水稻黄叶不育系突变体H08S的表型特征与遗传分析
2020-09-02林秋云谢振宇龙开意贺治洲
林秋云 谢振宇 龙开意 贺治洲
摘 要:光温敏不育系的育性因易受光温条件影响而出现波动,导致种子纯度不够,是水稻生产上常见的问题。本研究对1个水稻黄叶不育系突变体H08S进行温度敏感性分析和叶绿体超微结构观察,同时通过构建分离群体进行突变基因的遗传模式分析。结果表明,突变体H08S的表型受温度影响,为低温表达型叶色突变体,且其叶绿体结构出现异常,表现类囊体的片层结构减少,说明H08S基因的突变影响叶绿体的正常发育。突变体H08S分别与‘日本晴、‘02428构建F2群体和BC1F1群体,观察并统计群体植株叶色表型的分离情况,并进行卡方检验,结果表明该突变性状受1对隐性单基因控制。
关键词:水稻;黄叶不育系突变体;H08S;表型;遗传中图分类号:S511 文献标识码:A
Phenotypic Characteristics and Genetic Analysis of a Rice PTGMS Line Mutant H08S with Yellow Leaf
LIN Qiuyun, XIE Zhenyu, LONG Kaiyi, HE Zhizhou*
Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan 571101, China
Abstract: PTGMS lines are susceptible to light and temperature conditions during their fertility sensitive period, which results in fertility fluctuation and seriously affects the purity of the original or hybrid seeds of the male sterile lines. In this paper, the temperature sensitivity and chloroplast ultrastructure of a rice PTGMS mutantH08S with yellow leaf were analyzed, and the genetic model of the mutant gene was analyzed by constructing two segregated populations. The results showed that the phenotype of mutantH08Swas affected by temperature, and it was a low temperature expression type of leaf color mutant. The chloroplast structure of mutantH08S was abnormal, and the lamellar structure of thylakoid was reduced. This indicated that the mutation ofH08S gene affected the normal development of chloroplast. F2 population and BC1F1 population were constructed with mutantH08Sand ‘Nipponbare and ‘02428, respectively. The isolation of leaf color phenotypes was observed and counted, and chi-square test was carried out. The results showed that the mutant trait was controlled by a pair of recessive single genes.
Keywords: rice; a rice PTGMS line mutant with yellow leaf;H08S; phenotype; heredity
DOI: 10.3969/j.issn.1000-2561.2020.07.006
葉色突变体绝大部分都表现出叶绿素含量降低,而且多数会影响光合效率,有些叶色突变体甚至不能存活,有些叶色突变体即便能结实,其产量往往会显著降低,因此在过去很长一段时间都被认为是无意义的突变。随着水稻功能基因组学的快速发展,大量研究表明叶色突变体不仅是开展光合作用、光形态建成、叶绿素合成和叶绿体发育等基础研究的理想材料[1],而且其变异叶色表型可作为一种明显且易于识别的遗传形态标记,在育种上简化杂交稻种子纯度鉴定工作,很大程度上减少了种子纯度鉴定时间[2-3]。
‘深08S是目前生产上应用较广的水稻光温敏两用核不育系,其不仅保留了‘Y58S抗性好、配合力高的特点,还增强了抗稻瘟病、抗倒性、抗逆性等特性,已成为杂交水稻的重要骨干亲本。黄叶不育系突变体H08S是沈建凯等[4]在海南种植‘深08S时发现的1个自然突变体。沈建凯等[4]为了研究突变体H08S的配合力,将‘02428等共15个品种作为父本,分别与突变体H08S进行杂交获得杂交种子,对F1代进行产量特性分析,结果发现配组的杂交品种的产量优势较好,增产组合品种率高,说明突变体H08S的杂交组合优势较强。由于H08S携带黄叶标记,所以比野生型‘深08S更具有生产应用价值。本研究拟对突变体H08S的表型性状、细胞学特性和遗传模式进行分析,以期为挖掘新的水稻叶色基因和促进H08S叶色突变基因在水稻育种、种子生产和种子纯度鉴定上的应用提供参考。
1 材料与方法
1.1 材料
突变体H08S是光温敏两用核不育系‘深08S的一个黄叶自然突变体。2017年7月将‘深08S和突变体H08S种植于中国热带农业科学院热带作物品种资源研究所海南省儋州市水稻综合试验基地,水稻种植行距20 cm,株距13.3 cm,每穴1苗,田间水肥病虫管理同常规大田,种子完全成熟后收获并晒干保存备用。
1.2 方法
1.2.1 温敏试验处理前浸种催芽 将‘深08S和突变体H08S种子在室温下浸种2 d,随后置于智能型光照培养箱(武汉瑞华仪器设备有限责任公司,型号HP400G-C)中设定30 ℃条件下催芽0.5 d。
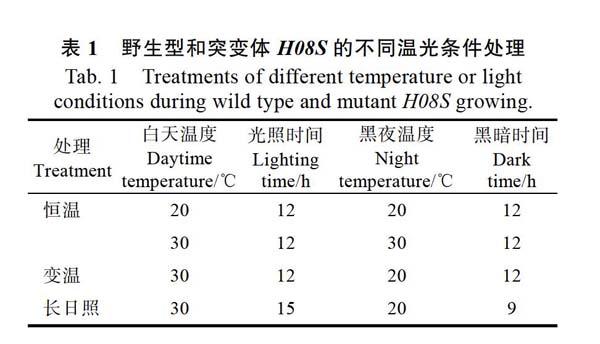
1.2.2 不同生长温度的处理 将催芽后的‘深08S和突变体H08S种子种植在光照培养箱中[光照强度为160 ?mol/(m2·s),空气相对湿度为85%],分别设定恒温、变温和长日照生长条件(见表1),生长2周后进行表型观察和色素含量测定。
1.2.3 光合色素含量测定 ‘深08S和突变体H08S在不同恒温条件下生长2周后,分别对第1不完全叶和第2叶进行光合色素含量测定,测定方法参照Li等[5]的方法。将所取叶片剪成约2~
3 mm的碎片,称量剪碎的新鲜样品约20~30 mg,重复3次,加入5 mL的95%乙醇后置于室温黑暗中避光放置48 h,期间多次混匀,直到叶片完全发白。提取完毕,用DU800紫外-可见分光光度计在黑暗条件下测定波长为470、649、665 nm下上清液的吸光值。根据以下公式计算色素浓度:
叶绿素a浓度Ca= 13.95D665?6.88D649
叶绿素b浓度Cb= 24.96D649?7.32D665
胡萝卜素浓度Cx=(1000D470?2.05Ca?114Cb)/ 245
再依据以下公式换算出组织中各细胞色素的含量:
细胞色素的含量(mg/g)=(叶绿素的浓度×提取液体积×稀释倍数)/样品鲜重
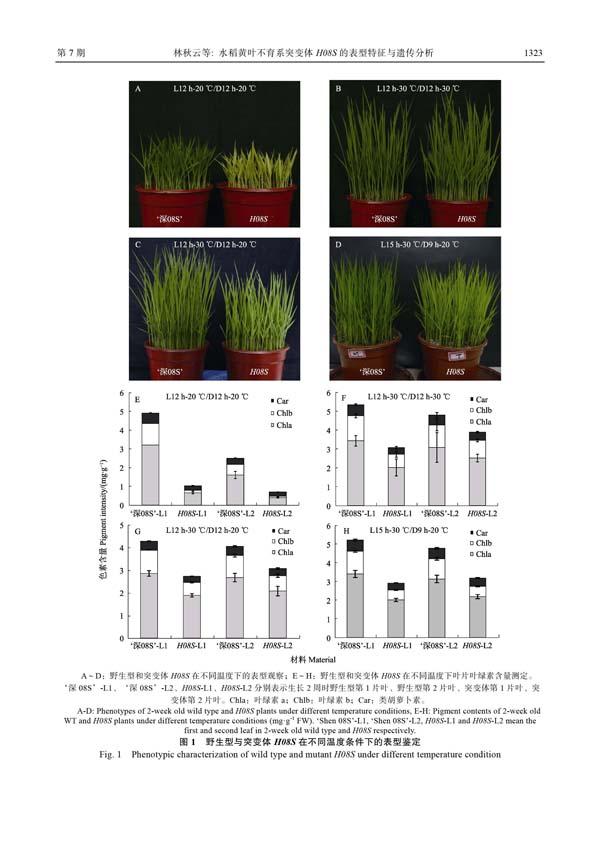
1.2.4 叶绿体超微结构的透射电镜观察 分别取20 ℃恒温条件下生长2周的‘深08S和突变体H08S的第2叶,横切成数段,每段约2 mm,置于3%的戊二醛固定液,抽气,室温固定12 h,4 ℃保存。将样品送至中国热带农业科学院环境与植物保护研究所电镜实验中心进行样品制备和透射电镜观察。
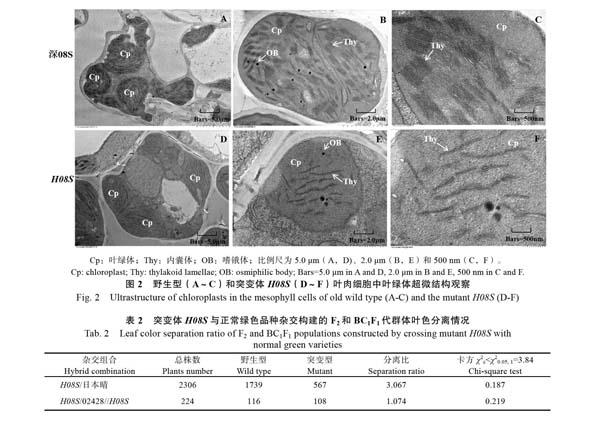
1.2.5 突變体H08S的遗传模式分析 将突变体H08S与‘日本晴杂交获得F1,并自交获得F2,将突变体H08S与‘02428杂交并回交获得BC1F1。将催芽后的‘深08S、突变体H08S和F1、F2、BC1F1群体的种子种植在光照培养箱中,在恒温20 ℃等昼夜生长条件下生长2周后,观察并统计F1、F2、BC1F1群体的叶色表型和分离情况,并进行卡方测验,分析突变体H08S的遗传模式。
2 结果与分析
2.1突变体H08S的温度敏感性分析
‘深08S和突变体H08S分别在等昼夜长的恒温20、30 ℃条件下生长2周(图1),突变体
A~D:野生型和突变体H08S在不同温度下的表型观察;E~H:野生型和突变体H08S在不同温度下叶片叶绿素含量测定。
‘深08S-L1、‘深08S-L2、H08S-L1、H08S-L2分别表示生长2周时野生型第1片叶、野生型第2片叶、突变体第1片叶、突变体第2片叶。Chla:叶绿素a;Chlb:叶绿素b;Car:类胡萝卜素。
A-D: Phenotypes of 2-week old wild type andH08Splants under different temperature conditions, E-H: Pigment contents of 2-week old WT andH08S plants under different temperature conditions (mg·g-1FW). ‘Shen 08S-L1, ‘Shen 08S-L2,H08S-L1 andH08S-L2 mean the first and second leaf in 2-week old wild type andH08Srespectively.
H08S在20 ℃条件下叶片黄化表型明显,光合色素含量明显降低(图1A、E),而在30 ℃下肉眼观察‘深08S与突变体H08S的叶片叶色几乎无差异(图1B)。将‘深08S和突变体H08S在变温条件下(光照12 h-30 ℃/黑暗12 h-20 ℃)生长2周,结果如图1C和图1G所示,突变体H08S的叶片黄化程度减弱,而延长光照时间(光照15 h-30 ℃/黑暗9 h-20 ℃),结果如图1D和图1H所示,肉眼观察突变体H08S的叶片黄化程度不明显,其接近野生型的表型。这些结果表明,突变体H08S的表型受温度影响,为低温表达型叶色突变体。
2.2 突变体H08S的叶绿体超微结构观察
突变体H08S在20 ℃生长条件下表现出极为明显的黄化表型和叶绿素含量减少。为了了解该基因突变是否影响到叶绿体的发育,本研究通过透射电镜观察突变体H08S第2叶黄化叶片中叶绿体的超微结构。如图2A与图2D所示,突变体H08S的叶绿体数目与野生型相比无显著差异。然而突变体H08S的叶绿体结构出现异常(图2B与图2E),与野生型的叶绿体结构相比,其类囊体的片层结构显著减少(图2C与图2F),说明突变体H08S的叶绿体发育不正常,H08S基因的突变影响其叶绿体的正常发育。
2.3 突变体H08S的遗传模式分析
突变体H08S与‘日本晴杂交获得的F1代植株叶片均表现正常绿色,而自交后的F2代群体分离出叶色差异明显的正常绿苗和黄化苗。突变体H08S与‘02428杂交后F1代植株叶片也都表现正常绿色苗,测交后代BC1F1分离出现正常绿苗和黄化苗。卡方测验结果表明,F2代群体中绿苗与黄化苗的分离比例符合3∶1,BC1F1群体中绿苗与黄化苗的分离比例符合1∶1(表2),说明该突变性状受1对隐性单基因控制。
Cp:叶绿体;Thy:内囊体;OB:嗜锇体;比例尺为5.0 μm(A,D)、2.0 μm(B,E)和500 nm(C,F)。
Cp: chloroplast; Thy: thylakoid lamellae; OB: osmiphilic body; Bars=5.0 μm in A and D, 2.0 μm in B and E, 500 nm in C and F.
3 讨论
目前已发现的叶色突变体的表型非常丰富,很难找到完善的分类方法来区分所有的叶色突变体。若从叶色标记表达受温度影响程度来区分,则叶色突变体分为低温表达型、高温表达型和温钝型3种[6]。其中,大多数属于低温表达型,如已报道的水稻叶色突变体v1、v2、W1、W17、W25 和tsc-1以及本研究中的突变体H08S,在低温20 ℃条件下均出现显著的不正常叶色表型,而当生长温度提升到30 ℃时,这些突变体的叶色转为浅绿或正常绿色[7-9]。此外,已有研究表明,叶色突变体表型还受不同光强、不同光质或光照时间的影响,因此叶色突变体又可分为依赖于光诱导型和非依赖于光诱导型[10]。
大部分水稻叶色突变性状受1对隐性核基因控制,而由显性基因控制或是细胞质基因控制的叶色突变体非常少见[11]。据不完全统计,目前已有超过130个水稻叶色突变基因被定位[12],遍及水稻基因组12条染色体。遗传分析表明突变体H08S受1对隐性单基因控制,其候选基因仍有待下一步研究。
近年来,育种家越来越关注叶色突变体的叶色变异表型在育种上的应用。目前成功地实现叶色标记在不育系上的应用分有2种类型,一是全生育期均出现变异叶色表型;二是变异叶色表型只在生育期中某一阶段表达。不育系H08S携带黄化叶色突变表型,且其作为两系不育系,配组自由,比三系法更容易培育出抗性更好、产量更高和品质更优的杂交水稻组合,对优质高产杂交水稻新品种的选育具有重要的应用价值。
参考文献
- Fambrini M, Castagna A, Dalla V F, et al. Characterization of a pigment-deficient mutant of sunflower (Helianthus annuus L.) with abnormal chloroplast biogenesis, reduced PS II
activity and low endogenous level of abscisic acid[J]. Plant science, 2004, 167(1): 79-89.
- Su N, Hu M L, Wu D X, et al. Disruption of a rice pentatricopeptide repeat protein causes a seedling-specific albino phenotype and its utilization to enhance seed purity in hybrid rice production[J]. Plant physiology, 2012, 159(1): 227-238.
- Zhao Y, Wang M L, Zhang Y Z, et al. A chlorophyll-reduced seedling mutant in oilseed rape, Brassica napus, for utilization in F1 hybrid production[J]. Plane Breeding, 2000, 119(2): 131-135.
- 沈建凯, 林秋云, 贺治洲, 等. 水稻叶色黄化不育系H08S杂交组合产量分析[J]. 热带农业科学, 2019, 39(2): 29-32.
- Li J Q, Wang Y H, Chai J T, et al. Green-revertible Chlorina 1 (grc1) is required for the biosynthesis of chlorophyll and the early development of chloroplasts in rice[J]. Journal of Plant Biology, 2013, 56(5): 326-335.
- 景晓阳, 吴殿星, 舒庆尧, 等. 60Co-γ射线诱发的籼型温敏核不育水稻叶色突变系变异分析[J]. 作物学报, 1999, 25(1): 64-69.
- Kusumi K, Sakata C, Nakamura T, et al. A plastid protein NUS1 is essential for build-up of the genetic system for early chloroplast development under cold stress conditions[J]. The Plant Journal: for Cell and Molecular Biology, 2011, 68(6): 1039-1050.
- Sugimoto H, Kusumi K, Noguchi K, et al. The rice nuclear gene, VIRESCENT 2, is essential for chloroplast development and encodes a novel type of guanylate kinase targeted to plastids and mitochondria[J]. The Plant Journal: for Cell and Molecular Biology, 2007, 52(3): 512-527.
- Sugimoto H, Kusumi K, Tozawa Y, et al. The virescent-2 mutation inhibits translation of plastid transcripts for the plastid genetic system at an early stage of chloroplast differentiation[J]. Plant and Cell Physiology, 2004, 45(8): 985-996.
- Kusumi K, Komori H, Satoh H, et al. Characterization of a zebra mutant of rice with increased susceptibility to light stress [J]. Plant Cell Physiology, 2000, 41(2): 158-164.
- 魏彥林, 施勇烽, 吴建利. 水稻核基因控制的叶色变异[J]. 核农学报, 2011, 25(6): 1169-1178.
- 邓晓娟, 张海清, 王 悦, 等. 水稻叶色突变基因研究进展[J]. 杂交水稻, 2012 , 27(5): 9-14, 30.
