7种青冈属植物种子休眠类型鉴定及休眠打破
2022-06-12史喜兵焦雪辉申潇潇
史喜兵 焦雪辉 申潇潇


摘要:为研究青冈属(Cyclobalanopsis)植物种子休眠特性,缩短播种育苗周期,以7种青冈属植物种子为材料,通过超微结构观察、吸水性测定、白菜种子萌发受内源抑制物影响试验,对休眠类型进行鉴定,并利用机械处理、激素处理等方法进行休眠打破。结果表明,7种青冈属植物种子的种壳呈现非常明显的3层结构,由外而内依次为角质层、外表皮细胞和内表皮细胞。角质层细胞石质化,形成蜡质层,非常坚硬;外表皮细胞呈栅栏状排列,增加了种皮的厚度和硬度。去壳种子和划口种子,其吸水率均高于种壳完整种子,说明种壳对种子的吸水性有一定的阻碍作用。经种壳和子叶浸提液培养的白菜种子萌发率降低,对其根和叶的生长有明显的抑制作用,说明种壳和子叶中含有阻碍种子萌发的物质。去种壳及种壳划口处理可以打破种子休眠,使萌发时间明显提前,萌发率和发芽势显著提高。GA3处理对种子休眠打破影响不大,对部分树种种子的发芽率和发芽势有显著提高作用。表明7种青冈属植物种子的种壳影响胚的生长发育和种子的吸水性,种壳的物理阻碍作用是青冈种子休眠的主要因素。去种壳和种壳划口处理是打破种子休眠的有效方法。
关键词:青冈属;休眠类型;休眠打破;机械处理;激素处理;种壳;子叶
中图分类号: S722.1+4 文献标志码: A
文章编号:1002-1302(2022)10-0151-08
种子休眠的现象普遍存在于高等植物中,是植物本身适应环境和延续生存的一种特性。胚未分化或发育未成熟、代谢抑制物等是导致种子休眠的内源性原因,胚外周组织的物理、机械或化学性抑制等是外源性原因[1]。Nikolaeva按照影响種子萌发的因素形成了一套种子休眠分类系统[2]。Baskin 等对这一分类系统进行了改进并提议作为国际通用的种子休眠分类系统,该系统主要包括物理休眠(physical dormancy,简称PY)、形态休眠(morphological dormancy,简称MD)、生理休眠(physiological dormancy,简称PD)、复合休眠(PY+PD)和形态生理休眠(morphophysiology dormancy,简称MPD)5种[3]。无论是农作物,还是林木、果树、花卉,均存在种子休眠现象。
众多学者对种子休眠因素和种子萌发特性进行了大量研究。相关研究表明,硬度较高、密度较大的种皮(包括果皮),可能由于上披油脂或蜡质,导致种子休眠[4]。郭聪聪等认为,白皮松种子成熟时,种胚已发育完整,胚外部覆盖组织、内外种皮含有的抑制物是影响种子萌发的主要原因,胚乳对萌发无抑制作用[1]。耿文娟等发现,野生欧洲李种子各个部分的浸提液不影响白菜种子萌发,但对胚根生长有明显的抑制作用,其各个部分的浸提液抑制白菜种子萌发的强弱表现为种胚>种皮>种壳[5]。有些植物种子还存在上胚轴休眠现象。仇云云等认为,紫斑牡丹种皮致密、种皮和胚乳含有阻碍萌发物质是胚轴休眠的原因,胚内部原因造成了上胚轴休眠,需要低温解除休眠[6]。也通过机械处理、低温层积、热水浸种、化学药剂浸种等方法,对打破种子休眠进行了研究。
青冈属植物种子大多属于顽拗性种子,不经历成熟脱水,脱落时含水量相对较高,不耐干燥,一般情况下干燥至15%~20%含水量时大多数或者全部死亡。种皮和种壳坚硬致密是导致顽拗性种子休眠的重要原因。李金华研究过赤皮青冈(Cyclobalanopsis gilva)种子萌发特性,采用不同激素、不同果皮和种皮处理方式对种子进行处理,发现果皮和内种皮对种子萌发存在物理性抑制,去除种壳后种子可打破休眠,提高萌发率,激素处理也可以提高萌发率[7]。彭颖姝等研究过不同沙藏处理对青冈栎种子萌发的影响,发现去种皮后种子发芽率明显高于完整种子。种皮透气性很差、透水性不受影响;低温贮藏发芽率高于常温贮藏[8]。李庆梅等研究了栎属7种植物种子的发芽抑制物,认为不仅种皮影响萌发,其种子内源抑制物也可能是种子休眠的原因[9]。目前对青冈属种子休眠的研究多集中在休眠打破方法上,少有从形态结构及内源物质等方面对休眠因素进行系统研究。因此,本研究通过对7 种青冈属植物种子超微结构、种壳吸水性及内源抑制物的研究,希望寻找到阻碍种子萌发的原因,并在此基础上探索休眠打破的方法,为生产中提供种子萌发率、降低繁育成本提供理论支持。
1 材料与方法
1.1 试验材料
2020年秋季采集青冈属植物种子,包括青冈(Cyclobalanopsis glauca)、小叶青冈(C. myrsinaefolia)、细叶青冈(C. gracilis)、赤皮青冈(C. gilva)、曼青冈(C. oxyodon)、多脉青冈(C. multinervis)、云山青冈(C. sessilifolia),其中青冈栎、小叶青冈种子采集地点为中国林业科学研究院亚热带林业实验中心年珠林场,赤皮青冈采集地点为浙江省庆元县实验林场,曼青冈和多脉青冈采集地点为湖北神农架,云山青冈采集地点为浙江省九龙山自然保护区。7个树种分别标记为FC1、FC2、FC3、FC4、FC5、FC6、FC7,从中挑选大小均匀、饱满、无病虫害的种子,放置于郑州市农林科学研究所实验室,洗净晾干备用。
1.2 休眠类型鉴定方法
1.2.1 超微结构观察
取7个树种新鲜种子的种壳,组织块面积不超过3 mm2,用磷酸盐缓冲盐溶液(PBS)轻轻漂洗干净,标记好扫描面,放入电镜固定液中2 h,最后保存于4°冰箱;固定好的样品经 0.1 mol/L PBS(pH值7.4)漂洗3次,每次15 min;0.1 mol/L PBS(pH值7.4)配制1%锇酸室温避光固定1~2 h。0.1 mol/L PBS(pH值7.4)漂洗3次,每次 15 min;用梯度浓度为30%、50%、70%、80%、90%、95%、100%、100%乙醇逐级处理,每次 15 min,然后用醋酸异戊酯处理15 min,再使用临界干燥仪干燥后进行离子溅射仪喷镀,置于Hitachi SU8100扫描电子显微镜下观察。
1.2.2 种壳吸水性
分别取完整种子、划口种子(从种子胚端开始,使用解剖刀向下对称划开种壳,至种子1/2~2/3处,露出种胚但不损伤种胚)及去壳种子各5粒,称量后放入烧杯,加入40 mL蒸馏水,置于25 ℃恒温环境,每隔一定时间取出种子,先用吸水纸吸掉表面水分,再用电子天平称质量,重复3次,种子质量不再增加为止。前8 h,每浸泡2 h取1次,每次取5粒,之后每隔12~48 h可取1次,每次取5粒。使用如下公式计算种子吸水速率:
种子吸水速率=(吸水后种子质量-吸水前种子质量)/吸水前种子质量×100%。
1.2.3 内源抑制物对白菜种子萌发影响
1.2.3.1 甲醇浸提液的提取
取7个树种新鲜的种子,剥开并分离种壳和子叶,分别称取 4 g,放入预冷的研钵内研碎,加入20 mL 80%甲醇溶液,摇晃均匀,置于4 ℃恒温冰箱浸提24 h,为充分浸提,每4 h晃动1次,24 h后过滤,按此方法连续浸提3次,将3次滤液倒入旋转蒸发仪,减压蒸干后蒸馏水冲洗,最后定容到100 mL作为培养液。
1.2.3.2 白菜种子萌发测定
挑选出部分饱满的白菜种子,先用0.5%高锰酸钾溶液消毒,蒸馏水冲洗干净后自然风干备用。培养皿中放入湿润的脱脂棉,脱脂棉上盖1层湿润的滤纸,加入25 mL培养液,对白菜种子进行萌发试验,以蒸馏水为对照。每个培养皿中放入白菜种子30粒,每个处理3个重复,25 ℃恒温光照培养箱培养。24 h后统计每个培养皿中发芽的白菜种子数量,以发芽长度超过种子直径为标准,计算平均发芽率,48、72 h后测量根长,记录生长情况。
1.3 休眠解除
挑选健康饱满的青冈种子,去除杂质和有虫种子,清洗干净晾干,过种子筛进行分级。用3%高锰酸钾溶液处理20 min,清水冲洗3次,将部分种子进行划口和去壳处理。按照不同方法处理后,自种子发芽开始,每天记录发芽数,种子连续15 d不再发芽时结束试验(对照种子培养3个月)。计算发芽率和发芽势。
1.3.1 种壳处理方式的影响
取FC1~FC7完整种子(T0)、划口种子(T2)、去壳种子(T6)各15粒(表1),每组重复3次,常温水浸泡48 h,放入光照培养箱,设置温度25 ℃、8 h黑暗、16 h光照。
1.3.2 激素的影响
取FC1~FC7划口种子各15粒,分别放入常温水浸泡48 h(T2)及GA3 400 mg/L浸泡48 h(T5)(表1)。每组重复3次,置于25 ℃光照培养箱培养,光照16 h、黑暗8 h。
1.3.3 光照时间的影响
取FC5、FC6划口种子15粒,常溫水浸泡48 h,置于25 ℃光照培养箱培养。光照时间设2个梯度:光照16 h、黑暗8 h(T1),全黑暗(T4)(表1),重复3次。
1.3.4 培养温度的影响
取FC5、FC6划口种子15粒,常温水浸泡48 h,置于光照培养箱培养,光照 16 h、黑暗8 h。光照培养箱设置2个温度梯度:25 ℃(T2),20 ℃(T3)(表1)。重复3次。
2 结果与分析
2.1 休眠类型鉴定
2.1.1 种壳扫描电镜观察
从图1可以看出,不同树种种壳厚度及结构不同,其中FC4种壳最厚,其次为FC6、FC7。从横断面图来看,种壳结构分为3层,最外面为角质层,然后是外表皮细胞和内表皮细胞。角质层细胞石质化,形成蜡质层,非常致密,对吸水有严重阻碍作用。外表皮细胞呈栅栏状排列,主要由单层长柱状的大石细胞组成,其细胞壁厚度增加,呈木质化。外表皮细胞增加了种皮的厚度和硬度,对水分进入种子有很大的阻碍作用。内表皮细胞厚度因树种而异,其中FC4的内表皮细胞层很厚,占据了整个种壳的大部分厚度,但排列较为疏松,其他树种内表皮细胞相对较薄。内外表层细胞之间有1层被挤压的薄壁组织,较薄。内表面图显示有很多细长管状或带状的表皮毛,内表皮细胞排列不规则,内表面凹凸不平,外表面图显示大部分树种种壳外表面比较规则、平整,其中FC5、FC7表面非常光滑,其他树种表面有空隙,这对种子的吸水性也有一定影响。
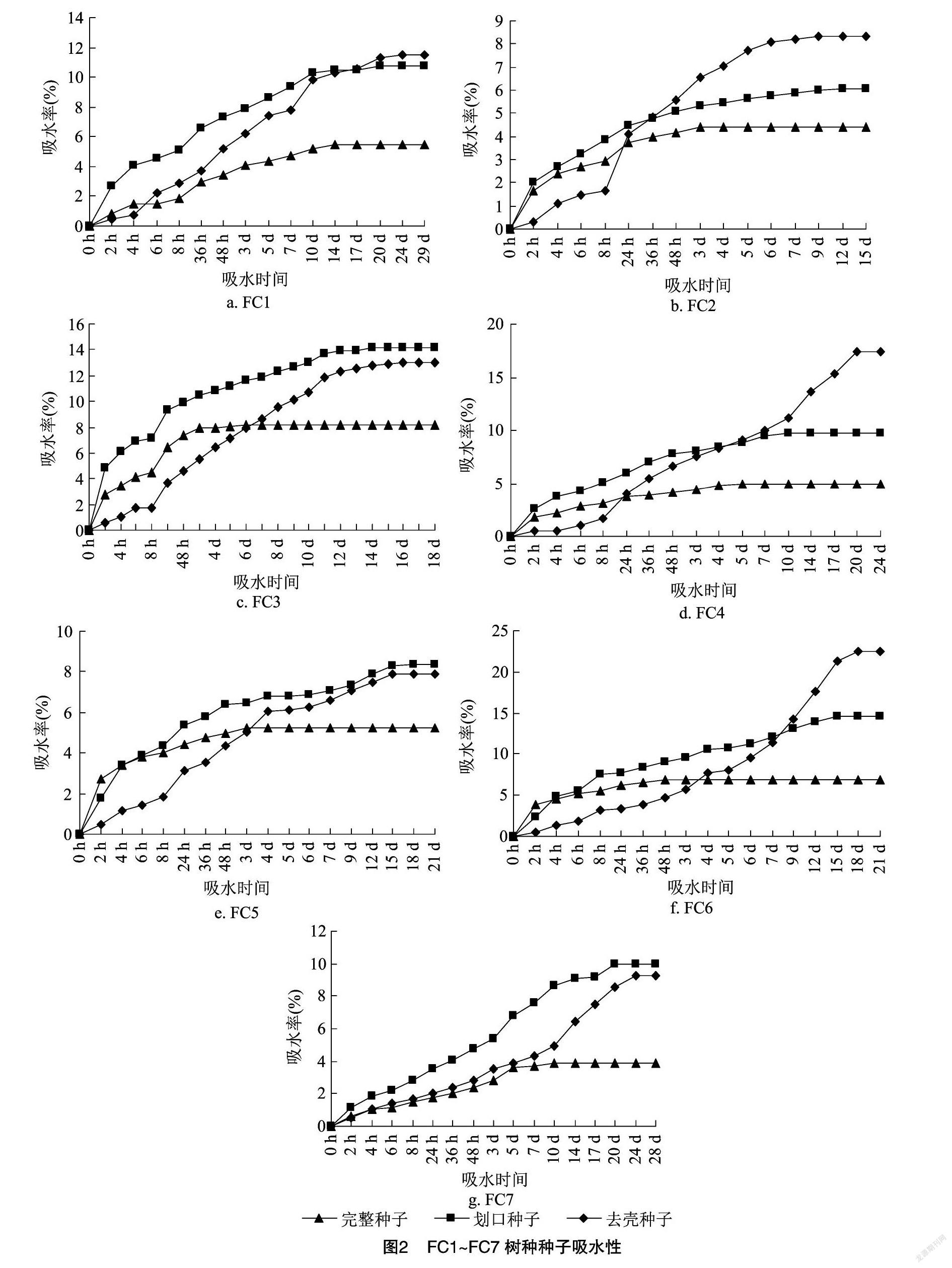
2.1.2 种壳吸水性
青冈属种子本身属于含水量较高的种子。从图2可以看出,对于所有树种,去壳种子和划口种子的最终吸水率均明显高于完整种子。完整种子吸水率最终达到5%左右,而去壳种子和划口种子大部分在10%左右,有的甚至超过20%。前8 h为快速吸水期,吸水率迅速上升,8 h后吸水率逐渐下降,最后趋于平缓。去壳种子可吸水时间最长,吸水率达到稳定的时间最晚。FC1、FC3、FC5、FC7的去壳种子和划口种子最终吸水率差异不明显,但均明显高于完整种子。FC2、FC4、FC6的去壳种子、划口种子和完整种子之间均差异明显,且去壳种子最终吸水率明显高于其他2个处理。可见种壳对种子吸水存在一定的阻碍作用。
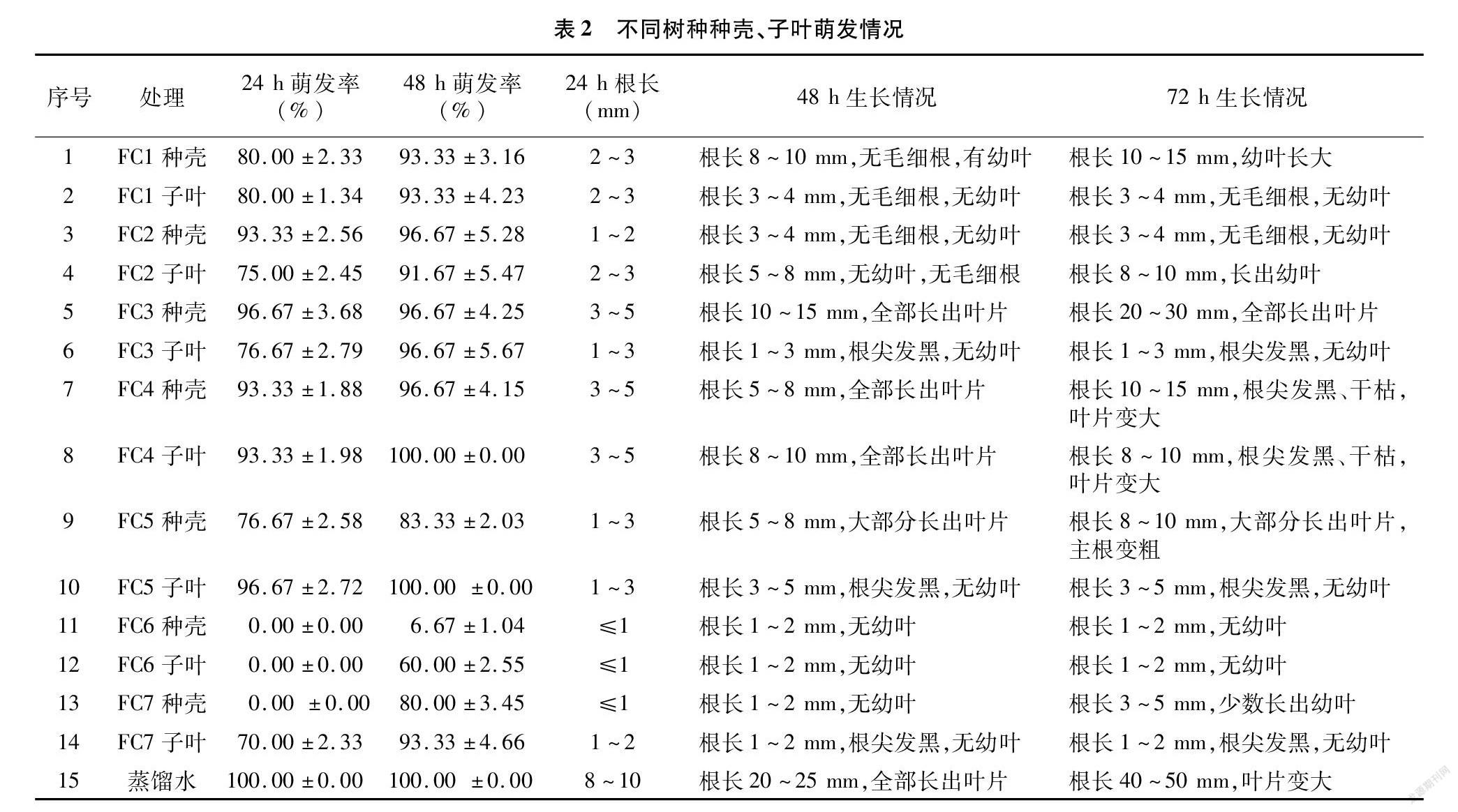
2.1.3 内源抑制物对白菜种子萌发影响 从表2可以看出,蒸馏水培养24 h后,白菜种子萌发率可达到100%,48 h后根长为20~25 mm,并长出叶片。经7个树种的种壳和子叶浸提液培养的白菜种子,除FC2、FC3、FC4种壳以及FC4、FC5子叶处理后的萌发率较高外,其他24 h萌发率均在80%及以下。其中FC6的种壳和子叶、FC7的种壳浸提液培养24 h后,萌发率为0,48 h后萌发率仍低于80%。培养72 h后,对照根长增加至40~50 mm,叶片明显增大,其他处理根长均无显著增加,且有几组处理出现根尖发黑、干枯的现象。经种壳和子叶浸提液培养后的白菜籽,仅有部分萌发出叶片,即使有叶片萌发,大部分根的长度也明显短于对照,根的生长状态也明显比对照差。由此可见,浸提液对白菜种子根的伸长有抑制作用,7个树种种壳及子叶内可能含有抑制种子萌发物质。不同树种种壳和子叶的抑制作用程度不同。FC1种壳和子叶浸提液培养的白菜种子萌发率一致,FC2和FC3子叶浸提液培养的白菜种子萌发率小于种壳,FC4、FC5、FC6和FC7种壳浸提液培养的白菜种子萌发率小于子叶。除FC2和FC6外,其他树种子叶浸提液培养的白菜种子72 h后根长均小于种壳,叶片生长情况也比种壳浸提液培养的白菜种子差。
2.2 休眠打破
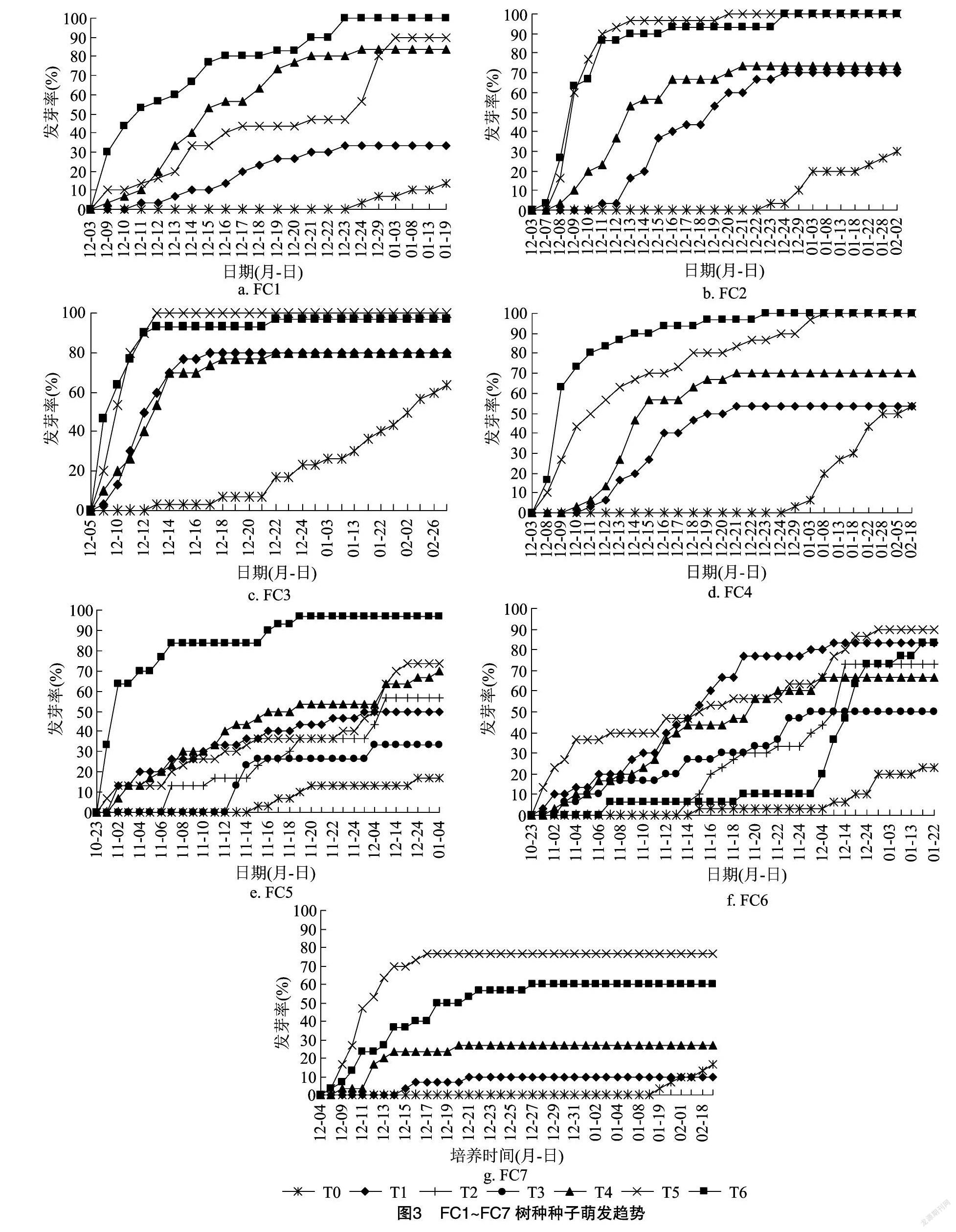
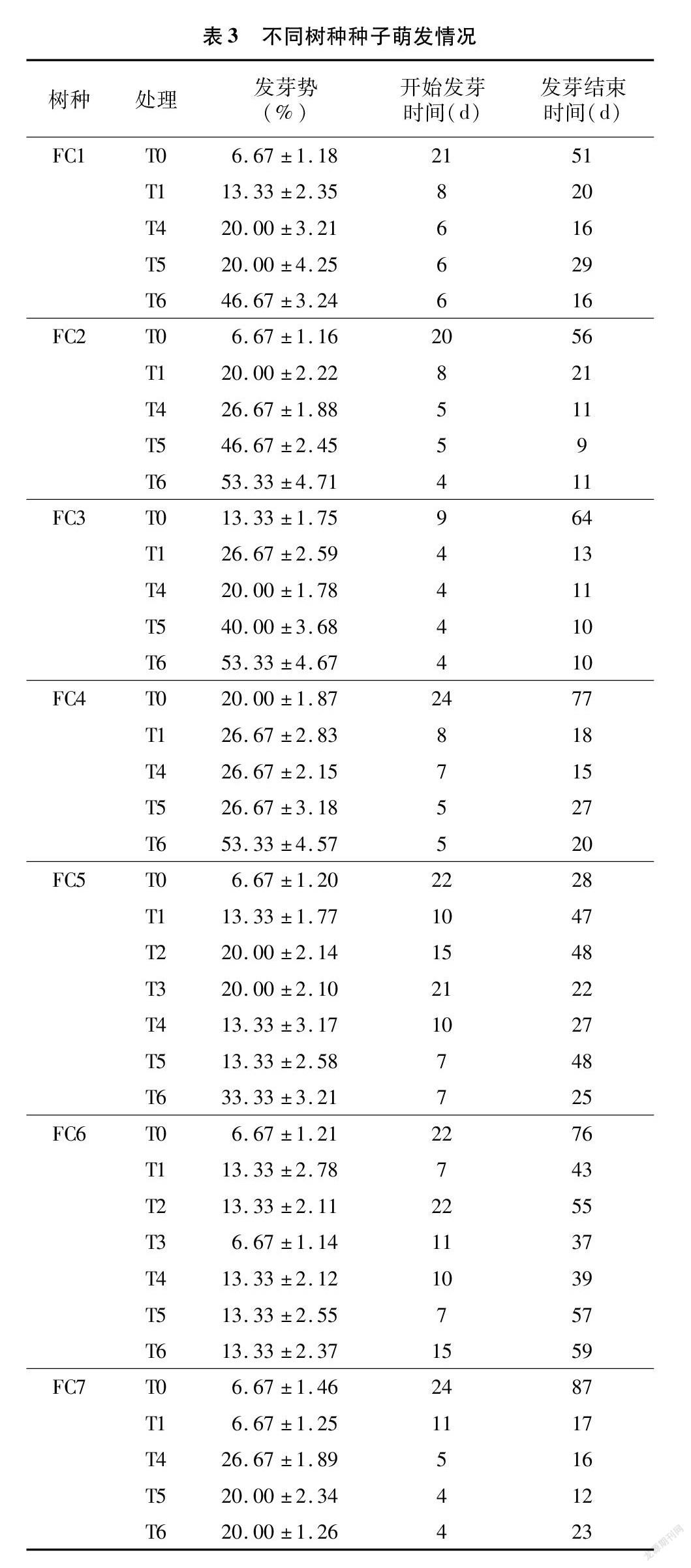
从图3、表3可以看出,大部分树种在T5、T6处理条件下萌发最早,一般比对照早15~20 d。除FC7外,其他树种在T6处理下最终发芽率均达80%以上,且FC1、FC2和FC4最终发芽率均为100%。FC1~FC5在T6处理下,发芽势均在30%以上,其中 FC2~FC4发芽势均高达53.33%。FC2、FC3、FC7等3个树种在T6处理下发芽最早,培养4 d后即开始发芽,其中FC3在T5处理下,仅8 d即全部萌发。FC5、FC6在大部分处理条件下种子萌发持续时间较长。
2.2.1 光照对解除青冈种子休眠的影响
从FC5的T2、T3处理试验结果可以看出,二者发芽势一致,均为20%。全黑暗条件下发芽较晚,但持续时间短,最终发芽率较低;而光照16 h条件下发芽较早,但持续时间长,最终发芽率较高。从FC6的T2、T3处理试验结果可以看出,二者发芽势都比较低,但T3处理下发芽时间明显早于T2处理,但最终发芽率明显低于T2处理。从这2个树种来看,光照时间16 h相较于全黑暗条件可以促进种子萌发。
2.2.2 培养温度对解除青冈种子休眠的影响
从FC5的T1、T2处理试验结果可以看出,二者最终发芽率一致,但与20 ℃相比,25 ℃条件下种子萌发更早,发芽势略低。从FC6的T1、T2处理试验结果可以看出,25 ℃条件下种子发芽明显早于20 ℃,二者的发芽势一致,但发芽率差异明显。从这2个树种来看,25 ℃ 相较于20 ℃,种子发芽更早,发芽率更高。因此25 ℃条件下更有利于休眠解除。
2.2.3 种子处理方式对解除青冈种子休眠的影响
对比T0、T1和T5处理可以发现,在7个树种中有5个树种在T5处理下最终发芽率均最高,有6个树种在T5处理下发芽均最早。而且FC2在T5处理下,种子在发芽后5 d时间内发芽率达到100%,FC3在T5处理下,种子在开始发芽后7 d时间内发芽率达到100%,发芽速度快。T0处理下发芽最晚,发芽周期一般也最长,发芽率除FC4外均是最低的。因此,相较于划口种子和完整种子,去种壳处理对休眠打破、提高发芽势和发芽率有极大的促进作用。
2.2.4 激素对解除青冈种子休眠的影响
2.2.4.1 划口种子 对于划口种子,对比所有树种T1和T4处理,除FC7外,使用400 mg/L GA3浸泡与使用常温水浸泡2种处理条件下种子开始萌发时间差异不大。除FC7外,其他树种在2种处理下的发芽势差异也不显著 ,但除了FC2和FC3外,其他树种在2种处理下的最终发芽率差异显著。因此对于划口种子,400 mg/L GA3對最终发芽率影响较大,但对种子开始萌发时间和种子发芽势影响不明显。
2.2.4.2 去壳种子 对比所有树种T5和T6处理可知,使用400 mg/L GA3浸泡与使用常温水浸泡对种子休眠解除和发芽率差异不明显,2种条件下种子开始萌发时间除了FC6外,其他树种差异均不大,发芽率除了FC5外,其他树种差异均不大。FC1、FC4、FC5等3个树种在T6处理下的发芽势均明显高于T5处理,且发芽周期也明显短于T5处理。
3 讨论与结论
大多种子休眠都是由多种原因造成的[1],可能是种皮结构致密或栅栏组织发达,也可能是种子表面角质层较厚或上覆蜡质[10-12]。周健等通过对种皮、胚乳细胞结构的观察和分析,发现阻碍种子吸水的主要原因是发达的栅栏层,揭示了大花四照花种子的休眠解除机制[13]。本研究中7种青冈属种子表面均有角质层,角质层细胞排列紧密,有保护种胚的作用[14]。然而表面蜡质和致密的角质层等种壳构造导致青冈属种子的吸水性很差,本研究也证明了去掉种壳后可以增加种子吸水速率,延长种子吸水饱和状态的时间。角质层里面是细胞紧密排列、细胞壁较厚的栅栏组织层,增加了种皮硬度。因此,角质层和栅栏组织层极有可能对胚的萌发有机械阻碍作用,影响胚根的萌发和伸长,同时也影响吸水性 这是种胚无法突破种皮进行正常萌发的原因之一。内表皮细胞排列相对疏松,可使水分迅速穿过,但有的树种内表皮细胞厚度也很大,在一定程度上也会阻碍种子萌发。青冈属种子结构中,种脐位置种壳最薄,且没有坚硬的角质层,应该是整个种子透水透气性最好的位置。但由于此次研究的7种植物胚芽均在种脐端相反位置,因而并未专门对种脐的吸水性进行研究。
植物体内产生抑制发芽物质或存在与抑制发芽相关的物质,此类物质对同种或异种种子萌发具有延迟或抑制作用[15]。朱铭玮等认为,酚类物质是凤丹种皮内的主要抑制物成分,有机酸类物质是凤丹种胚内的主要抑制物质成分[16]。栎类植物为防止昆虫捕食种子,产生了多酚(polyphenols)[17] 和单宁酸[18]等抑制种子萌发的次生代谢物。也有研究称栎属植物种子各部分甲醇浸提液都对白菜种子萌发、根长、苗高具有较强的抑制作用,其种皮、子叶、胚的浸提液对白菜种子的萌发抑制作用依次增强,而且对白菜种子萌发率的影响低于对根长、苗高的影响[19]。这与本研究的结果基本一致。本研究对子叶和种壳浸提液进行了对比,大部分树种子叶对白菜种子的抑制作用要比种壳强,且对根的伸长和叶片萌发的抑制作用要强于对发芽率的作用。说明青冈属种子中确实含有抑制白菜种子萌发的物质。但是试验中浸提液浓度很高,若进行稀释,是否还对白菜种子萌发有抑制作用,这些并未进行详细研究。而且自然界中的种子各部分中抑制物质的含量以及该含量是否可以抑制青冈种子萌发,也都需要进一步试验论证。
景美清等认为,内果皮、种皮的机械阻力和透气性是影响赤皮青冈种子休眠的主要原因,去除了内果皮、种皮后即可打破休眠。高浓度的GA3处理也有一定的作用[20]。本研究对比了光照时间、培养温度、种壳处理方式及激素等因素对休眠打破的影响,发现种壳机械处理对种子萌发影响最大,其他因素也有一定影响。本试验未对胚的形态进行研究,不能从形态结构上确定胚是否存在生理后熟。但种子在划口和去壳后迅速萌发,去壳后效果尤为明显,从侧面验证了胚不存在生理后熟,种壳对胚的机械阻碍是导致休眠的主要原因。
参考文献:
[1]郭聪聪,沈永宝,史锋厚. 白皮松种子休眠研究进展[J]. 南京林业大学学报(自然科学版),2019,43(2):175-183.
[2]Nikolaeva M G. Factors controlling the seed dormancy pattern[M]//Khan A A. The physiology and biochemistry of seed dormancy and germination. Amsterdam:North-Holland Publication Co.,1997.
[3]Baskin C C,Baskin J M. Seeds:Ecology,biogeography and evolution of dormancy and germination[M]. 2nd ed. San Diego:Academic Press,2014.
[4]孟慶伟,高辉远. 植物生理学[M]. 北京:中国农业出版社,2011:317-318.
[5]耿文娟,冯贝贝,梅 轩,等. 野生欧洲李种子萌发特性[J]. 经济林研究,2017:35(1):20-25.
[6]仇云云,崔 健,刘 雪,等. 紫斑牡丹种子休眠原因初探[J]. 浙江农林大学学报,2018,35(3):497-504.
[7]李金华. 赤皮青冈种子萌发及幼苗生长调控技术研究[D]. 长沙:中南林业科技大学,2017.
[8]彭颖姝,李铁华,文仕知,等. 不同沙藏处理对青冈栎种子萌发的影响[J]. 中南林业科技大学学报,2016,36(8):44-48.
[9]李庆梅,刘 艳,刘广全,等. 栎属7种植物种子的发芽抑制物质研究[J]. 生态学报,2013,33(7):2104-2112.
[10]周 健,苏友谊,代 松,等. 紫荆种子成熟过程中种皮和胚乳超微结构观察[J]. 南京林业大学学报(自然科学版),2016,40(6):27-32.
[11]杨万霞,洑香香,方升佐. 青钱柳种子的种皮构造及其对透水性的影响[J]. 南京林业大学学报(自然科学版),2005,29(5):25-28.
[12]Greipsson S. Effects of stratification and GA3 on seed germination of a sand stabilising grass Leymus arenarius used in reclamation[J]. Seed Science and Technology,2001,29 (1):1-10.
[13]周 健,代 松,钱存梦,等. 大花四照花种子休眠解除过程中种皮和胚乳结构变化[J]. 东北林业大学学报,2014,44(5):34-38.
[14]钱领元,施拱生. 乌桕籽“蜡被”形成过程的研究[J]. 浙江林学院学报,1986,3(1):1-5.
[15]Evenari M. Germination inhibitors[J]. The Botanical Review,1949,15(3):153-194.
[16]朱铭玮,邹雨婷,李永荣,等. 油用牡丹‘凤丹’种子内源抑制物研究[J]. 西南林业大学学报,2019,39(6):64-70.
[17]Rakic S,Povrenovic D,Tesevic V,et al. Oak acorn,polyphenols and antioxidant activity in functional food[J]. Journal of Food Engineering,2006,74(3):416-423.
[18]Shimada T. Nutrient compositions of acorns and horse chestnuts in relation to seed-hoarding[J]. Ecological Research,2001,16(4):803-808.
[19]李庆梅. 几种北方落叶栎种子发育的形态生理及萌发特性研究[D]. 北京:北京林业大学,2013.
[20]景美清,李志辉,杨模华,等. 赤皮青冈种子质量与萌发特性研究[J]. 中国农学通报,2012,28(34):27-30.
