lncRNA-6617调控猪肌内前体脂肪细胞分化的筛选与功能研究
2022-07-07李睿霄姬梦婷蔡春波高鹏飞郭晓红曹果清李步高
孟 珊,杨 阳,李睿霄,姬梦婷,张 娜,路 畅,蔡春波,高鹏飞,郭晓红,曹果清,李步高
(山西农业大学动物科学学院,太谷 030801)
脂肪组织传统上被认为是机体内主要的能量储存器官。随着分子研究层面的不断深入,研究发现脂肪组织也是机体最大的内分泌器官之一,通过分泌大量的脂肪细胞因子影响全身新陈代谢。脂肪组织分布在动物机体内的多个部位,如皮下脂肪和内脏脂肪等,其中存在于肌纤维之间和肌纤维束之间的肌内脂肪,是决定动物肉品质的重要因素之一,影响着肉的口感和风味。研究显示,肌内脂肪细胞来源于肌肉血管基质部分(sromal vascular fraction, SVF)中的肌内前体脂肪细胞,逐步分化形成成熟的脂肪细胞。近年来,随着功能基因组学研究和转录组测序技术的应用,越来越多的调控因子被报道参与脂肪生成的调控。长链非编码RNA(long noncoding RNA, lncRNA)是一类转录本长度大于200 nt的RNA分子,通过影响下游关键因子的表达在脂肪细胞生成过程中发挥着重要的调控作用。但关于lncRNA调控肌内脂肪生成的研究还较少,其调控网络尚不够完善。目前,关于lncRNA调控脂肪细胞生成的研究主要集中于下游的分子机制,lncRNA本身是如何被调控?其调控的分子机制还尚待研究。N6-甲基腺苷(N6-methyladenosine,mA)是一类在真核生物RNA上广泛存在的表观遗传修饰,研究发现,lncRNA本身存在着mA修饰,mA修饰水平的变化影响着lncRNA 功能的发挥,从而参与细胞增殖、纤维化、癌症等的调控。2014年,首次发现了去甲基化酶通过mA修饰的靶mRNA的选择性剪切,影响脂肪生成。但是mA修饰是否可以通过介导lncRNA的作用参与成脂分化的调控还需进一步揭示。
马身猪是山西省地方品种,表现出肉质优良、繁殖力较高、抗逆性强,且背最长肌中肌内脂肪含量极显著高于大白猪。研究发现,在马身猪肌肉组织中,脂肪细胞生成的关键调控因子423、和表达量显著升高,说明马身猪的肌内脂肪细胞生成相较于大白猪显著增加。因此,本研究通过对肌内脂肪含量高的马身猪和肌内脂肪含量较低的大白猪背最长肌组织进行lncRNA测序,从而筛选到一个在马身猪中特异高表达的lncRNA-6617。通过生物信息学软件分析其生物学特性,比较分析lncRNA-6617在马身猪和大白猪肌肉组织间的表达差异。利用体外分离培养的猪肌内前体脂肪细胞为模型,干扰lncRNA-6617,探究其对猪肌内前体脂肪细胞成脂分化的影响。为了寻找lncRNA-6617上游作用机制的调控方式,检测马身猪和大白猪肌肉组织中mA修饰相关酶的表达差异,为后续分子作用机制的深入研究提供理论基础。
1 材料与方法
1.1 试验材料
选取饲养在相同条件下的1、90、180日龄健康马身公猪和大白公猪各3头,分别屠宰采集其心、肝、脾、肺、背部和腹部皮下脂肪、背最长肌、腰大肌、股二头肌等组织,在液氮中速冻,随后于-80 ℃冰箱保存备用。
1.2 主要试剂与仪器
1.2.1 主要试剂 胎牛血清和Opti-MEM购自美国Gibco公司;胰酶、青链霉素、高糖DMEM和胰岛素购自美国Sigma公司;Dex、IBMX、4%多聚甲醛和饱和油红O染液购自北京Solarbio公司;吲哚美辛购自阿拉丁;Trizol Reagen和PARISKit购自美国Life Technologies公司;Prime Script RT reagent Kit with gDNA Eraser和SYBR Premix Ex Taq II等购自北京全式金;Lipofectamine 2000 Reagent购自Invitrogen公司;引物由上海生工合成;siRNA委托上海吉玛公司设计并合成。
1.2.2 主要仪器 细胞成像系统(EVOS FL Auto,Life Technologies,美国),Nanodrop 2000核酸蛋白测定仪(Thermo,美国),普通PCR仪(Bio-RAD,美国),实时荧光定量PCR仪(Bio-RAD,美国)。
1.3 试验方法
1.3.1 RNA提取及cDNA文库的构建和测序 按照Trizol说明书方法提取1、90和180日龄马身猪和大白猪背最长肌组织RNA,经质量检测合格后,进行Illumina Hiseq上机测序。
1.3.2 猪lncRNA-6617的筛选及生物信息学分析 通过edgeR和cuffdiff软件对1日龄马身猪与大白猪背最长肌lncRNA测序结果分析,选定马身猪中特异高表达的lncRNA-6617为目标lncRNA。 利用UCSC-Blast和Ensemble-Blast明确lncRNA-6617的基因组定位;通过CPC、CPAT在线网站预测lncRNA-6617的蛋白编码能力。
1.3.3 lncRNA-6617的鉴定 根据测序得到的lncRNA-6617序列信息,用Primer Premier 6.0软件进行引物设计,以马身猪背最长肌组织cDNA为模板,通过RT-PCR对lncRNA进行扩增,随后用1%琼脂糖凝胶电泳检测扩增产物,对目的片段进行胶回收,连接转化,挑取阳性单菌落进行PCR鉴定,将筛选出的阳性菌送至华大基因测序。
1.3.4 猪lncRNA-6617的核质定位 试验选用未分化的猪肌内前体脂肪细胞,按照试剂盒说明书进行细胞核和细胞质RNA分离,把RNA反转录成cDNA,通过qRT-PCR检测lncRNA-6617在猪肌内前体脂肪细胞的细胞核和细胞质中的表达量。
1.3.5 猪肌内前体脂肪细胞的成脂分化 马身猪肌内前体脂肪细胞由实验室前期分离培养和保存,将传代细胞接种于6孔板,采用“激素鸡尾酒”法诱导成脂。待细胞完全汇合后,更换为诱导培养基(完全培养基+10 μg·mLIns+0.5 mmoL·LIBMX+1 μmoL·LDEX+100 μmoL·LIND),4 d后更换为维持培养基(完全培养基+10 μg·mLIns),每2 d换1次培养基,直至脂滴融合成大脂滴。
1.3.6 细胞转染及油红O染色 将肌内前体脂肪细胞接种于6孔板,密度达到60%左右时进行转染。转染前更换为新鲜培养基,siRNA试剂按照Lipofectamine 2000说明书步骤转染,每组设置3个重复孔。转染48 h后收集细胞,检测干扰效率。同时收集转染后诱导分化7 d的细胞,检测分化中后期成脂关键基因的表达。待细胞脂滴明显形成后,用预冷的PBS清洗两遍,4%多聚甲醛固定30 min,弃去甲醛,60%异丙醇浸洗1 min,弃去异丙醇。待细胞完全干燥,油红O染液(饱和油红O∶蒸馏水=3∶2)染色10 min,PBS洗去多余的油红O染液,显微镜下观察拍照。
1.3.7 总RNA的提取与实时定量PCR 按照Trizol说明书方法提取RNA。按照全式金反转录试剂盒说明书合成第一链cDNA,稀释为20倍。然后以该cDNA作为模板,通过qRT-PCR检测基因表达情况,-为内参基因,引物序列见表1。反应体系为10 μL:SYBR为5 μL,cDNA为4.4 μL,上、下游引物各为0.3 μL。反应程序:95 ℃预变性3 min;95 ℃变性10 s,60 ℃退火20 s,72 ℃延伸30 s, 40个循环。基因相对表达量根据2法计算。

表1 引物信息
1.3.8 数据统计与分析 试验均设置3个生物学重复,所有数据结果使用SPSS Statistics 22.0软件one-way ANOVA和独立样本检验进行显著性分析,采用Duncan′s法进行多重比较,<0.05表示差异显著,<0.01表示差异极显著。
2 结 果
2.1 lncRNA-6617的筛选
利用edgeR和cuffdiff软件分析不同发育阶段马身猪和大白猪背最长肌组织中差异表达的lncRNA,结果显示,1日龄马身猪与90日龄马身猪、1日龄马身猪与180日龄马身猪、90日龄马身猪与180日龄马身猪相比,分别有88、60、18个差异表达的lncRNAs (图1A、1B),1日龄马身猪与1日龄大白猪、90日龄马身猪与90日龄大白猪、180日龄马身猪与180日龄大白猪相比,分别有87、21、16个差异表达的lncRNAs(图1C、1D)。
通过比较分析1日龄马身猪与大白猪背最长肌差异lncRNA的表达量,筛选到在马身猪中特异高表达的lncRNA-6617(图2)。
2.2 lncRNA-6617的生物学鉴定
利用测序得到的lncRNA-6617序列放入UCSC和Ensemble数据库进行比对,发现lncRNA-6617位于猪第18号染色体上,由3基因所在DNA链的反义链转录而来,与3基因内含子3具有278 bp的碱基互补区域(图3A),提示该lncRNA 为反义型lncRNA。
通过CPAT及CPC在线网站对lncRNA-6617进行蛋白编码能力预测分析,选择已报道的非编码基因1为参考,结果表明,lncRNA-6617序列不具有编码蛋白质的能力(图3B)。利用未分化的猪肌内前体脂肪细胞进行核质分离试验,确定lncRNA-6617 的亚细胞定位,结果显示,lncRNA-6617主要分布于细胞核(图3C)。
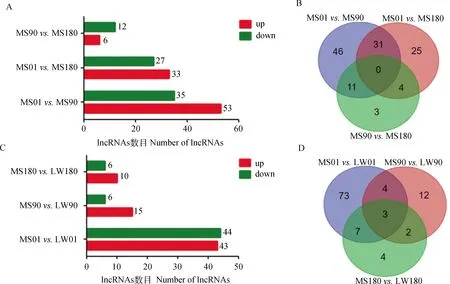
MS. 马身猪;LW. 大白猪。下同MS. Mashen pig; LW. Large White pig. The same as below图1 马身猪和大白猪不同阶段差异表达的lncRNA数量Fig.1 The number of differentially expressed lncRNAs at different stages of MS and LW pigs

图2 1日龄马身猪和大白猪差异lncRNA热图(A)和火山图(B)Fig.2 The difference lncRNAs heat map (A) and volcano map (B) of 1-day-old MS and LW pigs
为验证序列的存在,以转录组测序获得的序列为模板设计引物,通过RT-PCR扩增lncRNA-6617。琼脂糖凝胶电泳结果显示,扩增出长278 bp的目的条带(图3D),条带清晰且单一,Sanger测序结果表明条带序列比对一致(图3E)。

A. 基因组位置;B. 蛋白编码能力预测:a为CPAT结果,b为CPC结果;C. 亚细胞定位;D. PCR扩增;E. 测序结果与已知序列比对A. Genomic location; B. Protein coding ability prediction: a is the CPAT result, b is the CPC result. C. Subcellular localization; D. PCR amplification; E. Alignment of sequencing results with known sequences图3 lncRNA-6617的生物学特性Fig.3 Biological characteristics of lncRNA-6617
2.3 lncRNA-6617的表达模式分析
为明确lncRNA-6617组织表达特性,通过qRT-PCR检测了lncRNA-6617在马身猪心、肝、脾、肺、背部和腹部皮下脂肪、背最长肌、腰大肌、股二头肌等9个组织中的表达情况。结果显示,lncRNA-6617 在各组织中均有表达(图4A),背部皮下脂肪中表达量最高,其次是腹部皮下脂肪等组织。进一步发现,lncRNA-6617的表达量在肌内脂肪含量高的马身猪肌肉组织中极显著高于肌内脂肪含量较低的大白猪(<0.01,图4B)。
对猪肌内前体脂肪细胞进行成脂诱导分化,分别收集诱导0、1、3、5、7 d的细胞,qRT-PCR检测和lncRNA-6617在成脂诱导分化中的表达情况。结果表明,随着成脂分化进程的进行,lncRNA-6617 表达量持续升高,与成脂核心调控因子的趋势一致(图4C、4D),相较于未分化阶段,该lncRNA主要集中在分化中后期表达。
2.4 干扰lncRNA-6617抑制猪肌内前体脂肪细胞成脂分化
将siRNA和对照组silnc-NC分别转染至猪肌内前体脂肪细胞中,48 h后收集细胞检测干扰效率。结果显示,与silnc-NC组相比,silnc-6617组极显著降低了lncRNA-6617的表达(<0.01,图5A)。干扰lncRNA-6617后,对猪肌内前体脂肪细胞进行诱导分化7 d后,分化的成熟脂肪细胞明显减少(图5C),成脂关键基因、和2的表达均极显著下调(<0.01,图5B),表明干扰lncRNA-6617抑制猪肌内前体脂肪细胞分化。
2.5 m6A去甲基化酶FTO调控lncRNA-6617的表达
为了探究lncRNA-6617上游的调控机制,本研究分析了马身猪和大白猪肌肉组织mA修饰水平的差异,结果显示,mA去甲基化酶的表达水平在马身猪背最长肌和股二头肌中极显著高于大白猪(<0.01),在马身猪腰大肌中显著高于大白猪(<0.05), 上游调节因子217的表达水平在马身猪各个肌肉组织中均极显著高于大白猪(<0.01,图6)。
基于SRAMP数据库,利用lncRNA-6617序列预测mA修饰位点,发现在lncRNA-6617上存在mA潜在的修饰位点GGACU/A(图7A)。在猪肌内前体脂肪细胞中干扰,发现干扰极显著降低了lncRNA-6617的表达(<0.01,图7B),说明mA修饰可能介导lncRNA-6617的表达。
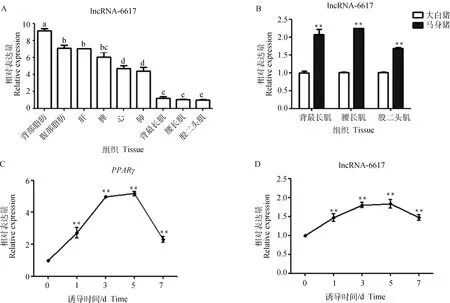
A. 组织表达谱,不同小写字母表示差异显著(P<0.05);B. 马身猪和大白猪肌肉组织间的比较;C、D. 诱导肌内前体脂肪细胞分化过程中PPARγ和lncRNA-6617的时序表达,**. P<0.01,*. P<0.05,下同A. Tissue expression profile, different lowercase letters indicate significant difference(P<0.05); B. Comparison of muscle tissues between Mashen pigs and large White pigs; C, D. Temporal expression during induction of intramuscular preadipocytes differentiation, **. P<0.01, *. P<0.05, the same as below图4 lncRNA-6617表达模式Fig.4 lncRNA-6617 expression patterns
3 讨 论
lncRNA是一类转录本长度大于200 nt的RNA分子,通常不编码蛋白质,而是以RNA的形式在表观遗传修饰、转录调控以及转录后修饰等生物学事件中参与蛋白编码基因调控。随着高通量测序的发展,一系列调控脂肪细胞生成的lncRNA 相继被鉴定,其发挥作用的机制也不断被揭示。lncRNA通过多种作用方式影响关键转录因子和等调控脂肪细胞生成。最近研究也发现,lncIMF4和IMFlnc1参与调控猪肌内脂肪细胞的生成。但是,目前在猪肌内脂肪细胞生成的研究中,更多侧重于测序数据的分析,对于lncRNA的调控成脂分化功能研究尚不深入。
由于lncRNA具有明显的时空表达特异性,使得研究肌内脂肪细胞生成需要进一步筛选鉴定特异表达的lncRNA。此前有研究者分别对绒山羊和肉用山羊肌内脂肪细胞成脂分化前后的细胞进行高通量测序,筛选与肌内沉积相关的功能候选基因。李嫒等利用RNA-Seq技术对大白猪和莱芜猪肌内脂肪中circRNA进行鉴定,以筛选出调控猪脂肪沉积的circRNA,证明高通量测序是筛选功能基因的重要手段。本研究利用本课题组对肌内脂肪含量高的马身猪和肌内脂肪含量较低的大白猪背最长肌组织lncRNA测序结果,筛选到在马身猪中特异高表达的lncRNA-6617。进一步通过生物信息学分析和功能鉴定,发现lncRNA-6617在马身猪肌肉组织中显著高表达,说明lncRNA-6617可能是影响肌内脂肪沉积差异的一个关键因子。同时发现lncRNA-6617在猪肌内前体脂肪细胞成脂分化早期表达量逐渐上升,干扰lncRNA-6617后,显著抑制肌内脂肪细胞的生成,说明lncRNA-6617是调控猪肌内脂肪细胞生成的关键因子。lncRNA-6617功能的发现可以进一步在理论上完善肌内脂肪细胞生成的调控网络,为解析肌内脂肪组织发育机制和遗传选育筛选靶标的确立提供理论依据。
作为遗传信息的传递载体,RNA上的修饰对其功能的发挥起着至关重要的作用。前期已有研究对金华猪、长白猪及长金猪的脂肪组织与mRNA mA修饰水平进行相关性分析,发现mA修饰酶在肉脂型金华猪脂肪组织中表达量最高。同样有研究发现,肌内脂肪含量高的金华猪背最长肌中mA修饰水平显著低于含量较低的长白猪。本研究中,的表达在肌内脂肪含量高的马身猪肌肉组织中显著高于肌内脂肪含量较低的大白猪,且与lncRNA-6617的组织表达模式一致,提示马身猪和大白猪肌内脂肪含量的差异可能与mA修饰相关。研究发现,在猪肌内前体脂肪细胞中,过表达甲基转移酶3显著抑制了脂肪细胞的生成。mA修饰作为RNA表观修饰方式之一,同样影响着lncRNA功能的发挥。于是本研究在肌内前体脂肪细胞中干扰,发现lncRNA-6617 的表达显著下调,说明mA修饰的去甲基化酶可能介导lncRNA-6617功能的发挥,但其具体作用机制还需进一步研究。
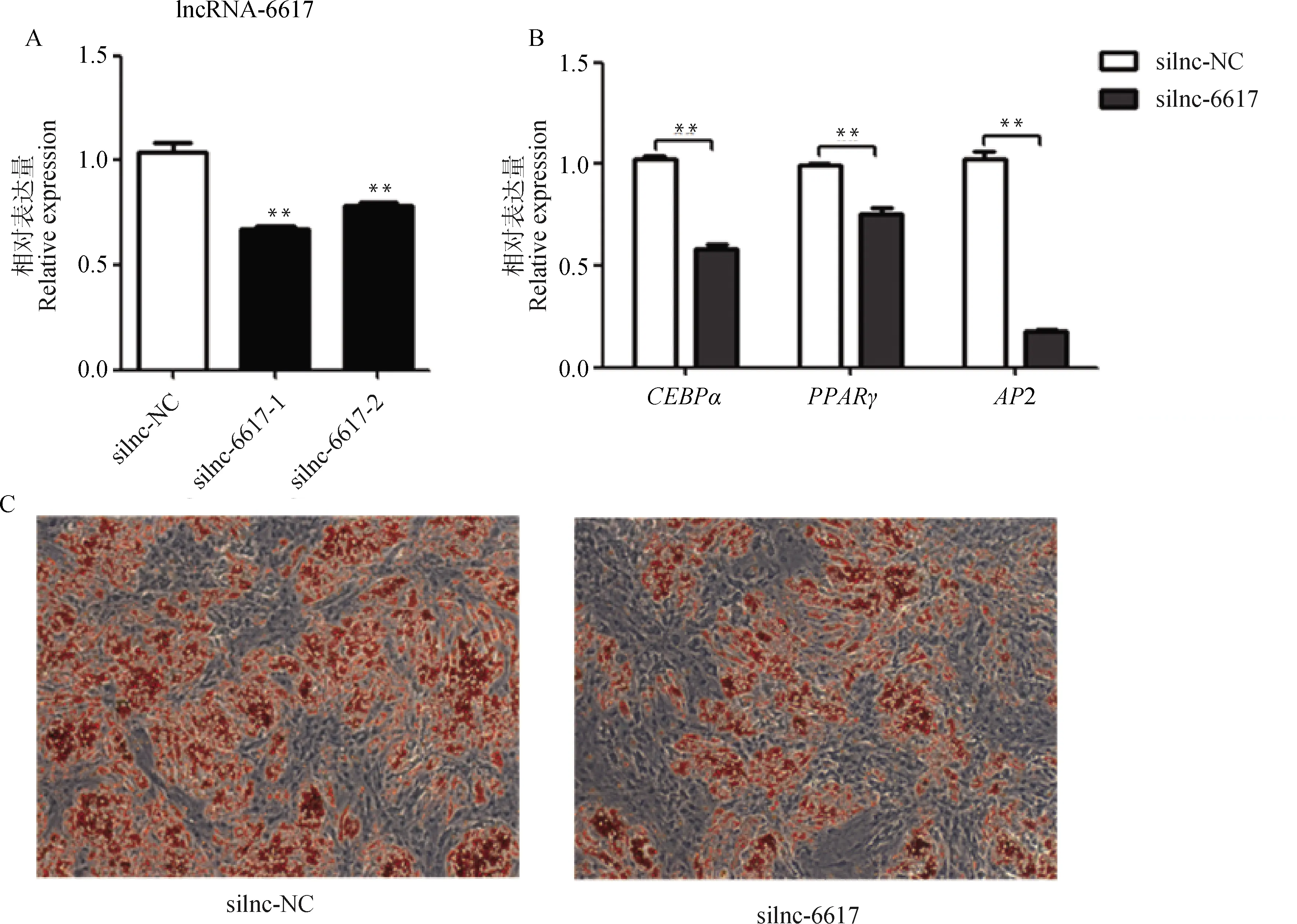
A. 干扰效率;B. 干扰lncRNA-6617后分化7 d成脂相关基因的表达变化;C. 油红O染色(100×)A. Interference efficiency; B. The expression changes of adipogenesis-related genes after interfering with lncRNA-6617 on day 7 of diferentiation; C. Oil Red O staining (100×)图5 干扰lncRNA-6617抑制猪肌内前体脂肪细胞成脂分化Fig.5 Interfering with lncRNA-6617 inhibits adipogenic differentiation of pig intramuscular preadipocytes

图6 马身猪和大白猪肌肉组织中m6A修饰相关酶的表达Fig.6 The expression of m6A modification related enzymes in muscle tissues of MS and LW pigs
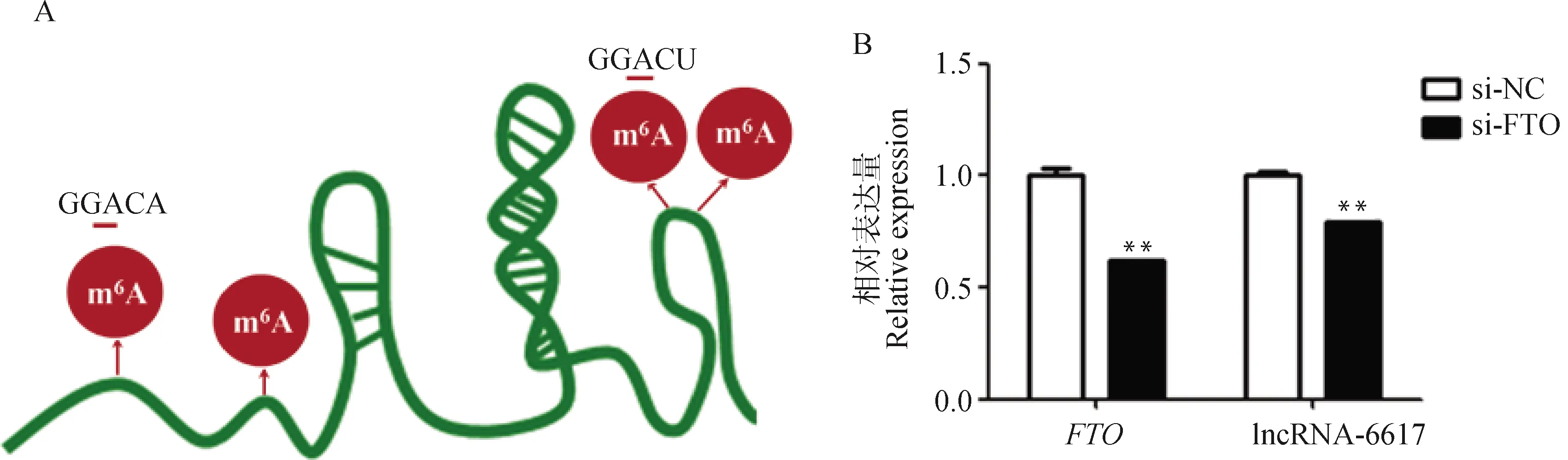
A. lncRNA-6617上m6A修饰位点预测;B. FTO干扰效率以及干扰FTO后lncRNA-6617的表达变化A. Prediction of m6A modification site on lncRNA-6617; B. The expression changes of FTO and lncRNA-6617 after interfering with FTO图7 FTO可能调控lncRNA-6617的表达Fig.7 FTO may regulate the expression of lncRNA-6617
4 结 论
本研究鉴定了在马身猪肌肉组织中特异高表达的lncRNA-6617;干扰lncRNA-6617后,显著抑制猪肌内前体脂肪细胞分化;lncRNA-6617的表达也可能受到上游mA去甲基化酶的调控;这些结果说明,mA修饰可能介导lncRNA-6617参与肌内脂肪细胞生成的调控,为后续深入研究猪肌内前体脂肪细胞生成的分子机制提供了理论基础。
