中华羊茅内生真菌发酵液对垂穗披碱草种子萌发的影响
2022-06-29许文博王美宁
许文博,王美宁,田 沛
(兰州大学草地农业生态系统国家重点实验室 / 兰州大学农业农村部草牧业创新重点实验室 /兰州大学草地农业科技学院, 甘肃 兰州 730020)
近年来,禾草内生真菌共生体的研究受到人们的广泛关注。禾草内生真菌是指在植物体内渡过部分或者全部生命周期的一大类真菌,内生真菌与禾草共生不会使禾草产生外部症状[1-2],目前对内生真菌的研究主要集中于Epichloë属内生真菌[3]。内生真菌与禾草互惠互利,一方面内生真菌生存于宿主植物的细胞间隙,获取宿主体内的营养物质来维持生长;而另一方面,内生真菌可以合成生物碱等来提高宿主植物对生物胁迫和非生物胁迫的抗性,增强宿主的适应性[3-4]。已证实内生真菌可以提高宿主的生长以及抗逆性,研究发现多年生黑麦草(Lolium perenne)内生真菌(Epichloë festucaevar. lolii)对小麦(Triticum aestivum)根腐离蠕孢(Bipolaris sorokiniana)、新月弯孢菌(Curvularia lunata)、细交链孢菌(Alternaria alternata)、德氏霉(Drechslerasp.)等病原菌有明显的抑制作用[5]。被内生真菌感染的禾草(endophyte-infected, E+)具有较强的抗旱、耐寒和耐盐碱性[6-7],尤其在羊茅属的植物中,内生真菌能显著提高禾草地上与地下部分的生长,增加宿主植物的生物量[8]。与未被内生真菌感染的植物相比,禾草-内生真菌共生体对外界环境中重金属的耐性更强,被内生真菌侵染可以提高紫羊茅(Festuca rubra)、细羊茅(F. rubrassp.commutata)对铝(Al)、锌(Zn)、铜(Cu)的耐受性,增加其生物量,降低镉(Cd)对宿主植物的毒害作用[9]。
中华羊茅(F. sinensis)是分布在我国西北地区的多年生羊茅属植物,绝大多数中华羊茅均被内生真菌感染,部分带菌率高达100%。带菌的中华羊茅具有抗旱、抗寒、再生性好等优良特性,是天然草地上垂穗披碱草(Elymus nutans)和垂穗鹅观草(Roegneria nutans)的优良伴生种[10-11]。垂穗披碱草是分布于青藏高原地区的多年生披碱草属植物,部分植株也被内生真菌感染,其植株带菌率低至10%,平均带菌率为35%[12]。中华羊茅与垂穗披碱草在天然草地上广泛分布,混播时中华羊茅可提高垂穗披碱草的产量与抗性,增强草地群落稳定性,提高生产力[13-14]。然而,中华羊茅内生真菌在宿主伴生中的作用尚不得而知。
基于此,本研究从不同地理种群的中华羊茅种子中分离得到3 株内生真菌,探究其不同浓度的发酵液对不带菌(E−)的垂穗披碱草种子萌发的影响,以期为中华羊茅内生真菌的开发和利用提供一定的科学依据。
1 材料与方法
1.1 试验材料
1.1.1 培养基
纯化培养基为马铃薯葡萄糖琼脂(potato dextrose agar, PDA)培养基,参照Li 等[15]的制作方法。本试验采用马铃薯葡萄糖(potato dextrose broth, PDB)发酵培养基。
1.1.2 试验菌株
团队成员前期从来自甘肃(35°12′ E,102°31′N)、四川(32°48′ E,102°33′ N)、青海(36°20′ E,102°06′ N)的中华羊茅种子中分离出3 株中华羊茅内生真菌(Epichloë sinensis),分别为菌株1、菌株111 和菌株41D,内生真菌经分离纯化后转接至90 mm 的PDA 培养基上,然后放置于25 ℃真菌培养箱中,保存于兰州大学草地农业科技学院草地保护研究所。
1.1.3 供试种子
团队成员于2016 年8 月在青海省门源县(101°24′ E,37°25′ N)采集垂穗披碱草种子,带回实验室清选后并镜检确认不带内生真菌后,放置于4 ℃冰箱中,保存于兰州大学草地农业科技学院草地保护研究所。
1.2 试验方法
1.2.1 中华羊茅内生真菌的活化
将保存的3 株中华羊茅内生真菌从培养箱中取出,挑取边缘部分菌丝接种在PDA 培养基上,于25 ℃、黑暗条件下生长1 个月备用。
1.2.2 中华羊茅内生真菌发酵液的制备
将已活化的内生真菌菌株用6 mm 打孔器打孔,放入250 mL 锥形瓶中,每个锥形瓶中含有150 mL PDB 培养基,每个锥形瓶内均放入3 枚菌饼,用封口膜封口后放入摇床,于25 ℃、150 r·min−1条件下培养。21 d 后将菌液过滤,此部分为100%浓度,并将部分内生真菌发酵液用超纯水稀释为20%和60%两个浓度。
1.2.3 不同浓度发酵液对种子发芽的影响
参照《牧草种子检验规程》中发芽试验的相关规定采用纸上发芽法[16],将冰箱中保存的垂穗披碱草种子在5%的次氯酸钠溶液消毒8 min,随后用无菌水冲洗3 次,然后用75%的酒精消毒5 min,接着用无菌水冲洗3 次,再用无菌滤纸轻轻将种子表面的水分吸干,最后将种子均匀地摆放于铺有灭菌滤纸的培养皿(直径为9 cm)中,每皿含有50 粒种子,加入3.5 mL 不同浓度中华羊茅内生真菌发酵液,对照为不同浓度的PDB 液体培养基。本试验总共设置3 个菌株(1、111、41D)及1 个无菌株对照(CK),每个菌株及对照均设置3 个浓度(20%、60%和100%),共计12 个处理,每个处理4 个重复。将种子置于25 ℃恒温培养箱中培养,按照种子胚根长度超过2 mm 为发芽标准,每天统计种子发芽数,并添加适量相对应的发酵液以保持滤纸的湿润状态,发芽期为12 d。第5 天测定种子发芽势,21 d 后每个重复随机选取10 株幼苗,测定并记录其胚根长、胚芽长、鲜重及干重,计算发芽率(germination rate, GR)、发芽指数(germination index, GI)、活力指数(vigor index,VI)、胚根胚芽比(root/shoot, R/S)、幼苗含水量、萌发抗旱指数(germination drought resistance index,UDRI)、活力抗旱指数(vigor drought resistance index,VDRI)。
指标计算方法[17-24]:
发芽率 = (发芽种子数/供试种子数) × 100%;
发芽势 = (发芽数达高峰的前1/3 天数内正常发芽种子数/供试种子数) × 100%;∑
发芽指数= (Gt/Dt);
式中:Gt为在时间t日的发芽数;Dt为相应的发芽日数。
活力指数=GI×Sx;
式中:GI为发芽指数;Sx为平均单株鲜重。
幼苗含水量 = 幼苗单株鲜重 − 幼苗单株干重;
胚根胚芽比 = 胚根长度/胚芽长度;
萌发抗旱指数 = 处理组萌发指数/对照组萌发指数;
活力抗旱指数 = 处理组活力指数/对照组活力指数。
1.3 数据处理
采用Excel 2010 录入并绘图,SPSS 18.0 统计分析软件进行显著性分析。采用双因素方差(two-way ANOVA)分析比较分析不同处理间的差异,根据方差分析的结果,对发芽率、发芽势、发芽指数、活力指数、胚芽长、胚根长、胚根胚芽比和幼苗含水量进行双因素方差分析,通过LSD 法(least significant difference test)检验其差异显著性,用Duncan 法进行多重差异比较;而对种子萌发抗旱指数和活力抗旱指数采用单因素(发酵液或内生真菌)进行分析,结果用均值 ± 标准差表示。
2 结果与分析
双因素方差分析结果(表1)表明,内生真菌菌株及发酵液浓度显著影响了所有检测的指标(P<0.05);而二者相互作用显著影响了发芽率、发芽势、发芽指数、活力指数、胚芽长、胚根长、胚根胚芽比和幼苗含水量(P< 0.05)。

表1 发酵液浓度和菌株对垂穗披碱草种子萌发指标的双因素方差分析结果Table 1 Two-way ANOVA on seed germination indicators of Elymus nutans affected by endophyte strain and fermentation broth concentration
2.1 不同内生真菌菌株发酵液对垂穗披碱草种子萌发特性的影响
在不同发酵液处理下,垂穗披碱草种子的发芽率有显著差异(P< 0.05) (图1)。发酵液浓度为20%时,菌株111 发酵液处理下种子发芽率显著低于菌株1 (P< 0.05) ,降低了19%;浓度为60%时,菌株111 发酵液处理下种子发芽率最高且显著高于对照(P< 0.05),提高了14%;浓度为100%时,3 株内生真菌发酵液处理下种子发芽率无显著差异(P>0.05),但均显著高于对照组发芽率(P< 0.05),菌株1、111和41D 分别比对照提高了28%、30%和32%。对照处理下垂穗披碱草种子发芽率随着PDB 发酵液浓度升高而显著下降(P< 0.05);菌株1 和菌株41D 发酵液浓度为100%处理时种子发芽率显著低于其他两个处理(P< 0.05),菌株111 发酵液浓度为60%处理时种子发芽率显著高于其他两个浓度处理(P< 0.05)。

图1 不同浓度中华羊茅内生真菌菌株发酵液对垂穗披碱草种子发芽率的影响Figure 1 Effects of different fermentation broths of Epichloë sinensis strains on germination rate of Elymus nutans seeds
垂穗披碱草种子发芽势也受到内生真菌发酵液及浓度的显著影响(P< 0.05) (图2)。发酵液浓度为20%时,3 株菌株发酵液以及对照处理发酵液间种子发芽势无显著差异(P> 0.05);浓度为60%和100%时,3 株内生真菌发酵液处理下垂穗披碱草种子发芽势均显著高于对照(P< 0.05),且3 株内生真菌发酵液处理之间种子发芽势无显著差异(P> 0.05)。对照处理下垂穗披碱草种子发芽率随着PDB 发酵液浓度升高而显著下降(P< 0.05);菌株111 发酵液浓度为60%时种子发芽势显著高于其他两个浓度处理(P< 0.05),分别比20%浓度和100%浓度增加了41%和52%;菌株41D 发酵液处理下种子发芽势在20%和60%浓度处理时均显著高于100%浓度处理(P< 0.05);菌株1 发酵液处理下种子发芽势在60%浓度处理时最高,显著高于100%浓度处理(P<0.05),而与20%浓度处理无显著差异(P> 0.05)。

图2 不同浓度中华羊茅内生真菌菌株发酵液对垂穗披碱草种子发芽势的影响Figure 2 Effects of different fermentation broths of Epichloë sinensis strains on germination force of Elymus nutans seeds
垂穗披碱草种子发芽指数也受到内生真菌发酵液及浓度的显著影响(P< 0.05) (图3)。发酵液浓度为20%时,3 株菌株发酵液以及对照处理发酵液间种子发芽指数无显著差异(P> 0.05);浓度为60%和100%时,3 株内生真菌发酵液处理下垂穗披碱草种子发芽指数均显著高于对照PDB 发酵液处理(P< 0.05),且3 株内生真菌发酵液处理之间种子发芽指数无显著差异(P> 0.05)。对照处理下种子发芽指数随着PDB 发酵液浓度的升高而显著下降(P<0.05)。菌株1 和41D 内生真菌发酵液处理下种子发芽指数在20%和60%浓度处理时显著高于100%浓度处理(P< 0.05),而菌株111 发酵液浓度为60%时种子发芽指数显著高于其他两个浓度处理(P<0.05),分别比20%和100%浓度提高了17%和27%。

图3 不同浓度中华羊茅内生真菌菌株发酵液对垂穗披碱草种子发芽指数的影响Figure 3 Effects of different fermentation broths of Epichloë sinensis strains on germination index of Elymus nutans seeds
在不同发酵液处理下,垂穗披碱草种子活力指数也有显著差异(P< 0.05) (图4)。发酵液浓度为20%时,菌株41D 处理的种子活力指数显著高于其他两个菌株和对照(P< 0.05),分别比菌株1 和111以及对照提高了41%、49%和11%,且菌株1 和111发酵液处理下的种子活力指数显著低于对照和菌株41D 处理(P< 0.05)。发酵液浓度为60%时,菌株41D 处理下种子活力指数显著高于同浓度其他处理(P< 0.05),分别比菌株1 和111 以及对照提高了41%、49%和43%。发酵液浓度为100%时,3 株内生真菌发酵液处理下种子活力指数均显著高于对照处理(P< 0.05),菌株1、111 和41D 分别比对照提高了59%、49%和62%。对照处理下垂穗披碱草种子活力指数随PDB 发酵液浓度升高而显著下降(P<0.05),菌株1 和111 内生真菌发酵液处理下种子活力指数在不同浓度间差异不显著(P> 0.05),菌株41D 的发酵液浓度为20%、60%处理下种子活力指数显著高于100%处理(P< 0.05),均提高了36%。

图4 不同浓度中华羊茅内生真菌菌株发酵液对垂穗披碱草种子活力指数的影响Figure 4 Effects of different fermentation broths of Epichloë sinensis strains on vigor index of Elymus nutans seeds
2.2 不同内生真菌菌株发酵液对垂穗披碱草种子幼苗根和芽生长的影响
不同发酵液处理下,垂穗披碱草幼苗的胚根长、胚芽长以及胚根胚芽比有显著差异(P< 0.05)(表2)。浓度为20%时,菌株111 发酵液处理下幼苗胚根长、胚芽长以及胚根胚芽比均最低,胚根长显著小于其他3 个处理(P< 0.05),而胚芽长显著小于菌株1 处理(P< 0.05),胚根胚芽比显著小于对照和菌株1 处理(P< 0.05)。浓度为60%时,菌株41D 发酵液处理下幼苗胚根长均显著高于其他3 个处理(P< 0.05),胚芽长与菌株1 和菌株111 处理无显著差异(P> 0.05),胚根胚芽比与菌株111 处理无显著差异但显著高于菌株1 处理和对照(P< 0.05);对照处理下幼苗胚根长、胚芽长以及胚根胚芽比均最低,胚根长显著小于其他3 个处理(P< 0.05),胚芽长显著小于菌株41D 处理(P< 0.05),而胚根胚芽比显著低于菌株111 和41D 处理(P< 0.05)。浓度为100%时,菌株41D 处理下幼苗胚根长、胚芽长和胚根胚芽比均显著大于对照和其他两个菌株发酵液处理(P< 0.05),且菌株111 处理下幼苗胚根长显著高于菌株1 和对照处理(P< 0.05),菌株111 处理下胚根胚芽比显著高于菌株1 和(P< 0.05)。对照处理和菌株1 发酵液处理下,垂穗披碱草幼苗胚根长以及胚根胚芽比随PDB 发酵液浓度升高而显著下降(P< 0.05),而菌株1 幼苗胚芽长在浓度20%处理下显著高于其他两个菌株处理(P< 0.05);菌株1 发酵液处理下,垂穗披碱草幼苗胚根长在浓度100%处理下显著低于其他两个菌株处理(P< 0.05);菌株111 和41D 发酵液处理下,垂穗披碱草胚根胚芽比在浓度100%处理下显著低于60%处理(P< 0.05)。
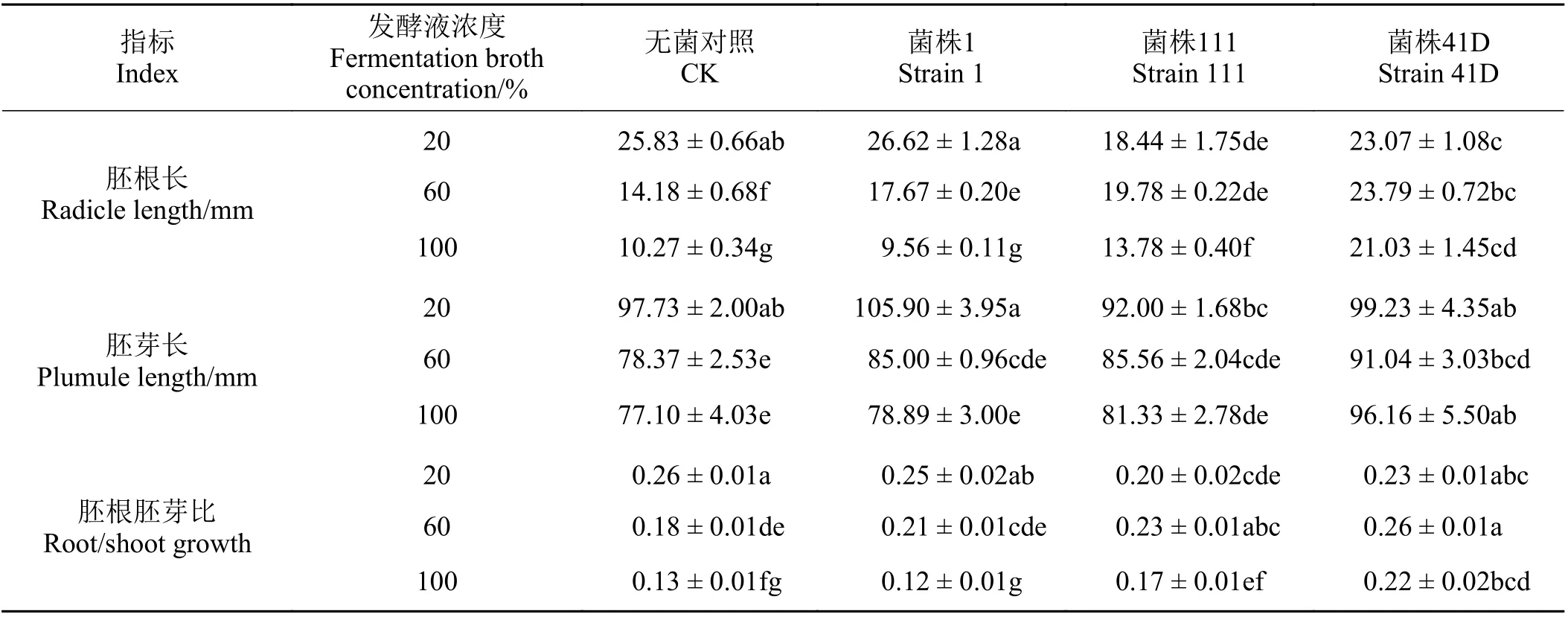
表2 3 株中华羊茅内生真菌菌株发酵液对垂穗披碱草胚根长、胚芽长和胚根胚芽比的影响Table 2 Effects of three Epichloë sinensis endophyte fermentation broths on Elymus nutans plumule length, radicle length, and root to shoot ratio
不同处理对垂穗披碱草幼苗含水量也有显著影响(P< 0.05) (图5)。浓度为20%时,菌株1 发酵液处理的垂穗披碱草幼苗含水量显著高于对照和其他两个菌株处理(P< 0.05),分别比菌株111 和41D以及对照提高了15%、18%和17%。浓度为60%时,内生真菌菌株1 和菌株111 发酵液处理下幼苗含水量显著高于对照处理(P< 0.05),且菌株111 发酵液处理下幼苗含水量显著高于菌株41D 处理(P<0.05)。浓度为100%时,内生真菌菌株41D 发酵液处理下幼苗含水量显著高于对照处理和菌株111 处理(P< 0.05),且菌株111 发酵液处理下幼苗含水量显著高于对照处理(P< 0.05)。对照处理和菌株111发酵液处理下,垂穗披碱草幼苗含水量随PDB 发酵液浓度升高而显著下降(P< 0.05);菌株1 和41D 发酵液浓度为20%处理时幼苗含水量显著高于其他两个浓度处理(P< 0.05)。
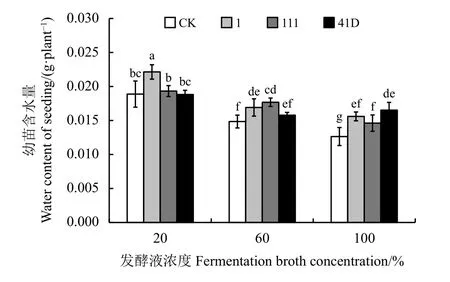
图5 不同浓度中华羊茅内生真菌菌株发酵液对垂穗披碱草幼苗含水量的影响Figure 5 Effects of different fermentation broths of Epichloë sinensis strains on water content of Elymus nutans seedlings
2.3 不同地理种群中华羊茅内生真菌发酵液对垂穗披碱草种子萌发抗旱性的影响
不同发酵液处理下垂穗披碱草种子的萌发抗旱指数与活力抗旱指数不同(表3)。同一浓度不同菌株处理下,20%浓度下菌株41D 处理时种子的萌发抗旱指数和活力抗旱指数均显著大于菌株111 (P<0.05);60%浓度的菌株111 处理时,垂穗披碱草种子的萌发抗旱指数显著大于菌株1 和41D (P<0.05),而菌株41D 处理时种子的活力抗旱指数均显著大于菌株1 和111 (P< 0.05);100%浓度时,菌株41D 和111 处理下种子的萌发抗旱指数显著高于菌株1 (P< 0.05),而菌株41D 处理下种子的活力抗旱指数均显著大于菌株1 和111 (P< 0.05)。同一菌株发酵液处理下,垂穗披碱草种子萌发抗旱指数在60%和100%浓度处理下显著大于20%浓度处理(P<0.05),而活力抗旱指数均随发酵液浓度的增加而显著增加(P< 0.05),同一菌株浓度为100%处理时,垂穗披碱草种子的萌发抗旱指数与活力抗旱指数达到最大。

表3 3 株中华羊茅内生真菌菌株发酵液对垂穗披碱草种子萌发抗旱指数和活力抗旱指数的影响Table 3 Effects of fermentation broths of three Festuca sinensis endophyte strains on Elymus nutans germination drought resistance index and vigor drought resistance index
3 讨论
对高羊茅(Festuca arundinacea)与多年生黑麦草-内生真菌共生体研究中,已开展了有关内生真菌对宿主以外其他植物生长影响的研究,比如Renne 等[25]发现高羊茅提取物对南美草原植物种子萌发的抑制作用与是否感染内生真菌并无显著关系,但Vázquez-de-Aldana 等[26]的研究表明感染内生真菌(E+)的高羊茅对车前草(Plantago lanceolata)、百脉根(Lotus corniculatus)等幼苗的抑制作用显著高于未感染内生真菌(E−)的高羊茅。Sutherland 等[27]研究发现感染内生真菌的多年生黑麦草对三叶草(Trifolium repens)生长的影响显著高于E−多年生黑麦草。而中华羊茅内生真菌发酵液对中华羊茅和醉马草(Achnatherum inebrians)种子萌发有不同程度的促进或者抑制作用,同时发酵液浓度对其也有影响,表现为稀释液促进作用较强,而原液则表现为抑制作用[28]。而Epichloë bromicola内生真菌发酵液促进了拟南芥(Arabidopsis thaliana)幼苗鲜重、干重、叶片数和不定根数等生物量的增加[29]。闫继辰等[30]研究发现,真菌发酵液可以显著促进大豆(Glycine max)种子的萌发与活力指数。本研究以不同地理种群中华羊茅内生真菌发酵液为材料处理E−垂穗披碱草种子,结果表明,高浓度(100%)内生真菌发酵液处理下垂穗披碱草种子的发芽率、发芽势、发芽指数均显著高于对照PDB 发酵液处理,说明内生真菌发酵液对非宿主植物种子萌发有促进作用。在田间条件下,可能通过化感作用影响垂穗披碱草的生长。高浓度内生真菌(菌株111 和41D)发酵液处理下垂穗披碱草种子的培根长和胚芽长均显著高于对照PDB 发酵液处理,而菌株1 发酵液处理与对照处理间无显著差异,说明不同菌株的内生真菌发酵液对非宿主植物种子萌发的促进作用并不一致。
真菌发酵液的抑制或促进作用与其浓度有明显关联。孙锋[31]利用不同浓度哈茨木霉(Trichoderma harzianum)菌株发酵液处理抗虫棉(Gossypium herbaceum)种子,发现稀释10、50、100 倍的木霉发酵液处理的种子发芽势、发芽率、幼苗生长状况均显著优于对照,其中以稀释50 倍处理抗虫棉种子生长最为良好。本研究用分离自不同地理种群中华羊茅的内生真菌发酵液及对照PDB 处理垂穗披碱草种子,除菌株1 发酵液在100%浓度处理下的胚根长小于对照处理外,3 株中华羊茅内生真菌发酵液浓度为60%和100%时垂穗披碱草种子发芽率、发芽势、发芽指数、幼苗含水量、胚根胚芽生长均大于对照,说明中华羊茅内生真菌对垂穗披碱草种子有促进作用。但是随着发酵液浓度增加,其发芽率、发芽势、发芽指数均呈现先升高后下降趋势;除菌株41D 在100%浓度处理下的胚芽长外,胚根长和胚芽长均呈现下降趋势。这与以前真菌发酵液促进种子萌发且与其浓度有关的研究结果一致[28,31]。
真菌发酵液对种子萌发与幼苗生长的影响与其产生的化合物有关。内生真菌不仅在体外产生次生代谢物,在宿主体内也产生代谢物,也能诱导宿主植物产生一些次生代谢物。内生真菌对宿主的有益作用和对家畜健康的影响均与共生体产生的次生代谢物有关[32-34]。赵昕梅等[35]从野生铁皮石斛(Dendrobium officinale)中分离的4 株内生真菌菌株中,镰刀菌属(Fusarium)菌株发酵液对宿主生长具有一定的抑制作用,其中菌株TPSH4 发酵液的促进作用最为显著。王伟等[36]研究发现,核桃(Juglans regia)内生真菌菌株可以产生对小麦(Triticum aestivum)的幼芽和幼根有不同程度抑制作用的代谢产物。陈颐辉等[37]研究发现从水稻(Oryza perennis)具有化感作用的品系中分离得到的11 株内生真菌中多数其发酵液对常规非化感水稻品系种子的萌发均有一定的促进作用,对稗草(Echinochloa crusgalli)种子萌发均有一定程度抑制作用,其中从水稻化感品系中分离得到的燕麦镰刀菌(Fusarium avenaceum)和塔宾曲霉(Aspergillus tubingensis)的发酵产物能促进非化感水稻幼苗生长,并能大幅度提升水稻超氧化物歧化酶和过氧化物酶活性,这两种内生真菌处理稗草其脂肪酶与抗氧化酶活性均显著低于对照,说明这两种内生真菌产生的次生代谢产物中具有能够影响水稻与稗草生长的物质。本研究中中华羊茅内生真菌发酵液对垂穗披碱草种子的促进作用可能也与内生真菌产生的次生代谢物有关。目前已从Epichloë发酵液中分离鉴定出多种化合物,比如倍半萜烯(sesquiterpenes),chokol A、B 和 C[38],3 种酚醛甘油酯[39],4 种C-18 羟基不饱和脂肪酸[40],一种环状芳香甾醇化合物[41]以及一些吲哚衍生物和二乙酰基胺[42]。但是是否是这些化合物对宿主或宿主以外植物种子萌发产生影响尚未开展研究。文先[29]在Epichloë bromicola发酵液中鉴定出生长素如吲哚乙酸(indole acetic acid, IAA)、吲哚乙醇(indole-3-ethanol)和吲哚甲醛(indole-3-carboxaldehyde)等物质,这些化合物可能是影响拟南芥生长的活性物质。而中华羊茅内生真菌是否在发酵条件下产生生物碱还需进一步检查。因此,今后继续对发酵液的次生代谢物成分进行分析,以期寻找出对种子萌发有促进作用的化合物。
本研究从不同地理种群中华羊茅种子中分离出3 株内生真菌菌株,用其发酵液处理E−垂穗披碱草种子,结果表明不同菌株处理下种子生长状况不同,分离自青海省平安县的中华羊茅内生真菌(即菌株41D)处理下在浓度为20%和60%时垂穗披碱草种子发芽率、发芽势和发芽指数显著高于菌株41D 发酵液在浓度为100%时,浓度为100%时种子抗旱指数、幼苗含水量均大于同浓度下其他菌株处理,且幼苗胚根长、胚芽长、胚根胚芽比显著大于其他浓度处理,因此,高浓度下的菌株41D 发酵液处理对种子生长的促进作用强于对应浓度的对照、菌株1、菌株111。而来自不同地理区域的这3 株内生真菌对种子萌发作用的差异,这与前期开展的关于其他生理生化特性以及生长速度的研究结果一致,均明确了这3 株菌株存在差异。从中华羊茅分离出的内生真菌是一个新种,命名为Epichloë sinensis[43],虽然这3 个菌株为同一个种,但不同菌株存在差异,可能是不同的基因型。
本研究结果表明从中华羊茅分离出的内生真菌对与其伴生的垂穗披碱草的种子萌发及幼苗生长有促进作用,但是促进的机制和内生真菌在化感中所起的作用尚未可知。明确发酵液中的化合物成分及功能将有助于解释内生真菌在中华羊茅与垂穗披碱草混生中的化感作用。
