基于非靶向代谢组学的焙焦油莎豆粕精酿啤酒主发酵期代谢物变化研究
2022-06-23张璐璐孙洪蕊刘香英康立宁
张 雷,张璐璐,孙洪蕊,刘香英,康立宁*
(1.吉林省农业科学院 农产品加工研究所,吉林 长春 130033;2.吉林农业科技学院 食品工程学院,吉林 吉林 132101;3.北华大学 林学院,吉林 吉林 132013)
精酿啤酒是特殊的营养原料与创新的发酵工艺共同缔造的新型啤酒类型。精酿啤酒和普通啤酒最大的区别就是精酿啤酒最重视口感,为了口感好,在制作时可以不必在乎节省成本,而选择更好的原料[1]。随着社会的发展,人们更加喜欢独特的、健康的饮品,而精酿啤酒就符合现在人们的要求和需求。相对于普通啤酒的单一,精酿啤酒具有独特性和多样性[2]。伴随消费者购买力及消费观念的提升,中高端啤酒产品的需求将持续扩大范围,精酿市场将迎来多元化的消费升级,精酿啤酒消费开始由感官性需求向追求高营养过渡[3-4]。
研发出更加营养、健康、口感独特的精酿啤酒一直是有关人士追求的终极目标。油莎豆(Cyperus esculentus)又称虎坚果,为百合科植物油莎豆的干燥根茎,是东北地区的重要药材,其性平味甘,具有养阴润燥,生津止渴之功效[5]。有科学研究表明,油莎豆具有降血压、降血脂的作用,且可预防中风,其醇提取物还能有效的延缓衰老、抗菌、防癌[6-7]。由于油莎豆含糖量较高,且随着人们对个性化啤酒研究逐渐成为热点,这使其作为开发特色精酿啤酒原料增加了更多的可能性。本课题组前期研究表明,油莎豆粕精酿啤酒风味物质丰富,口感宜人,货架期稳定性好[8],但该产品在主酵过程中物质变化还不明确。代谢组学研究的是生命体对外界刺激、病理生理变化以及本身基因突变而引起的分子质量<1 500 Da的代谢产物种类、数量及其变化规律,主要通过高通量检测和数据处理,进行信息整合及生物标记物鉴定的科学[9]。已被广泛应用于食品研究中,钟成等[10]采用代谢组学分析技术分析工业啤酒发酵过程中风味物质生成规律。刘予煊等[11]对枸杞发酵产品的研究多关注于枸杞中挥发性成分及其香气特征。王娜[12]基于组学技术分析了中国黄酒陈酿香气组分及酒龄的识别。综上所述代谢组学技术已经在酒的酿造中凸显出其重要作用。本研究利用代谢组学技术分析了油莎豆粕精酿啤酒主发酵期代谢物的变化,旨在为深入解析本产品特色代谢物的形成提供基础组学数据。
1 材料与方法
1.1 材料与试剂
油莎豆粕:世和农业科技有限公司;大麦芽:山东金酿生物科技有限公司;萨兹酒花、US-04酵母:帝伯仕精酿有限公司;乙腈(色谱纯):德国Merck集团;乙酸铵(≥98%):美国Sigma公司。
1.2 仪器与设备
XS-10多功能麦芽粉碎机:上海兆申科技有限公司;300 L啤酒全套生产线:哈尔滨顺成创新科技发展有限公司;AL104电子天平:梅特勒-托利多仪器有限公司;LB-90手持糖度计:河北慧采科技有限公司;DW-86L388A超低温保存箱:海尔股份有限公司;DHG-9053A鼓风干烘箱:上海一恒科学仪器有限公司;Triple TOF 6600质谱仪:美国SCIEX公司;Agilent 1290 Infinity LC超高效液相色谱仪(ultra-high performance liquid chromatography,UPLC):美国安捷伦公司;Eppendorf 5430R低温高速离心机:德国艾本德公司;Waters186004801色谱柱:美国Waters公司。
1.3 方法
1.3.1 原料处理方法
将油莎豆粕进行微波杀菌,温度85 ℃,时间为15 s。干燥箱中170 ℃条件下干燥20 min。以料液比为1∶2.5(g∶mL)加40 ℃温水,浸提3 h后过滤,煮沸杀菌,静置冷却10 h,下层分液,即为发酵液A,用于补料发酵。
1.3.2 啤酒发酵液制备工艺流程与操作要点
糖化→过滤→煮沸→入罐→发酵→补料→二次发酵→灌装
糖化:以100 L发酵体积为例,将粉碎好的20 kg大麦芽投入温度为35 ℃的糖化锅中,加热到45 ℃保持20 min,再加热到50 ℃保持50 min,再加热到72 ℃保持10 min,再加热到78 ℃,进行过滤。
过滤:用筛板过滤后的麦糟用70 ℃的60 L水洗糟两次,制得麦汁。
煮沸:麦汁煮沸温度为100 ℃,开始煮沸时投入35 g酒花,煮沸时间为1 h,煮沸结束时投入30 g酒花,煮沸结束后测糖度为11%。
入罐:麦汁在回旋槽中进行回旋沉淀,静置20 min。入罐温度控制在20 ℃,控制氧气压力0.5 MPa,充氧1 min。
发酵:酵母菌种添加量为0.1%,起始发酵温度为18 ℃,每隔24 h排一次沉淀物,同时测定发酵液糖度,待糖度至4.5%左右,投酒花45 g;随后,每24 h降2 ℃,并同时排酵母与沉淀物,降至4 ℃时停止降温,即为发酵液B。
补料:向发酵液B中加入油莎豆粕液A约为15%。
二次发酵:温度控制在14~16 ℃,7 d后封罐,控制压力在1.5 MPa,每天降2 ℃,排沉淀物,最后温度降至2 ℃后储7 d,即为成品。
灌装:出酒前1 d利用二氧化碳钢瓶将罐内压力补压至2.0 MPa,碳化24 h,备压灌装。
1.3.3 样品制备
分别在主发酵第3天(F-3d),主发酵第6天(F-6d)取样,在-80 ℃超低温冰箱中保存。样本在4 ℃环境下缓慢解冻后,取适量样本加入预冷甲醇/乙腈/水溶液(2∶2∶1,V/V),涡旋混合,低温超声30 min,-20 ℃静置10 min,14 000 g、4 ℃离心20 min,取上清真空干燥,质谱分析时加入100 μL乙腈-水溶液(乙腈∶水=1∶1,V/V)复溶,涡旋,14 000 g、4 ℃离心15 min,取上清液进样分析。
1.3.4 超高效液相色谱分析
样品采用超高效液相色谱(UPLC)系统HILIC色谱柱进行分离;柱温25 ℃;流速0.5 mL/min;进样量2 μL;流动相组成A:水+25 mmol/L乙酸铵+25 mmol/L氨水,B:乙腈;梯度洗脱程序如下:0~0.5 min,95%B;0.5~7 min,B从95%线性变化至65%;7~8 min,B从65%线性变化至40%;8~9 min,B维持在40%;9~9.1 min,B从40%线性变化至95%;9.1~12 min,B维持在95%;整个分析过程中样品置于4 ℃自动进样器中。
1.3.5 飞行时间质谱分析
采用AB 6600三重飞行时间质谱(time of flight mass spectromete,TOF-MS)仪进行样本一级、二级谱图的采集。HILIC色谱分离后的电喷雾离子(electrospray ionization,ESI)源条件如下:
气体1:60 Psi;气体2:60 Psi;气帘气:30 Psi;离子源温度:600 ℃;电压:±5 500 V(正负两种模式);母离子质谱扫描范围:60~1 000 amu;子离子扫描范围:25~1 000 Da;母离子积累时间:0.20 s;子离子积累时间:0.05 s;二级质谱采用信息依赖型扫描,并且采用高灵敏度模式,去簇电压:±60 V(正负两种模式);碰撞能量(35±15)eV。
1.3.6 数据分析流程
Wiff格式的原始数据经ProteoWizard转换成.mzXML格式,然后采用XCMS软件进行峰对齐、保留时间校正和提取峰面积。对XCMS提取得到的数据首先进行代谢物结构鉴定、数据预处理,然后进行实验数据质量评价,最后再进行数据分析。数据分析内容包括单变量火山图统计分析(univariate volcanic map analysis,UVMA)、多维统计分析[包括主成分分析(principal component analysis,PCA)与偏最小二乘法判别分析(partial least squares discrimination analysis,PLS-DA)]、差异代谢物聚类分析与京都基因与基因组百科通路分析(kyoto encyclopedia of genes and genomes,KEGG),具体分析模式参数参考文献[13-14]。
2 结果与分析
2.1 数据质量控质分析
为了证明数据的可靠性与准确性,本研究通过谱图进行重叠比较,结果见图1A。由图1A可知,各色谱峰的响应强度和保留时间基本重叠,说明在整个实验过程中仪器误差引起的变异较小。当对质量控制(quality control,QC)样本进行Pearson相关性分析,相关系数>0.9时说明相关性较好。相关性图谱(图1B)结果表明,QC样本间的相关系数R均在0.9以上,说明实验重复性较好,仪器分析结果可靠,分析条件稳定,为进一步数据分析及生物信息学分析奠定了基础。
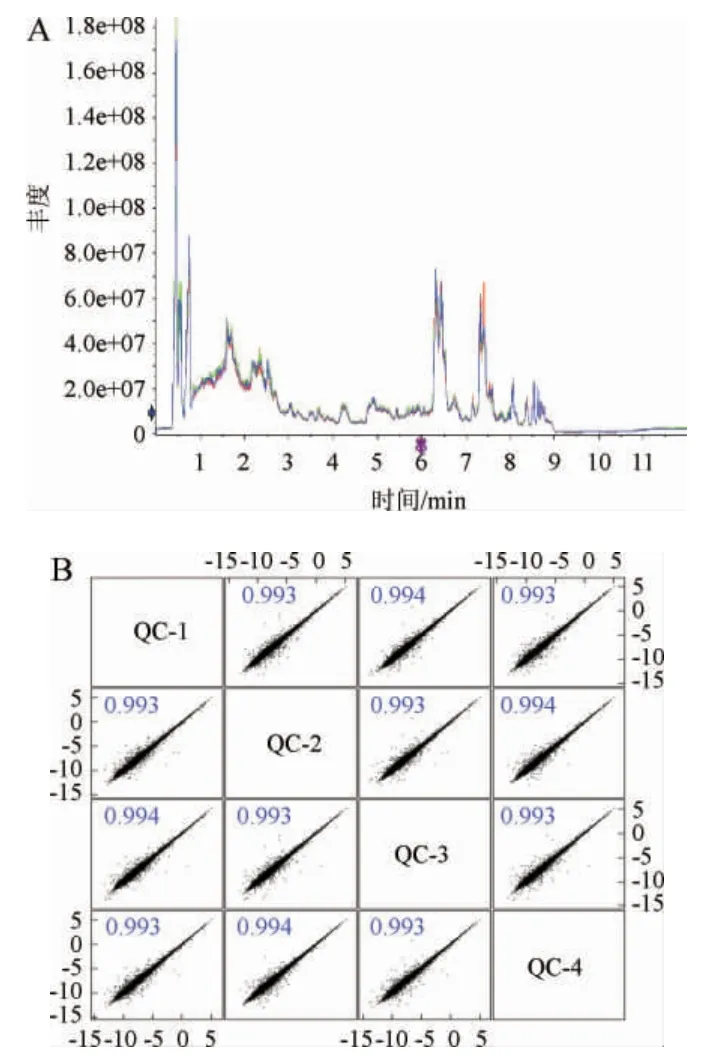
图1 质量控制样本总离子流色谱图(A)及样本相关性(B)Fig. 1 Total ion chromatogram of quality control sample (A) and correlation of samples (B)
2.2 油莎豆粕啤酒主酵期发酵代谢物单变量统计分析
在进行两组样本间的差异分析时,单变量统计分析方法是最常用的统计分析方法之一,常用的单变量统计分析包括变异倍数分析、T检验/非参检验单变量分析,对检测到的所有代谢物(含未被鉴定的代谢物)进行差异分析。同时结合单变量统计分析结果,这些显著性差异代谢物可能是潜在的生物标志物。通过研究差异代谢物参与的代谢通路,逆推找出调节酶和基因,有助于揭示其参与的生命活动机制,完成调控通路等方面的研究。由图2可知,在油莎豆粕精酿啤酒主发酵期共有416种代谢物被检测到,具有显著性差异物质为52种,其中上调差异代谢物数目为30种,下调代谢物数目为22种,这表明啤酒在主酵期代谢反应活跃,主酵期在啤酒成味过程中发挥了重要作用。

图2 主发酵期差异代谢物单变量火山图分析Fig. 2 Univariate volcanic map analysis of metabolites in the main fermentation stage
2.3 油莎豆粕啤酒主酵期发酵代谢物多维统计分析
2.3.1 主酵期发酵代谢物主成分分析
主成分分析(principal component analysis,PCA)是一种非监督的数据分析方法,它将原本鉴定到的所有代谢物重新线性组合,形成一组新的综合变量,同时根据所分析的问题从中选取几个综合变量,使它们尽可能多地反映原有变量的信息,从而达到降维的目的[15]。同时,对代谢物进行主成分分析,还能从总体上反映样本组间和组内的变异度。因此在数据分析中,一般先采用PCA方法,观察组间样本的总体分布趋势和组间样本的差异度。主发酵期发酵液样本主成分分析模型得分图见图3。由图3可知,油莎豆精酿啤酒主发酵期第3天与第6天的12个检测样本都在各组95%置信区间内,说明实验的重现性好,能够保证试验结果的可靠性,在实验中获得的代谢谱差异能反映样本间自身的生物学差异。
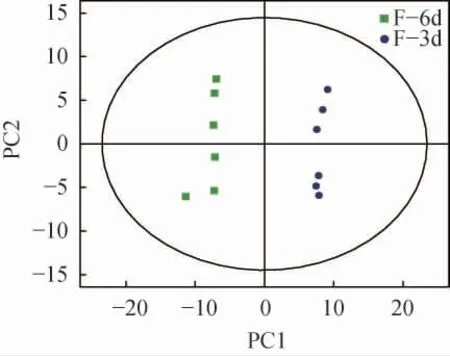
图3 主发酵期发酵液样本主成分分析模型得分图Fig. 3 Principal component analysis model score diagram of samples in the main fermentation stage
2.3.2 主酵期发酵代谢物偏最小二乘法判别分析
经7次循环交互验证得到模型评价参数。一般预测率Q2>0.5,表明模型稳定可靠;0.3≤Q2≤0.5,表明模型稳定性较好;Q2<0.3,表明模型可靠性较低。主发酵期发酵液样本偏最小二乘法判别模型得分图见图4。如图4A所示,本试验主发酵期样本PLS-DA得分结果与PCA结果相近,模型对X变量解释率(R2X)=0.43,模型对变量Y的解释率(R2Y)=0.99,Q2=0.97,表明模型稳定可靠。为避免有监督模型在建模过程中发生过拟合,采用置换检验Permutation test对模型进行检验,以保证模型的有效性。图4B显示了示例对比组PLS-DA模型的置换检验图,随着置换保留度逐渐降低,随机模型的拟合度R2和Q2均逐渐下降,说明原模型不存在过拟合现象,模型稳健性良好,适合探索油莎豆粕精酿啤酒主发酵期的代谢物差异。
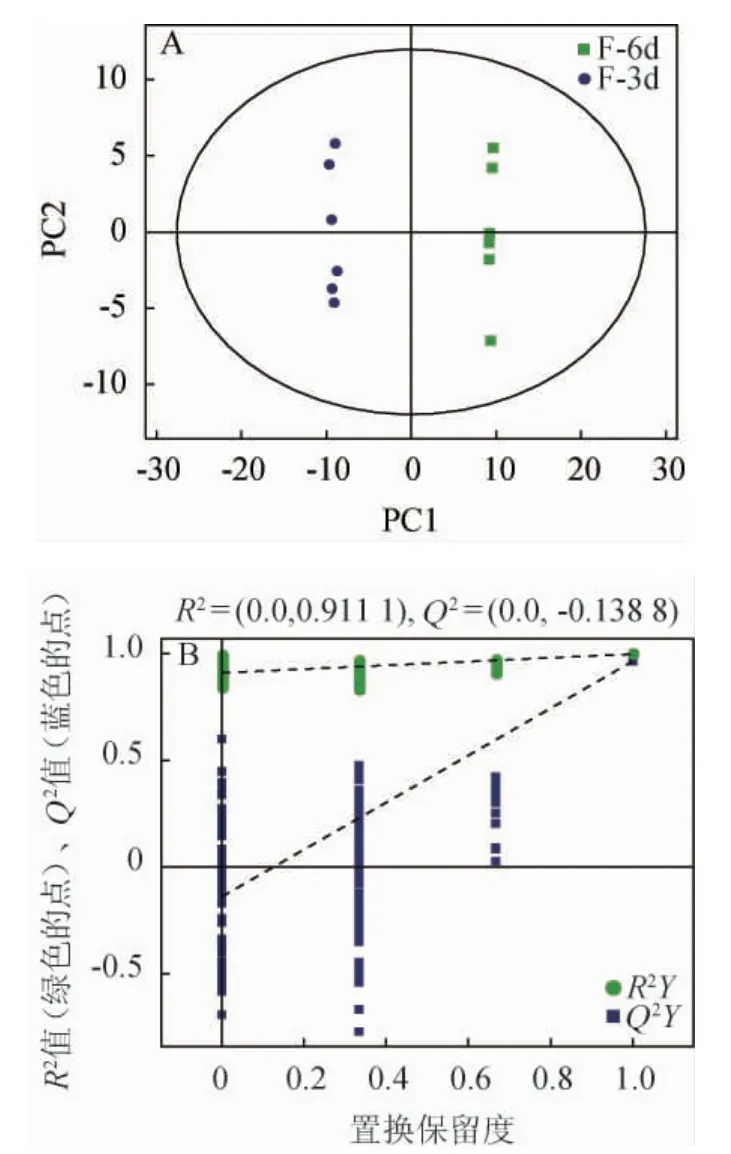
图4 主发酵期发酵液样本偏最小二乘法判别模型得分(A)与置换检验图(B)Fig. 4 Partial least square discriminant model score (A) and replacement chart (B) of fermentation broth samples in the main fermentation stage
2.4 油莎豆粕啤酒主酵期发酵代谢物差异代谢物鉴定
根据PLS-DA模型分析结果,将t检验的P<0.05的代谢物作为筛选标准,主酵期发酵代谢物差异代谢物含量及变化进行测定,结果见表1。
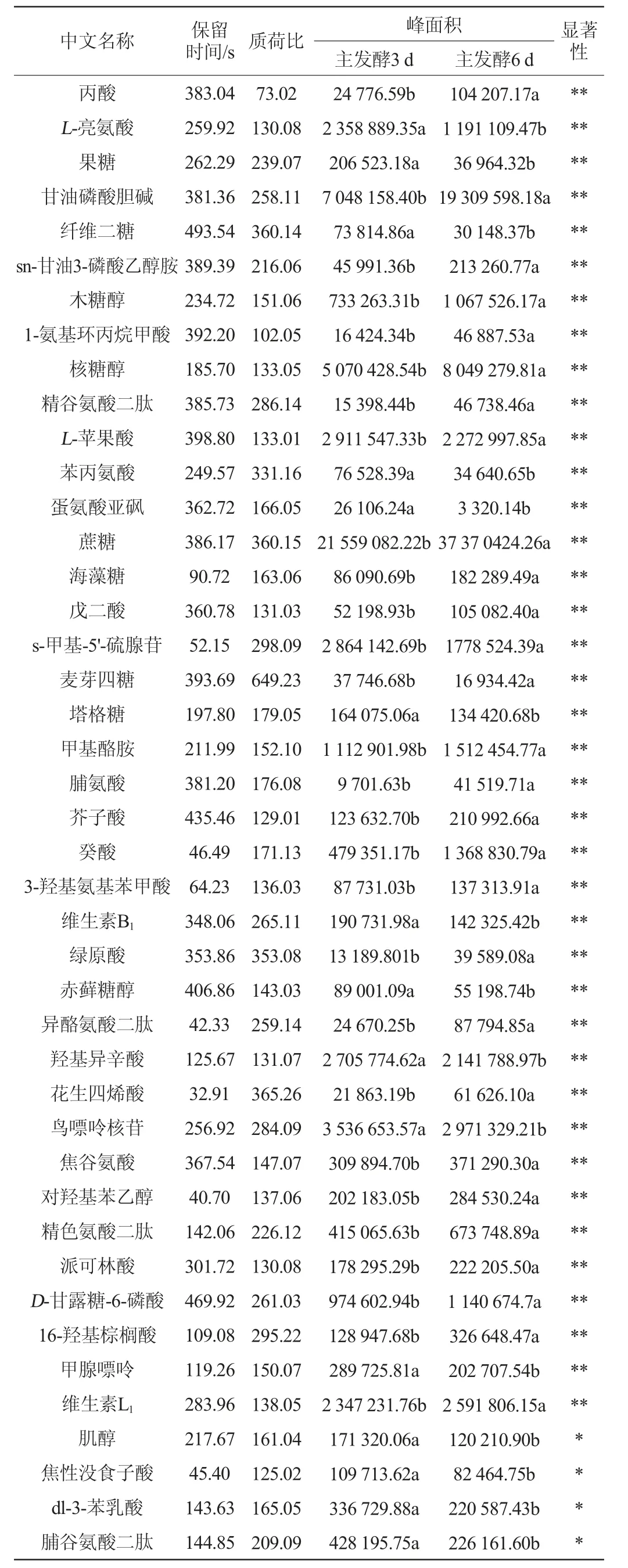
表1 油莎豆粕啤酒主酵期差异代谢物信息Table 1 Information of differential metabolites of Cyperus esculentus meal beer during the main fermentation stage
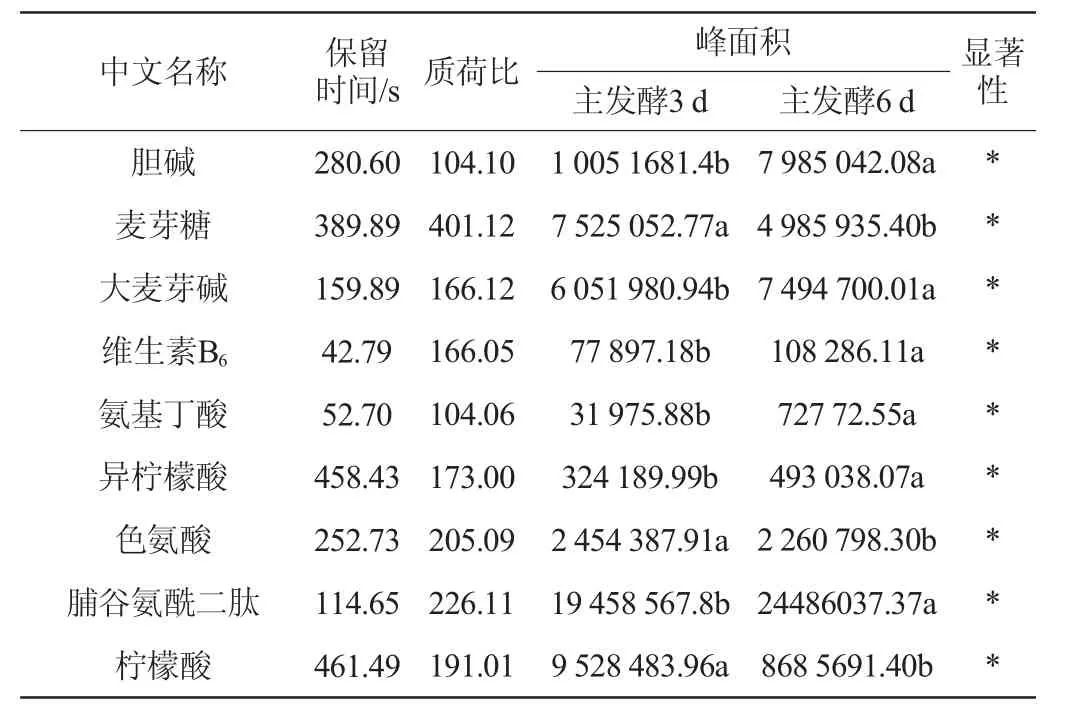
续表
由表1可知,在油莎豆粕啤酒主酵期中共检测到14类52种差异发酵代谢物,这些显著差异合成代谢物主要为有机酸类及其衍生物、氨基酸类及其衍生物、小分子肽类、糖类、脂肪酸类、醇类、维生素类等。有机酸类物质是影响啤酒风味与口感的重要组成部分[16],有机酸的含量与种类对啤酒的口感产生重要的影响[17-18]。绿原酸是植物在有氧呼吸过程中产生的苯丙素类化合物,是抗菌、抗病毒的主要有效药理成分之一,易溶于乙醇,其作为一种重要的生物活性物质,具有利胆、抗菌、降压、增加白血球及兴奋中枢神经系统、抗肿瘤、清除自由基等作用酸类物质[19-20]。本试验结果显示,在主发酵期3~6 d绿原酸含量增加近4倍,这对啤酒营养价值具有重要意义。花生四烯酸是人体的必需脂肪酸,其相关主要代谢产物在肿瘤细胞增殖和凋亡、肿瘤转移、肿瘤血管新生和炎症反应等方面具有重要作用[21]。在啤酒的主发酵期,花生四烯酸含量显著增加,这对啤酒的风味与营养物质的形成具有重要贡献。氨基酸及其衍生物是酵母发酵中重要组成部分,游离氨基酸除了具有生物学功能还具有鲜、甜、涩等特殊味道[22-23]。从啤酒主发酵期筛选出的氨基酸代谢物有脯氨酸、色氨酸、谷氨酸、亮氨酸、苯丙氨酸以及相应的二肽物质,这些物质都为油莎豆啤酒产生重要的风味提供代谢底物与呈味基础。此外,啤酒酯香气的合成与糖醇类物质有着密切的关系[24],主酵期产生了多种醇类与糖类物质,这些物质很可能对啤酒的酯香气合成具有重要功能。
2.5 油莎豆粕啤酒主酵期发酵代谢物差异代谢物聚类分析
代谢物分布可在视觉上分为上调和下调[25]。为了更全面直观地显示样本之间的关系以及代谢物在不同样本中的表达模式差异,将所有样本及差异代谢物的表达量进行距离矩阵计算,并进行层次聚类分析(hierarchical clustering analysis,HCA),建立样本层次聚类树,结果见图5A,依据52个差异代谢物的相对含量作出热图,结果见图5B。
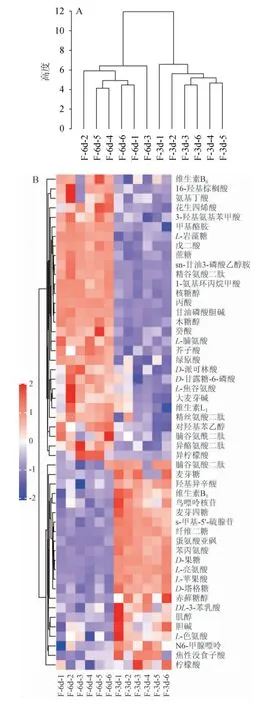
图5 样本层次聚类树(A)及差异代谢物层次聚类热图(B)Fig. 5 Sample hierarchical clustering tree (A) and hierarchical cluster heat map of metabolites (B)
由图5A可知,不同研究组中平行样本都聚在同一簇内,表明聚类结果可信度较高,聚类热图分析的结果可靠。由图5B可知,聚在同一簇内的代谢物具有相似的功能或者共同参与同一代谢过程或者细胞通路,图中每行代表一个差异代谢物(即纵坐标为显著性差异表达的代谢物),每列代表一组样品(即横坐标为样品信息)。红色代表显著性上调,蓝色代表显著性下调,颜色深浅表示上下调的程度,表达模式接近的代谢物聚在左侧同一簇下。根据上方样本聚类的树状图,将5B可以分成两个区域,主发酵期第6天的6个平行样本为第一区域,主发酵期第3天为第二区域。第一区中有22种代谢物,主要是糖类和氨基酸等物质,聚在同一簇中,含量显著低于第二区;有30种代谢物,主要是有机酸和醇类等物质,聚在同一簇中,含量显著高于第二区(P<0.05)。通过层次聚类分析可以看出,主酵期不同发酵阶段对啤酒中各类代谢物的含量具有显著影响,通过比较差异代谢物含量的异同,对鉴别经发酵后油莎豆精酿啤酒中的代谢产物具有一定的参考意义。
2.6 油莎豆粕啤酒主酵期发酵代谢物差异代谢物KEGG功能分析
通过与KEGG数据库比对,可将基因按照参与的pathway通路或行使的功能进行分类,可以获知代谢物参与的代谢通路信息,从而评价其对生物新陈代谢过程的影响[28]。主酵期发酵差异代谢物KEGG功能分析结果见表2。
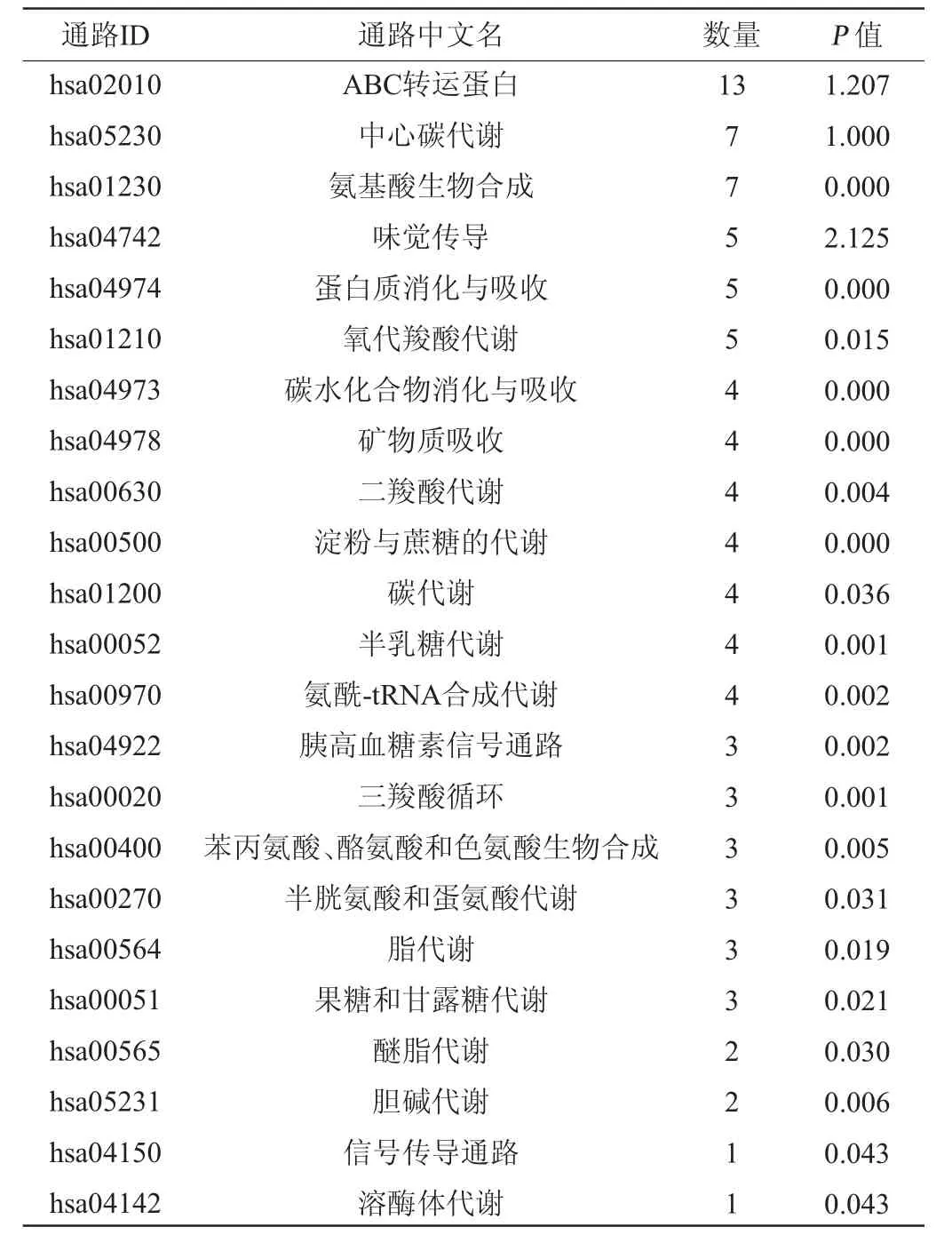
表2 主酵期发酵差异代谢物KEGG功能分析Table 2 Functional analysis of different metabolites KEGG during the main fermentation stage
由表2可知,啤酒主发酵期代谢物中共检索到差异代谢物参与的23条代谢通路,其中代谢物富集到前3的代谢通路为腺苷三磷酸结合转运蛋白(ATP-binding cassette transporter,ABC transporter)、中心碳代谢与氨基酸合成代谢。ABC转运蛋白是一个含有ATP结合结构域、高度保守的一大基因家族编码的一类跨膜蛋白,有机体中约有近千余种不同的转运蛋白,它们可转运的底物包括无机离子、单糖、聚糖、磷脂、氨基酸、肽、蛋白质、抗生素和异源物质等多种物质[29-30],所以在油莎豆粕精酿啤酒主酵期酵母代谢活跃,ABC转运蛋白为啤酒营养成分与风味物质的合成发挥了重要作用。同时氨基酸代谢与中心碳代谢在主酵期也表现了非常活跃的特征,为啤酒品质的形成具有重要的贡献。此外脂代谢、碳代谢、氨酰-tRNA合成代谢、三羧酸循环代谢通路等都在主酵期同样表现出了积极活跃的代谢过程,这为探究外源添加物改善啤酒质量提供了重要生物学信息。
3 结论
油莎豆粕精酿啤酒主发酵期发酵液差异代谢物主成分、偏小二乘判别分析、层次聚类分析反应出试验结果可信度高,且能够聚为同类;在啤酒主发酵期共鉴定到52种显著差异表达的代谢物,其中上调差异代谢物数目为30种,下调代谢物数目为22种。KEGG结果表明,ABC转运蛋白、中心碳代谢与氨基酸合成代谢、脂代谢、三羧酸循环代谢、碳代谢、氨酰-tRNA合成代谢等相关代谢途径在主酵期发挥了重要作用与生物学功能。
