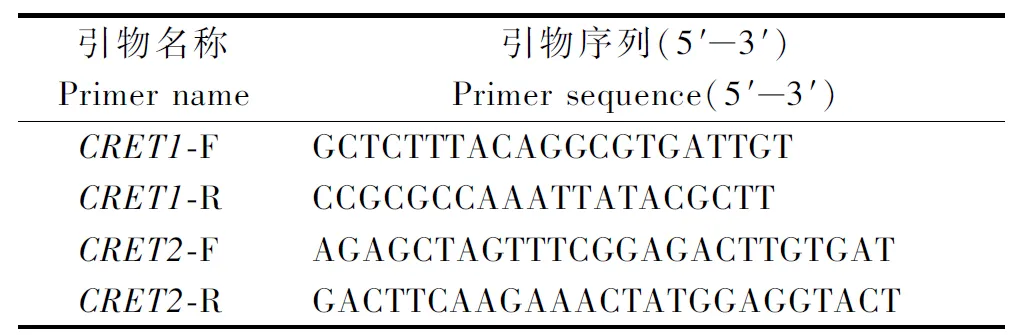
表3 拟南芥CRE1基因编辑植株的筛选与鉴定Tab.3 Primers used in screening and identification of of CRE1 gene editing plants
1.2.6 种子萌发试验 将筛选得到的GhCRE1过表达转基因株系、拟南芥CRE1基因编辑突变体及野生型Col-0 各50颗点种于1/2 MS固体培养基上,并在4 ℃下春化3 d后转入22 ℃ 16 h光照/8 h黑暗条件下培养,每天固定时间检测种子萌发势,每个处理3个重复。数据均采用SPSS 26.0软件进行统计分析,并用Origin 2019b作图。
2 结果与分析
2.1 陆地棉GhCRE1基因过表达转基因株系构建
2.1.1GhCRE1基因克隆 以拟南芥CRE1基因序列为模板,Blast比对得到陆地棉该基因序列,以陆地棉叶片为材料,液氮研磨后提取RNA,并反转录为cDNA。以陆地棉cDNA为模板,以GhCRE1-F及GhCRE1-R为引物进行PCR扩增得到GhCRE1基因编码序列的CDS片段。经琼脂糖凝胶电泳鉴定,结果显示(图2),PCR扩增得到3 000 bp左右条带,将扩增产物纯化后测序发现,克隆的基因长度为3 012 bp,克隆得到的条带与目的条带一致,结果表明,成功克隆了GhCRE1-CDS片段。

M.DNA Marker DL1000;1,2.GhCRE1-CDS片段。M.DNA Marker DL1000;1,2. GhCRE1-CDS segment.
2.1.2GhCRE1基因过表达载体的构建 根据GhCRE1序列,利用Primer Premier 5.0软件设计引物验证无缝克隆后的过表达载体阳性单克隆,如图3-A所示,将重组载体转化大肠杆菌(Escherichiacoli)后,用上述检测引物进行菌落PCR,所有菌落均扩增获得长913 bp的条带,将重组质粒用PstⅠ酶切后获得3 012 bp的条带(图3-B),测序结果显示,GhCRE1-CDS已插入pCUbi1390过表达载体,且无碱基突变,以上结果均符合预期,表明GhCRE1重组过表达载体构建成功。
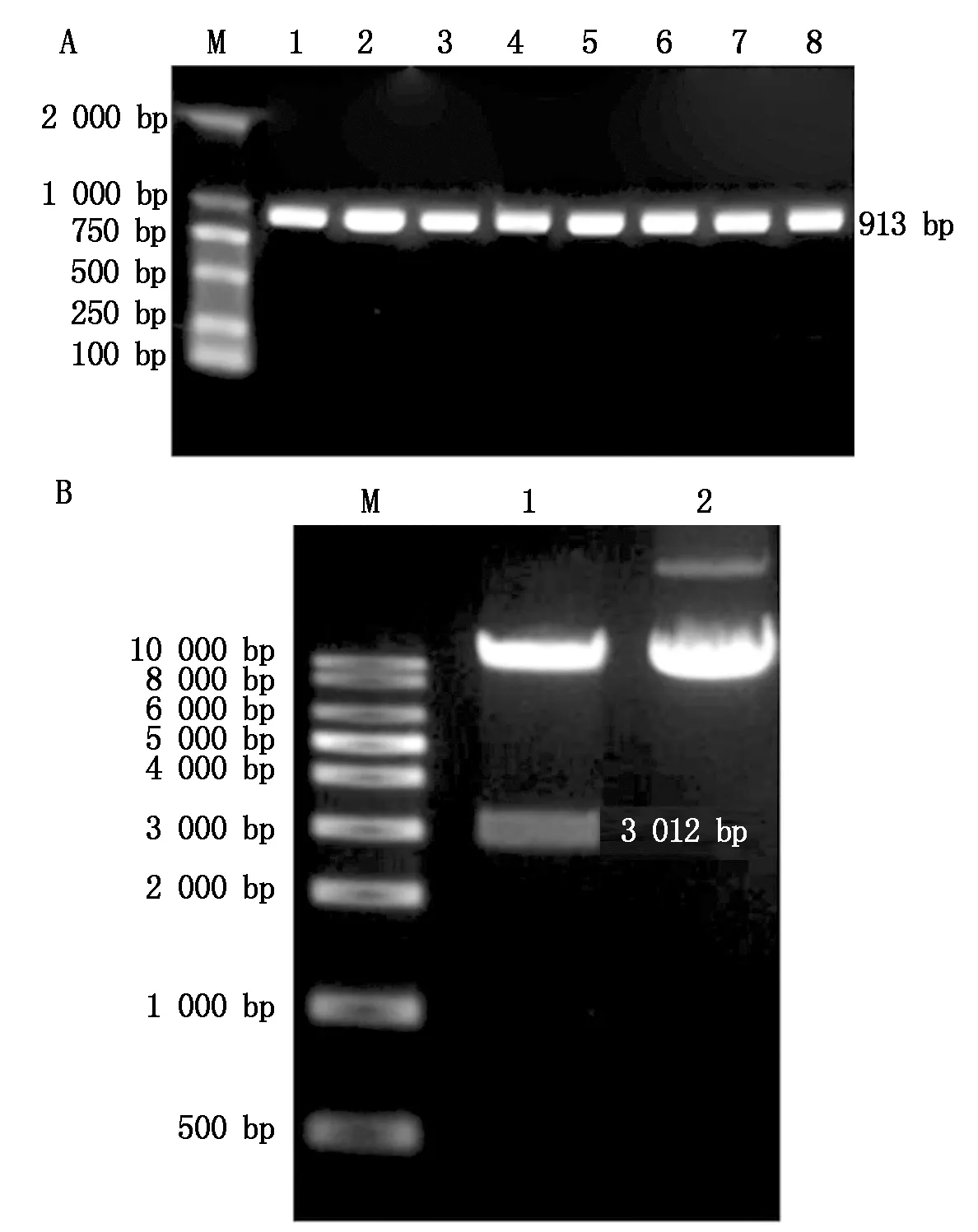
A.重组载体菌落PCR:M.DNA Marker DL2000;1—8.菌落PCR检测。B.重组质粒电泳及Pst Ⅰ酶切重组质粒:M.DNA Marker DL10000;1.Pst Ⅰ酶切重组质粒;2.重组质粒。
2.1.3 阳性拟南芥GhCRE1过表达材料的获得及筛选 将得到的重组质粒使用农杆菌介导法转化拟南芥Col-0,潮霉素筛选后获得的T0植株,经选取新鲜叶片提取DNA及引物GhCRE1-CDS-F/R序列扩增,共获得7个阳性株系(图4-A)。以Actin11为内参,进行qPCR分析后,选择GhCRE1相对表达量高的株系GhCRE1 5-1 与GhCRE1 9-2进行后续试验(图4-B)。
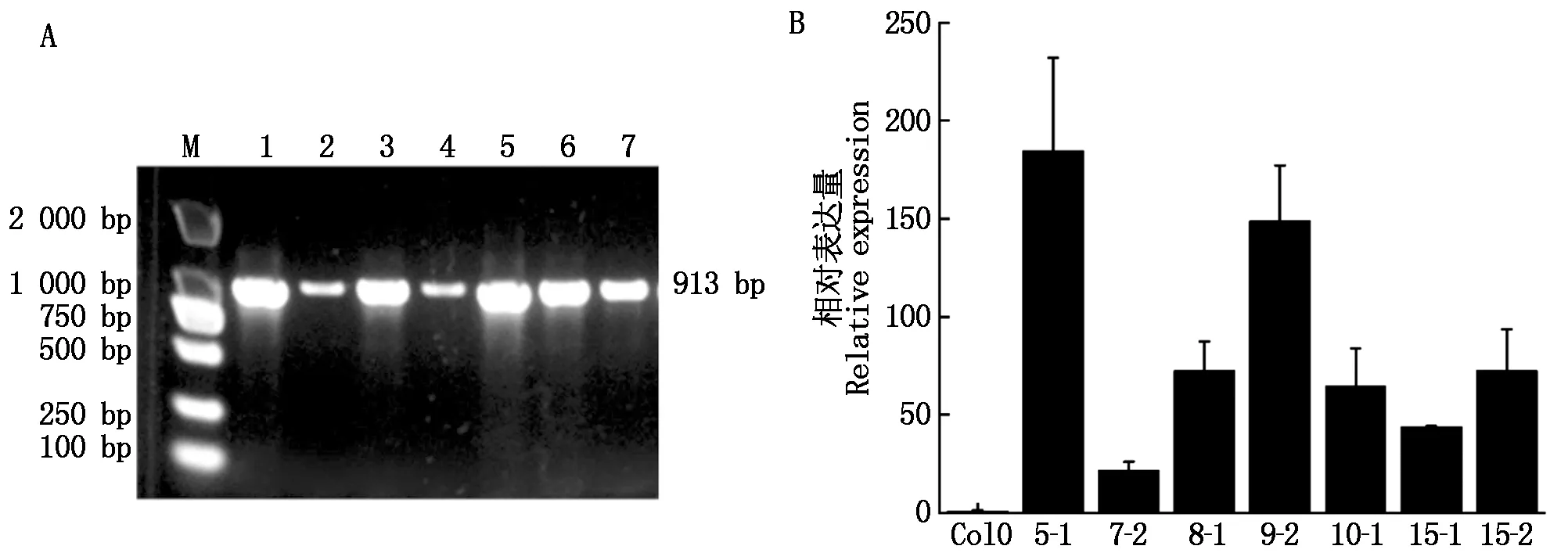
A.T0植株PCR检测:M.DNA Marker DL2000;1—7.GhCRE1 过表达检测PCR电泳条带。B.qPCR检测GhCRE1相对表达量。A.PCR assay of T0 generation plants:M.DNA Marker DL2000;1—7.PCR electrophoresis bands for GhCRE1 overexpression detection.B.qPCR assay of GhCRE1 relative expression.
2.2 拟南芥CRE1靶向敲除载体材料构建
2.2.1 基因编辑靶位点的选择及敲除载体的构建 根据CRISPR-P软件分析及CRE1基因序列分析,设计sgRNA序列分别为sgRNA-tg1:ATCCTCTCACAACTCATTACAGG(划线部分为靶位点1);sgRNA-tg2:GCTGCTGCGTTTGAAAGAAACGG(划线部分为靶位点2)。靶位点1位于5′UTR区,选用AtU3d-LacZ作为启动子,靶位点2位于外显子区,选用 AtU6-1作为启动子。
分别将靶位点1和靶位点2通过Overlapping PCR的方法与PYLsgRNA-AtU6-1、pYLsgRNA-AtU3d-LacZ中间载体连接。第一轮Overlapping PCR(图5-A),出现3条带,所需的条带为453 bp,泳道3和4分别为CRE12个靶位点的特异性产物。第二轮Overlapping PCR过后(图5-B),453 bp的条带更清晰。随后PCR增加BsaⅠ 酶切位点(图5-C),将重组后的2个中间载体分别连接T clone载体,之后转大肠杆菌,菌落PCR扩增结果见图5-D,表明重组中间载体构建成功。

M.DNA Marker DL2000;A.第一轮Overlapping PCR;B.第二轮Overlapping PCR:泳道3和4分别为CRE1-tg1和CRE1-tg2;C.增加Bsa Ⅰ酶切位点后跑胶;D.菌落PCR。M.DNA Marker DL2000;A.First round of Overlapping PCR;B.Second round of Overlapping PCR:lanes 3 and4 are CRE1-tg1 and CRE1-tg2;C.Running gel after addition of the Bsa Ⅰ digestion site;D.Colony PCR.
BsaⅠ酶切终载体pYLCRISPR/Cas9Pubi-B(图6-A),重组中间载体连接后转化大肠杆菌,利用SP-L及SP-R这一对检测引物进行PCR扩增,产生与预计大小一致的目的片段(图6-B),可见拟南芥CRE1双靶点敲除载体初步构建成功,将该载体命名为PYLCas9-CRE1-T1/T2。

M.DNA Marker DL2000;A.BsaⅠ酶切pYLCRISPR/Cas9;B.菌落PCR。M.DNA Marker DL2000;A.pYLCRISPR/Cas9 was digestion by BsaⅠ;B.Colony PCR.
载体PYLCas9-CRE1-T1/T2经序列比对发现,sgRNA已插入pYLCRISPR/Cas9Pubi-B线性化基因编辑载体,且无碱基突变,表明靶向CRE1基因的CRISPR/Cas9基因编辑载体构建成功(图7)。
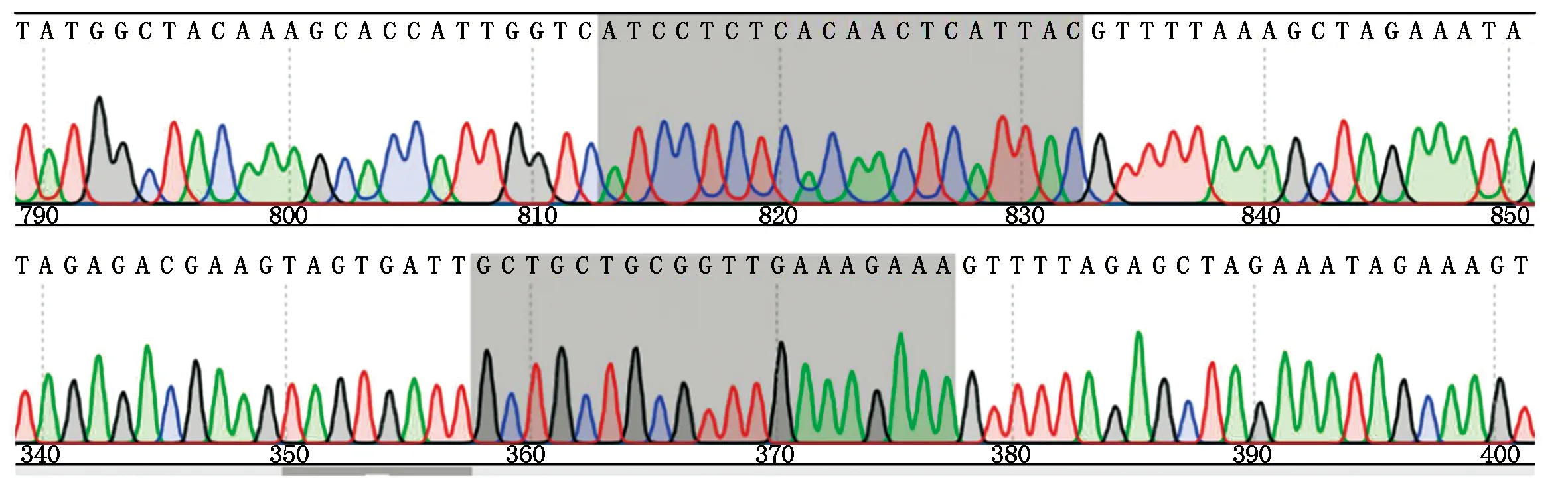
序列标记的阴影矩形部分为靶位点DNA序列(上为靶位点1,下为靶位点2)。The target DNA sequence is labeled with shadow rectangles(the top is target site 1,bottom is target site 2).
2.2.2 拟南芥CRE1靶向敲除材料的构建 T0植株经除草剂初筛后共获得27株生长良好的株系,经测序后发现4个株系(CRE1 T0-14、CRE1 T0-18、CRE1 T0-22、CRE1 T0-27)在靶位点1处测序结果为叠峰,说明这4个株系在靶位点1处发生了突变。经生物信息学分析得知,这4个株系均在靶位点处缺失了1个碱基T(图8),导致转录提前终止,造成拟南芥CRE1的功能缺失型突变。因为这4个株系编辑效果一致,筛选到纯合子后,采用cre127-1进行后续种子萌发试验。

红色标志序列部分ATCCTCTCACAACTCATTAC为靶位点1。The red marker sequence part ATCCTCTCACAACTCATTAC is the target site 1.
2.3 陆地棉GhCRE1基因影响拟南芥种子萌发
如图9所示,在1/2 MS 培养基中,相比野生型,cre127-1萌发率在第1天低9%左右,第2,3天低15%左右差异均达到显著水平(P<0.05);与野生型相比,GhCRE1 5-1与GhCRE1 9-2第2天萌发率分别高近15%与11%,差异达到显著水平(P<0.05)。因此,拟南芥CRE1功能缺失会导致种子萌发速度明显减慢,而陆地棉GhCRE1基因在拟南芥中过表达能显著加快种子萌发速度。

A.CRE1基因各突变体萌发势;B.CRE1基因各突变体种子萌发表型。A.Germination potential of each CRE1 mutant;B.Seed germination phenotype of each CRE1 mutant.
3 结论与讨论
作为植物必需的激素之一,细胞分裂素能调节诸如细胞分裂、组织和器官以及个体的生长发育、营养吸收等过程[19]。组氨酸激酶(CRE1)作为细胞分裂素的重要受体,对细胞分裂素信号转导途径中信息的传递具有重要的调节作用。但是,CRE1在调控除拟南芥之外的包括陆地棉等物种的种子萌发中的功能还未见相关的报道。因此,本试验研究陆地棉GhCRE1基因在种子萌发中的功能具有重要的意义。
本研究首次成功克隆了陆地棉GhCRE1基因,并获得GhCRE1的过表达拟南芥植株和CRE1基因编辑拟南芥。通过表型验证发现,CRE1对启动种子萌发具有正调控作用,为进一步研究陆地棉GhCRE1基因的功能提供植物材料,为明确植物萌发的分子机理奠定基础。
值得注意的是,湖南农业大学棉花研究所前期针对目前陆地棉矮秆种质资源匮乏的问题,通过对陆地棉矮化突变体陆矮1号(LA-1)和高秆近等基因系(LH-1)的研究,筛选得到陆地棉细胞分裂素受体基因(CRE1)与LA-1的矮化密切相关。但本试验经过对陆地棉GhCRE1过表达拟南芥成苗株高的测量,未观测到矮化现象,且在正常生长条件下,未观察到GhCRE1过表达拟南芥、CRE1基因编辑拟南芥在其他生长发育方面的差别,说明GhCRE1可能不参与调控拟南芥株高等生长发育的表型。
种子萌发受复杂的信号网络和基因表达调控,不同植物可能具有相似的分子机制,包括植物激素的调节、环境因素等[20]。湖南农业大学棉花研究所在培养基中添加不同浓度的外源脱落酸(Abscisic acid,ABA)处理本研究获得的拟南芥GhCRE1过表达种子及基因编辑突变体拟南芥cre127-1种子,结果发现,在外源ABA存在下,GhCRE1过表达种子比野生型萌发速度更慢,而突变体萌发速度相比野生型更快。众所周知,细胞分裂素与脱落酸具有拮抗作用[21],有研究发现,CK缺乏增加了植物对外源ABA的敏感性,但它也能导致ABA生物合成关键基因的下调,从而使CK缺乏植株的内源ABA,水平明显低于野生型[22]。因此,推测GhCRE1过表达拟南芥对外源ABA更敏感,而CRE1缺失突变体通过降低内源ABA的合成从而影响萌发势。本研究发现,外源ABA和CRE1能共同调控种子萌发,接下来会针对其调控的具体分子机制进行进一步的研究。这一发现有助于进一步了解脱落酸和细胞分裂素的拮抗作用发生机制。后期本课题组会将该基因转入陆地棉中,进一步验证该基因在调控棉花种子萌发及其他生理过程中的功能。
