河川沙塘鳢对猎物种类和大小选择性研究
2022-07-08刘怡然谢德兵胡少迪史阳白朱思华
任 艳 马 琴 刘怡然 谢德兵 胡少迪 王 辉 史阳白 李 波 朱思华
(1.武汉市农业科学院,武汉 430070;2.南昌师范学院,南昌 330006;3.江苏省淡水水产研究所,南京 210000)
肉食性鱼类对猎物种类和规格具有明显的选择性,多数肉食性鱼类的捕食活动受其口裂的限制[1—3],口裂从根本上限制猎物规格和类型的选择[4]。不同猎物的体型、防御结构、游泳速度和反捕食能力等不同,因而其存活能力也存在差异[2,5,6];捕食者多捕获纺锤形或圆柱形、软鳍条、无硬棘的饵料鱼[7,8]。
捕食者与猎物之间的大小关系是捕食者对猎物选择的基础[4,9]。多数肉食性鱼类在捕食过程中吞咽整个猎物[4],环境中猎物的规格范围限制了捕食者的捕食[10],超过捕食者口裂大小的猎物不易被其完全吞食,常咬伤、放弃或咬断猎物,造成饵料资源的浪费和捕食者能量的损耗。对于在捕食者口裂范围内的猎物,其规格过大则易卡在捕食者食道,从而损伤捕食者消化器官甚至卡死捕食者;猎物规格过小时,捕食者则需要捕获更多的猎物以满足其生长需求,从而消耗更多的能量[4],因此捕食者的捕获收益可直接影响捕食者对猎物的选择[11,12]。已有研究表明,猎物体高(体宽)为捕食者口裂的60%时捕食者可获得最大捕食收益,并以此确定最佳摄食规格[4,13]。尽管如此,捕食者对猎物的选择是复杂多变的[14],捕食者[4,13,15]、猎物[2,6,16]及环境因子[17,18]均影响捕食者对猎物的选择。
河川沙塘鳢(Odontobutis potamophilaGünther)为小型肉食性鱼类,肉质鲜美,售价80—200元/kg,在江浙沪等地已形成稳定的消费市场。在自然环境中,沙塘鳢常在隐蔽处伏击饵料生物,主要以虾和小型底层鱼类为食,兼食水生昆虫幼虫及小型的螺、蚬等底栖动物,偶尔残食同类[19,20];由于口裂大,沙塘鳢捕食的食物长度可达其体长的50%—60%[20]。养殖者普遍认为日本沼虾(Macrobrachium nipponensisde Haan)为沙塘鳢的优选饵料,但该虾的售价较高,作为饵料生物时养殖成本也很高。在配合饲料研制及其驯食技术没有解决之前,人工养殖的低成本的饵料鱼可作为沙塘鳢的饵料,但是,我们不清楚沙塘鳢喜欢摄食哪种饵料鱼及对饵料鱼大小的选择性。因此,通过室内外试验,我们研究了沙塘鳢对生活习性不同的8种饵料生物的种类选择性,对不同规格赤眼鳟的选择性,以及其不同生长阶段对饵料鱼大小的选择性,以期筛选适宜的饵料种类,确定适口的规格。
1 材料与方法
1.1 沙塘鳢对饵料生物种类和规格选择性研究
2019年7月1日至30日,在武汉市农业科学院水产所山坡养殖基地实验室开展沙塘鳢对饵料生物选择性研究。沙塘鳢及所需饵料生物均来自山坡养殖基地,麦鲮(Cirrhinus mrigalaHamilton)、赤眼鳟(Squaliobarbus curriculusRichardson)和鳊(Parabramis pekinensisBasilewsky)为人工养殖饵料鱼,来自饵料鱼养殖池塘,麦穗鱼(Pseudorasbora parvaTemminck &Schlegel)、寡鳞飘鱼(Pseudolaubuca engraulisNichols)、高体鳑鲏(Rhodeus ocellatusKner)、子陵吻鰕虎鱼(Rhinogobius giurinusRutter)和日本沼虾,来自同一水生植物净水塘。将沙塘鳢暂养在室外3个蓄水箱中(600 L),8种饵料生物分别暂养在室外3个具网兜的蓄水箱中(600 L),实验用水均为曝气超过48h自来水,暂养期间持续充气,暂养7d以上。暂养期间沙塘鳢饱食投喂,饵料生物每天定期投喂两次配合饲料,暂养期间溶解氧含量≥5 mg/L,水温24—28℃。选取统一规格的沙塘鳢,试验前饥饿48h。试验在室内玻璃缸中进行,7:00开灯,20:00关灯。试验鱼缸为透明玻璃缸,长120 cm,宽50 cm,高60 cm,缸体底部、左、右和后面4个面均用不透明黑色纸遮挡减少相邻缸中鱼体干扰。在玻璃缸1/3处架设不透明有机玻璃板,将玻璃缸分隔为大小两室,比例为2∶1,水深保持在50 cm。设置有、无仿真水草两个处理组,水草处理组在水族缸内放置2棵仿真金鱼藻(株高45 cm,覆盖面0.04 m2),分别放置在隔板两侧。
种类选择性试验:2019年7月18日随机选取16尾规格统一、饥饿处理的沙塘鳢(71.9±4.1) mm(均值±标准差),每个缸大室放置1尾沙塘鳢,小室放置8种不同生活习性的饵料生物(麦鲮、赤眼鳟、鳊、麦穗鱼、寡鳞飘鱼、高体鳑鲏、子陵吻鰕虎鱼和日本沼虾)各1尾,8种饵料生物全长、体重见表 1,每个处理组重复8次。适应2h后,21:00依次抽出隔板开始试验,定期观察被摄食饵料鱼种类,暗周期用弱光快速照射查看,24h后结束试验并统计剩余饵料生物种类。

表1 八种饵料生物全长、体重Tab.1 Total length (TL) and body weight (BW) of eight species prey
规格选择性试验:2019年7月22日随机选取16尾统一、规格饥饿处理的沙塘鳢(67.8±3.5) mm,口裂宽(7.6±0.7) mm,口裂高(6.5±0.5) mm(均值±标准差)。每个缸大室放置1尾沙塘鳢,小室放置3种规格(小规格、中规格和大规格)赤眼鳟各1尾,各规格饵料鱼全长、体重、体高和体宽见表 2,每个处理组重复8次。适应2h后,22:00依次抽出隔板开始试验,定期观察被摄食饵料鱼规格,暗周期用弱光快速照射查看,24h后结束试验并统计剩余饵料鱼规格。

表2 三种规格赤眼鳟全长、体重、体高和体宽Tab.2 Total length (TL),body weight (BW),body height (BH) and body width (BW) of S.curriculus with three sizes
1.2 沙塘鳢在不同生长阶段对饵料大小选择性研究
2020年7月1日至9月10日,在武汉市农业科学院水产所山南养殖基地开展不同生长阶段沙塘鳢对饵料鱼规格选择性研究。沙塘鳢及所需饵料生物均来自山南养殖基地,沙塘鳢及饵料生物均暂养在室外6个水泥池(12 m2)中,水泥池中放置6—8棵仿真金鱼藻(株高45 cm,覆盖面0.04 m2),暂养期间持续充气,水温和光照均为自然条件。暂养期间饱食投喂沙塘鳢,饵料鱼每日投喂2次商业饲料,驯化1周后开始试验。
2020年7月23日在3个水泥池(12 m2)中分别投放130尾沙塘鳢(59.3±6.8) mm,每个水泥池中放置6—8棵仿真金鱼藻。养殖过程中定期投放多种规格的饵料鱼(麦鲮),饵料鱼投放量超出沙塘鳢摄食需求量,每次投放前抽样测量饵料鱼全长、体重,试验期间饵料鱼每天投喂2次商业饲料。饵料鱼投放后第2天(2020年8月4日、8月22日和9月4日)捕捞并选取摄食的沙塘鳢,测量其全长、体长、体重、体高、体宽、口裂高和口裂宽,解剖后取其胃含物,测量胃含物中饵料鱼长度。保持完整的饵料鱼直接测量其全长或体长,对于部分消化的饵料鱼则测量其未消化骨骼的长度,依据骨骼长度推算其全长[21]。
1.3 数据处理与分析

式中,TLprey为饵料生物全长(mm);TLpredator为捕食者全长(mm);PPR为饵料生物全长与捕食者全长比。

沙塘鳢对不同种类、规格饵料生物的捕食选择性采用Chesson[22]提出的选择指数表示,式中ai为捕食者对第i种种类或规格饵料生物的喜好指数,ni(j)和ri(j)分别为第i(j)种种类或规格的饵料生物投放的数量和被捕食的数量,m是饵料生物种类或规格的分组数。
运用IBM SPSS Statistics 22.0和Excel软件进行数据分析及作图。不同处理组间沙塘鳢对饵料生物的种类、规格选择指数经正平方根转换后进行单因素方差分析(One-way ANOVA),差异显著时用Duncan’s test进行多重比较。运用Pearson相关指数分析沙塘鳢全长与其体重及饵料鱼全长和PPR值的相关性,运用双样本Kolmogorov-Smirnov Test法比较沙塘鳢胃含物中饵料鱼与环境中饵料鱼的大小分布频率[15]。P<0.05为差异显著水平。
2 结果
2.1 沙塘鳢对饵料生物种类和规格的选择性
如图 1所示,有、无仿真水草处理组间,沙塘鳢对同一种类饵料生物的选择指数均无显著性差异(P>0.05)。有仿真水草和无仿真水草条件下,沙塘鳢对鲮的选择指数均显著大于其对剩余7种饵料生物的选择指数(P<0.05),沙塘鳢对剩余7种饵料生物选择指数间无显著性差异(P>0.05)。
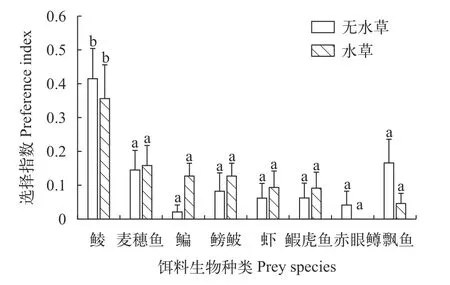
图1 不同水草处理组沙塘鳢对不同种类饵料生物的选择指数Fig.1 Preference index of eight species classes of O.pota mophila in different vegetation treatments
如图 2所示,有、无仿真水草处理组间,沙塘鳢对同一规格饵料生物的选择指数均无显著性差异(P>0.05)。在无仿真水草条件下,沙塘鳢对小规格饵料生物的选择指数显著大于其对大、中规格饵料生物的选择指数(P<0.05),而其对大、中规格饵料生物选择指数间无显著性差异(P>0.05);在仿真水草条件下,沙塘鳢对小规格饵料生物的选择指数显著大于其对大规格饵料生物选择指数(P<0.05),沙塘鳢对中规格饵料生物选择指数与其对大、小规格饵料生物选择指数均无显著性差异(P>0.05)。在无仿真水草条件下,沙塘鳢对小规格饵料生物的选择指数显著大于仿真水草条件下沙塘鳢对大规格饵料生物的选择指数(P<0.05)。
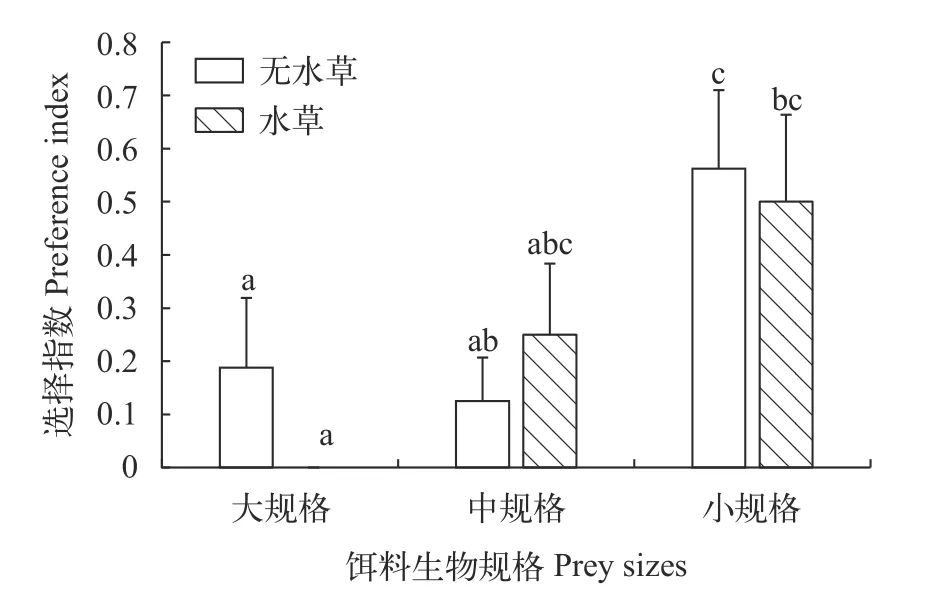
图2 不同水草处理组沙塘鳢对不同规格饵料生物的选择指数Fig.2 Preference index of three size classes of O.potamophila in different vegetation treatments
2.2 沙塘鳢不同生长阶段对饵料鱼大小选择
沙塘鳢全长为43—91.5 mm,投喂麦鲮全长为24—63 mm。沙塘鳢全长与体重相关关系为:W=0.0019 TL3.9855(n=96,R²=0.937,P<0.001)。沙塘鳢全长与麦鲮全长相关关系为:y=0.3448x+1.0834(n=39,R²=0.152,P=0.014)。
在沙塘鳢生长过程中,其胃含物中麦鲮的PPR值为0.23—0.73,PPR均值为0.49±0.11(mean±SD),PPR值在0.31—0.7频率较高,可达94.9%(图 3)。以不同采样时间进行分组,结果表明随着沙塘鳢规格的增加,摄入麦鲮的PPR值趋于降低(图 4)。沙塘鳢全长与PPR值相关关系未达到显著水平(P=0.206;图 5)。
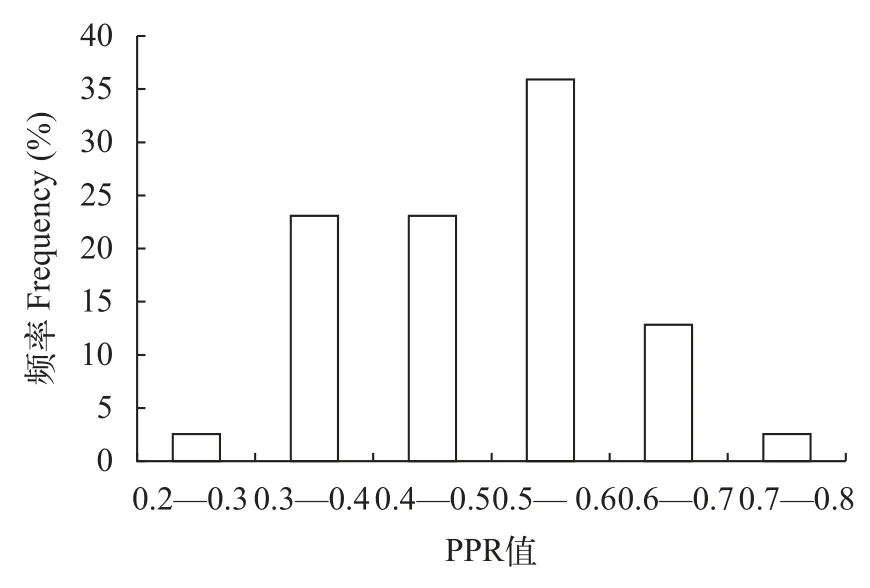
图3 沙塘鳢摄入麦鲮PPR值分布频率Fig.3 Frequency of PPR for C.mrigala fed by O.potamophila

图4 不同时期沙塘鳢摄入麦鲮PPR值相对频率变化Fig.4 Relative frequency of PPR for C.mrigala fed by O.potamophila during different stages

图5 沙塘鳢全长与摄入的麦鲮PPR值相关关系Fig.5 Relationship between PPR for the fed C.mrigala and total length of the predator O.potamophila
沙塘鳢在3个采样时间摄入麦鲮的规格分布与环境中麦鲮规格分布无显著性差异(P>0.05),但摄入麦鲮规格分布小于环境中麦鲮规格分布,摄食较多环境中偏小规格的麦鲮(图 6)。

图6 不同时期环境与沙塘鳢胃含物中不同规格麦鲮频率比较Fig.6 Length frequency histograms for C.mrigala captured in the environment and frequency polygon for C.mrigala fed by O.potamophila during different stages
3 讨论
3.1 沙塘鳢对饵料生物种类和规格选择
多种肉食性鱼类如鳜(Siniperca chuatsiBasilewsky)[21]、大口黑鲈(Micropterus salmoidesLacépède)[7]、白斑狗鱼(Esox luciusLinnaeus)[2]和金鲈(Perca flavescensMitchill)[15]等是受口裂限制的捕食者,吞食饵料鱼规格小于捕食者口裂,大于口裂的饵料鱼不易被捕食。在本研究中,在无仿真水草条件下,8种饵料生物均被沙塘鳢捕获,表明沙塘鳢可捕获口裂范围内多种饵料生物。而在有仿真水草条件下,赤眼鳟未被成功捕获,可能与赤眼鳟习性有关,赤眼鳟较其他种类饵料生物活跃,反捕食能力较强。不论仿真水草存在与否,较侧扁形鳊、鳑鲏及虾而言,沙塘鳢捕获较多中下层活动梭形饵料生物鲮和麦穗鱼,这与一些肉食性鱼类一样捕食较多梭形(圆形)、软鳍条的饵料鱼[2,6,14,21]。其中,沙塘鳢捕获较多人工养殖饵料鱼鲮,不仅与人工养殖鲮的游速慢、反捕食能力弱有关,人工养殖饵料生物长期生活在无捕食压力条件,反捕食意识较弱也起到一定作用[2,23]。复杂生境可以为捕食者和猎物提供隐蔽物[24,25]。不同于斜齿鳊(Rutilus rutilusLinnaeus)、赤睛鱼(Scardinius erythrophthalmusLinnaeus)和河鲈(Perca fluviatilisLinnaeus)等捕食者的捕食活动在复杂生境受到阻碍[26,27],伏击捕食者沙塘鳢同海马(Hippocampus abdominalisLeeson)和斑点雀鳝(Lepisosteus oculatusWinchell)常隐蔽于复杂生境中,捕食成功率增加[25,28]。较无仿真水草条件而言,在有仿真水草条件下,沙塘鳢捕获较多饵料生物,捕获鳊和鳑鲏数量增加,而沙塘鳢捕获到赤眼鳟和飘鱼数量降低,可能与饵料生物生活水层、运动能力等有关,在捕食者、仿真水草的影响下表现出不同的响应[29]。
沙塘鳢24h内对饵料生物规格选择研究结果表明,在无仿真水草条件下沙塘鳢成功捕获饵料鱼集中在9h内,随后未能成功捕获饵料鱼,而在仿真水草条件下沙塘鳢成功捕获饵料鱼的行为持续至试验24h。这与饵料生物的反捕食意识有关,饵料鱼投入初期,未经捕食刺激其反捕食意识较差,无水草条件下随着捕食者暴露及捕食刺激,反捕食意识、反捕食能力增加[2],不易被捕食;而复杂生境为伏击捕食者提供良好的隐蔽物,降低其暴露机会,提高其捕食率[25,28],仿真水草的存在弱化了饵料鱼的反捕食意识增强效应,利于沙塘鳢长时间成功伏击捕食饵料鱼。白斑狗鱼[2]、尖吻鲈(Lates calcariferBloch)[30]和金鲈[15]等肉食性捕食者多捕获反捕食能力弱、体型较小、单独的猎物[31,32]。沙塘鳢出现类似现象,无论仿真水草存在与否,24h内沙塘鳢均捕获较多小规格饵料鱼,不仅因为大规格饵料鱼的体高、体宽接近捕食者口裂大小,不易被捕食者咬住、吞食,小规格饵料鱼体高、体宽小于沙塘鳢口裂大小,更容易被成功捕获;而且与小规格饵料鱼的游速慢、反捕食能力较弱有关[2],较小个体猎物大规格个体具有较高的速度和动力,具有较强的反捕食能力[2,15,30],更易逃脱[33]。
3.2 沙塘鳢在不同生长阶段对饵料大小的选择
沙塘鳢口裂大,可摄食饵料规格较大,同鳜等多种捕食者类似,这些捕食者对饵料鱼的规格选择范围及食谱随捕食者规格的增加而增大[4,15,21,34],摄食饵料鱼的PPR常超过0.5,但随沙塘鳢规格的增加其摄食饵料生物PPR值呈减小趋势。尽管捕食者选择小个体猎物现象不符合最适摄食理论-捕食者以能量收益最大化为原则[4,30],但捕食者选择小个体饵料的现象依然较多[11,15]。相较于鳜摄食饵料鱼的PPR值集中在0.23—0.33[21,35],沙塘鳢养殖过程中其摄食饵料鱼PPR值为0.38—0.6,可能与捕食策略不同有关,较鳜可伏击、追击捕获饵料生物而言,沙塘鳢多伏击捕获饵料鱼,容易捕获较大规格饵料生物。捕食者和猎物的行为能力决定捕获猎物的规格[11,12],大规格饵料鱼的反捕食能力较强,鳜养殖过程中其摄食鲮规格分布显著低于环境中鲮的规格分布[35],沙塘鳢养殖过程中出现类似现象,其摄食鲮的规格分布低于环境中鲮规格分布,沙塘鳢捕食较多环境中偏小规格的饵料鱼。
河川沙塘鳢口裂大,能摄入规格超过其全长的一半的饵料生物。沙塘鳢可捕获口裂范围内的8种饵料鱼,仿真水草生境的存在影响沙塘鳢对饵料生物的捕获,反捕食能力弱的饵料鱼(养殖鲮)及小规格饵料生物更易被捕获。在养殖过程中,随着河川沙塘鳢规格的增加摄食饵料鱼的PPR值趋于降低,摄食饵料鱼规格小于环境中饵料鱼规格,养殖过程中适宜投喂饵料鱼PPR值为0.38—0.6。
