嗜酸硫杆菌在工农业中的应用
2022-06-10高雪彦陈林旭陈显轲庞昕潘登林建群
高雪彦 陈林旭 陈显轲 庞昕 潘登 林建群
(1. 山东第一医科大学(山东省医学科学院)医学科技创新中心,济南 250000;2. 山东大学微生物技术国家重点实验室,青岛 266237; 3. 山东亿安生物工程有限公司,济南 250000)
嗜酸硫杆菌(属)(Acidithiobacillus spp.)是一类嗜酸性专性化能自养细菌,能从亚铁、硫或还原性硫化物(reduced inorganic sulfur compounds,RISCs)的氧化中获得生长所必需的能量和还原力,以卡尔文循环固定空气中的二氧化碳为碳源,实现自养生长[1-3]。嗜酸硫杆菌广泛存在于陆地或海洋的酸性环境中,作为地质微生物的重要成员,因其亚铁和硫代谢的生理特点参与地球硫和铁元素循环,促进酸性环境的产生和酸性微生态系统的形成[4-5]。随着微生物分离培养技术的进步和生物信息学的发展,越来越多的嗜酸硫杆菌被发现、分离并鉴定。根据NCBI数据库上的信息,目前已经鉴定的嗜酸硫杆菌有9种,包括A. ferrooxidans、A. ferrivorans、A. ferridurans、A. ferriphilus、A. ferrianus、A. thiooxidans、A. caldus、A. albertensis和A. sulfuriphilus,这9种嗜酸硫杆菌都具有硫氧化的能力,其中A. ferrooxidans、A. ferrivorans、A. ferridurans、A. ferriphilus和A. ferrianus这5种细菌,既具有亚铁氧化的能力也具备硫氧化的能力,部分嗜酸硫杆菌还具有代谢氢的能力 (表1)[6-14]。
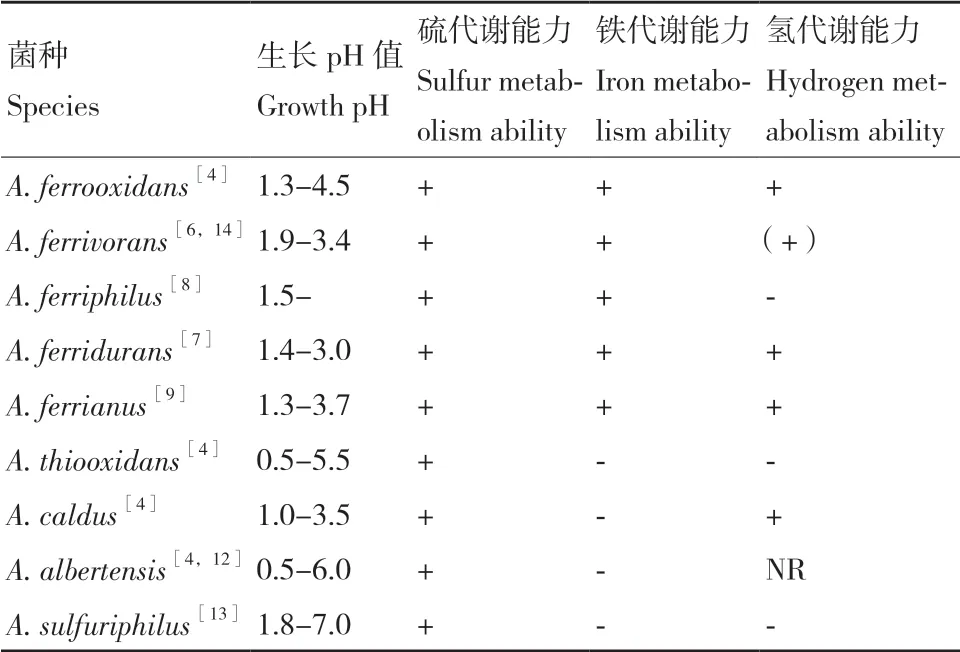
表1 九种嗜酸硫杆菌基本生理代谢特征Table 1 Physiological and metabolic characteristics of 9 species of Acidithiobacillus spp.
近年来,由于嗜酸硫杆菌独特的硫和亚铁代谢能力以及对重金属离子和酸性环境较强的适应能力,使其在工农业中的应用价值越来越显著[15-16]。本文综述了嗜酸硫杆菌基本的生理代谢特点和环境适应机制,介绍了嗜酸硫杆菌在金属矿产资源的开采、重金属的资源回收、重金属环境污染治理、生物脱硫以及土壤修复等方面的应用,结合社会需求和嗜酸硫杆菌的研究现状,分析了嗜酸硫杆菌在理论基础和应用实践中亟待解决的重要科学问题,助力嗜酸硫杆菌理论基础和产业应用的研究。
1 嗜酸硫杆菌生理代谢特点
1.1 嗜酸硫杆菌硫代谢机制
元素硫是构成生命体重要组成元素,具有-2到+6多种价态形式,从而产生了多种多样的含硫化物,包括连四硫酸盐(S4O62-)、硫代硫酸盐(S2O32-)、亚硫酸盐(SO32-)、硫化物(S2-)、硫酸盐(SO42-)和元素硫(S0)[4]。嗜酸硫杆菌可以高效氧化各种还原态的无机硫化合物(RISCs)以获得用于自养生长的能量,因而成为地球硫元素循环的重要组成部分。嗜酸硫杆菌具有复杂的硫代谢酶系,如元素硫氧化酶、硫代硫酸盐氧化途径中的酶、亚硫酸盐氧化酶等,这些酶分布在细胞膜、细胞周质、细胞质等不同的区域协同作用,将单质硫或还原的无机硫化合物氧化为最终产物硫酸盐[11,17-19]。
硫代谢过程涉及到复杂的酶促反应,根据在细胞中的反应区域可以划分为细胞外膜、细胞周质、细胞内膜和细胞质几个部分。首先,细胞外的元素硫被分布在细胞外膜上的蛋白(outer-membrane proteins,OMP)激活转运,进入周质空间[11,17]。单质硫活化转运是硫代谢过程中至关重要的环节,据报道,嗜酸硫杆菌以单质硫为能源培养时延迟期时间比较长,细胞生长缓慢,推测这一时期单质硫被活化转运。其次,在细胞周质空间,硫氧化酶系统(sulfur oxidizing enzyme system,Sox)发挥了重要作用[4]。典型的Sox系统由SoxYZ、SoxXA、SoxB和Sox(CD)2四种蛋白组成,二聚体蛋白SoxYZ在硫代谢过程中具有结合硫化物的作用,SoxXA能够催化氧化还原反应并获得电子[4]。Sox系统的4个组成蛋白形成一个循环反应,具有将单质硫、硫代硫酸盐、亚硫酸盐和硫化氢等还原性硫化物氧化成硫酸盐的能力,硫代硫酸盐氧化是该氧化途径的核心[20]。Sox系统以多种形式在部分嗜酸硫杆菌中存在,研究发现A. caldus,A. thiooxidans和A. albertensis中含有缺失了Sox(CD)2蛋白的两套不完整的Sox系统,Sox I和Sox II,分别由soxYZBhyp-resB-soxAX-resC和soxXYZA-hyp-soxB基因编码的蛋白构成。在Sox II系统基因簇的上游发现了依赖于sigma54转录因子的的双组分系统tspS-tspR[21]。具有硫和亚铁双能量代谢途径的A. ferrooxidans细菌缺失Sox系统[18-19]。然而,在A. ferrooxidans中存在硫代硫酸盐脱氢酶(thiosulfate dehydrogenase,TSD),推测可能是周质中存在的硫代硫酸盐代谢途径[4,19]。此外,由连四硫酸盐水解酶(tetrathionate hydrolase,TetH)和硫代硫酸盐-辅酶Q氧化还原酶(thiosulfate quinone oxidoreductase,TQO)介 导的连四硫酸盐和硫代硫酸盐的转化反应也是细胞周质空间中重要的硫氧化反应过程[4,18]。在细胞内膜和胞质中同样存在具有硫氧化作用的蛋白酶,如定位于内膜的硫化物-辅酶Q氧化还原酶(sulfide:quinone oxidoreductase,SQR)、HDR复合体(hdr-like complex),位于胞质中的硫加氧还原酶(sulfur oxygenase reductase,SOR)以及硫氰酸酶(rhodanese,RHD)等,部分氧化酶与辅酶Q相连,传递硫氧化过程中产生的电子,获得能量[11,17-18]。
由于元素硫的价态比较多,硫化物的种类繁杂、多样,因此嗜酸硫杆菌的硫代谢机制比较复杂,涉及到分布在不同细胞区域的几十种活性蛋白的协同作用。目前文献报道的嗜酸硫杆菌硫代谢模型大部分都是基于基因测序技术和生物信息学的模拟推测,尚有很多未解之谜值得继续深入研究。
1.2 嗜酸硫杆菌亚铁代谢途径
目前发现的9种嗜酸硫杆菌中,A. ferrooxidans、 A. ferrivorans、A. ferriphilus、A. ferridurans和A. ferrianus具有氧化亚铁获得能量和电子的能力。
早在20世纪80年代,Ingledew等[22]提出了亚铁氧化电子传递链的模型,提出了电子传递的分叉途径。经过几十年的研究,越来越多的亚铁氧化电子传递链中的传递体蛋白功能被证实。随着组学技术的进步和生物信息学的发展,2009年Quatrini等[18]通过基因差异表达转录谱和RT-qPCR实验,结合生物信息学分析提出了A. ferrooxidans 23270的亚铁氧化电子传递链模型,该电子传递链模型中存在顺电势和逆电势两条电子传递途径。
在顺电势电子传递途径中,亚铁离子氧化失去的电子被镶嵌在外膜上的细胞色素蛋白Cyc2传递给细胞周质中的铜蓝蛋白Rus,途经细胞色素蛋白Cyc1、细胞色素氧化酶aa3,传递给最终电子受体O2,并与质子结合生成H2O,同时形成跨膜质子动势,驱动ATP合成[18-19]。因此,顺电势电子传递途径中电子传递的顺序可以简要概括为Fe2+→Cyc2→Rus→Cyc1→ aa3→O2。顺电势电子传递途径中电子传递体蛋白由rus操纵子编码,rus操纵子包含8个编码基因,cyc2-cyc1-cup-coxBcoxA-coxC-coxD-rus[23-24]。研究表明,rus操纵子在以亚铁为能源时转录水平显著上调,表明了rus操纵子在亚铁代谢中的重要作用[24-25]。rus操纵子中,Cyc2位于细胞外膜上,是电子传递的第一个受体蛋白,在亚铁氧化电子传递模型中具有至关重要的作用。铜蓝蛋白Rus位于细胞周质空间,是顺电势和逆电势电子传递链中的分支蛋白[25]。
在逆电势电子传递途径中,以Rus为分支,电子经Cyc2、Rus 传递给细胞色素蛋白CycA1、细胞色素bc1复合体、泛醌、NADH-Q还原酶,最后传递给NAD(P)+,生成NAD(P)H,提供细胞固定CO2和参与有氧代谢所需的还原力,而A. ferrooxidans产生还原力的这一途径是耗能的。逆电势电子传递途径中电子传递体蛋白由pet I操纵子编码,pet I操纵子包含5个编码基因,cycA1-sdrA1-petA1B1C1[18,23]。
1.3 嗜酸硫杆菌氢代谢特点
部分嗜酸硫杆菌能够在H2的环境中生长,氧化H2获得质子和电子,O2或者Fe3+可以作为氢氧化的电子受体[7,26]。然而,目前关于嗜酸硫杆菌利用氢的报道比较少。
氢化酶(hydrogenase)作为参与氢代谢的酶广泛存在于细菌,古细菌和一些真核生物中。氢化酶是一组金属酶,许多纯化的氢化酶的催化活性是双向的,能够催化H2转化为质子和电子以及生成H2的逆向反应,反应方向取决于细胞内的生理环 境[27-28]。20世纪90年代,研究者在A. ferrooxidans中发现并纯化了氢化酶,序列分析表明A. ferrooxi- dans细菌的氢化酶与蓝藻的氢化酶同属于一个蛋白家族[29]。在结构上,A. ferrooxidans细菌的氢化酶是由64 kD的大亚基(HoxG)和34 kD的小亚基(HoxK)构成的异源二聚体,HoxG大亚基中独特的[NiFe]簇被认为是氢化酶的催化活性中心,[NiFe]活性位点的结构在所有[NiFe]氢化酶中非常相似。HoxK小亚基含有几个Fe-S簇,具有传递电子的作用[29-30]。 虽然嗜酸硫杆菌生活在酸性环境中,但是实验发现纯化的A. ferrooxidans细菌氢化酶最佳反应pH值为9[30]。当以氢气为培养条件时,A. ferrooxidans有氧生长,以O2为电子受体,生长速率慢,无氧生长则以Fe3+作为氢氧化的电子受体[7,26]。Gao等[31]在研究嗜酸性氧化亚铁硫杆菌群体密度感应系统时发现了一组与氢化酶和电子传递相关的涉及20个基因的基因簇(AFE_0700-AFE_0719)。其中,AFE_0701和AFE_0702两个基因在基因组上的注释分别是氢化酶的小亚基和大亚基(图1)。研究发现,该基因簇在以亚铁为能源培养时基因表达差异显著,推测在A. ferrooxidans中亚铁代谢与氢代谢之间存在某种联系,尚需进一步的实验证实。
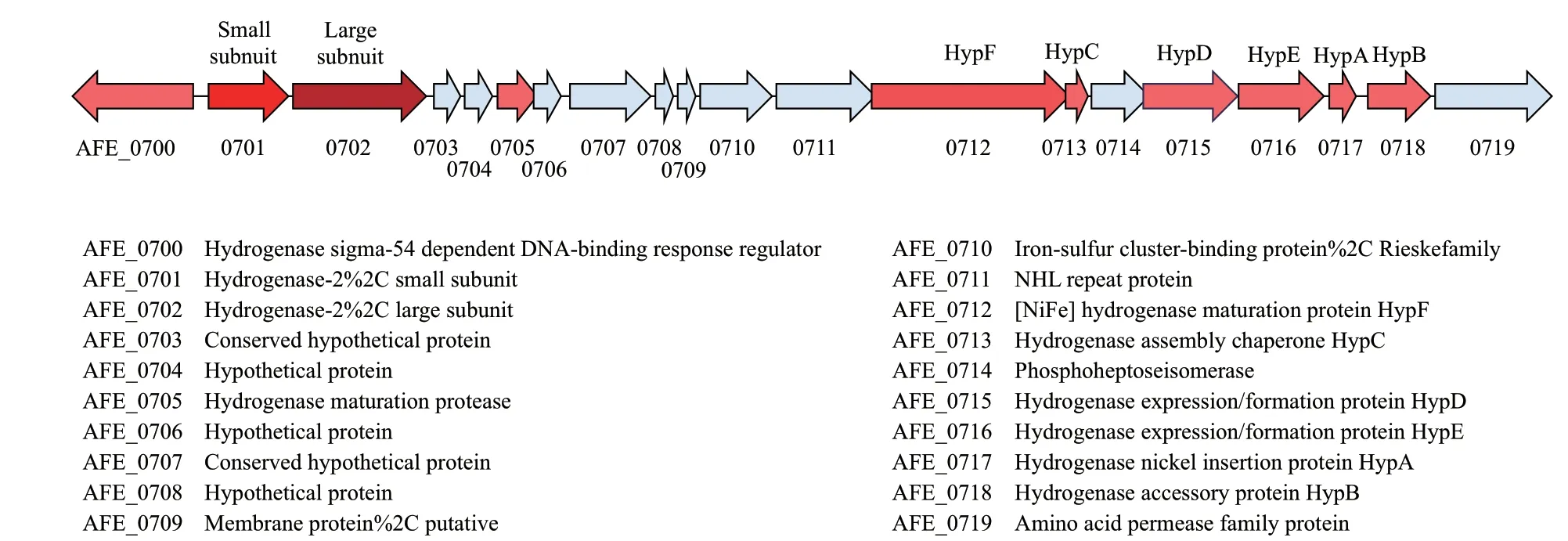
图1 氢化酶相关基因簇(AFE_0700-AFE_0719)Fig. 1 Hydrogenase related gene cluster(AFE_0700-AFE_0719)
2 嗜酸硫杆菌对极端环境的适应机制
嗜酸硫杆菌作为极端环境微生物的重要成员,在长期共同进化和自然选择中衍生出了对极端环境的特殊适应能力,充分了解嗜酸硫杆菌属的生长和环境适应机制对于理解菌株的生态行为和嗜酸硫杆菌的应用价值具有重要意义。
2.1 嗜酸硫杆菌群体感应系统与生物膜
群体密度感应(quorum sensing,QS)是一种普遍存在的现象,它使细菌能够通过产生、分泌和检测信号分子(也称为自诱导分子)感知群体密度和外界环境,建立细胞间的交流[32-33]。QS系统合成和分泌的信号分子种类繁多。其中N-酰基高丝氨酸内酯(acyl-HSL)是革兰氏阴性细菌中最常见的QS系统信号分子,分子结构上由高丝氨酸内酯环和酰胺侧链组成[32]。QS系统的研究已有50、60年的历史,近年来,借助生物信息学、分子生物学和化学分析等现代分析实验技术,越来越多的QS系统的信号分子、调控机制和功能被发现,然而相比于病原微生物和其他模式细菌,由于分子操作技术的限制,嗜酸硫杆菌中QS系统的研究比较局限。
2005年,Farah等[34]在A. ferrooxidans中发现了LuxI/R型QS系统(AfeI/R)。AfeI/R型QS系统包含3个编码基因,afeI-orf3-afeR。AfeI属于LuxI蛋白家族,以S-腺苷甲硫氨酸(SAM)和来自于脂肪酸代谢过程的acyl-ACP为底物合成acyl-HSLs类型的信号分子,在亚铁、硫和硫代硫酸盐能源底物培养条件下,A. ferrooxidans能够合成9种acyl-HSLs。AfeR是LuxR家族蛋白,具有识别和结合信号分子以及调节基因表达的功能,位于AfeI和AfeR之间的Orf3的功能目前未知[34-35]。生物信息学分析表明,AfeI/R型QS系统在嗜酸硫杆菌中的分布具有普遍性和特殊性,除了A. ferrianus和A. ferriphilus,AfeI/R型QS系统存在于其他具有硫和亚铁两种能量代谢途径的嗜酸硫杆菌中,而仅具有单一能源(硫)代谢途径的嗜酸硫杆菌中,只有在A. thiooxidans中发现了AfeI/R型QS系统[31]。此外,AfeI/R系统在A. ferrooxidans中依赖于能源底物的调节机制被揭示,A. ferrooxidans根据所处的能源底物环境(硫和亚铁能源底物),合成不同类型和浓度的acyl-HSLs,有针对性的调控硫代谢和亚铁代谢以及其他与环境底物相适应的遗传表型,反映了QS系统在嗜酸硫杆菌与能源环境之间的调控和适应机制[36]。
2007年,Rivas等[37]在A. ferrooxidans中发现了另一套QS系统(QS-II)。该QS系统包括4个共转录基因,glyQ、glyS、gph和act,分别编码甘氨酸t-RNA合成酶的α和β亚基、磷酸酶和酰基转移酶。在大肠杆菌中异源表达Act蛋白可以检测到C14-HSL。半定量RT-PCR实验表明act基因以亚铁为能源底物培养时表达量高于以硫为能源底物培养。然而,与Act对应的信号分子受体调控蛋白尚未发现,Act是否能在A.ferrooxidans中合成acyl-HSLs或其他类型的信号分子,至今未见报道。因此,Act介导的QS系统仍存在争议[38]。
许多嗜酸硫杆菌如A. ferrooxidans、A. ferrivorans和A. thiooxidans生长代谢受到acyl-HSLs添加的影响[38-39],而这些影响嗜酸硫杆菌生长代谢的acyl-HSLs并不一定是由菌株本身合成。人工合成的四唑类似物也能够刺激A. ferrooxidans基因的差异表达[40]。可以推测,嗜酸硫杆菌存在种间信号的交叉识别和通讯,相关的研究工作有待深入开展。
QS系统是调节细胞外聚合物(extracellular polysaccharides,EPS)合成和生物膜(biofilm)形成的重要途径[41]。生物膜是微生物在极端环境下“抱团”的一种群体行为。基于胞外多聚物形成的生物膜导致细胞聚集和特殊生态微环境的形成,有利于嗜酸硫杆菌在营造的微环境中生长繁殖,加速矿物氧化,抵制不良环境[42]。激光共聚焦显微镜和原子力显微镜观察到,添加acyl-HSLs可以影响黄铁矿表面嗜酸硫杆菌生物膜的形成[39,42]。转录组数据表明,酰基高丝氨酸四唑类似物添加刺激了100多个 A. ferrooxidans基因的差异表达,其中42.5%的差异表达基因与生物膜合成有关[40]。QS系统的过表达不仅可以促进EPS的合成和生物膜的形成,还可以增加细胞的硫氧化能力,增强A. ferrooxidans细胞对硫的侵蚀作用,提高细菌对金属硫化矿的氧化和金属离子的浸出效率[39,42]。
2.2 嗜酸硫杆菌对重金属离子的抗性及其分子 机制
生物冶金是嗜酸硫杆菌在工业上的一项重要应用,嗜酸硫杆菌通过直接或间接作用将矿石中的各种金属溶解到浸出液中,浸出体系中金属离子的浓度越来越高,成为制约微生物生长代谢和浸矿效率的重要环境压力,因此研究嗜酸硫杆菌对重金属离子的抗性及其分子机制是生物冶金应用领域的迫切需求[16]。
当重金属离子浓度过高而对微生物产生毒害作用时,微生物可以通过转运、螯合等多种方式降低重金属的毒害作用[43-44]。嗜酸硫杆菌在与重金属环境的适应进化中衍生出了高效的抗毒和解毒机制。其一是胞外吸附,嗜酸硫杆菌合成和分泌EPS,形成生物膜,吸附和抵制胞外重金属离子。EPS主要由蛋白质、多糖和脂类组成,通过与重金属离子建立共价连接或静电相互作用从而达到很好的吸附效果[43]。其二是离子外排,嗜酸硫杆菌可以利用金属离子外排系统将金属离子泵出到胞外,如金属离子外排P型ATPase家族蛋白,水解ATP提供能量,将金属离子逆浓度梯度主动排出到胞外,达到解毒的效果[44]。其三是胞内螯合,细胞内的一些氨基酸、金属结合蛋白、磷酸盐等组分通过螯合胞内的金属离子降低胞内金属离子的浓度。研究发现A. ferrooxidans能够积累多聚磷酸盐(polyphosphate,polyP)颗粒,polyP颗粒在多种细菌中都被证实具有供能和“隔离”重金属离子的双重功能[45]。在重金属离子浓度过高时,polyP水解生成的无机磷酸与重金属离子结合形成复合物,通过磷酸盐转运系统将重金属离子排出到细胞外[45]。此外嗜酸硫杆菌还可以通过生物转化将毒害作用大的重金属离子转化为毒性小或无毒的成分、建立金属离子渗透屏障、降低细胞重金属离子靶标蛋白的敏感性等多种方式实现对重金属的抗毒和解毒作用[46]。
2.3 嗜酸硫杆菌的酸耐机制
嗜酸硫杆菌能够在酸性环境下生长繁殖(pH 0.5-6.0),9种嗜酸硫杆菌生长的pH范围在表1中列出。嗜酸硫杆菌能够在外部酸性条件下维持相对中性的细胞内环境,避免酸性环境对胞内蛋白质、DNA等生物大分子的损伤。嗜酸硫杆菌在与酸环境漫长的适应过程中演化出多种多样的耐酸响应机制和调控策略包括,细胞膜组分和蛋白通道的调节、质子外排和消耗、反向跨膜电位的产生、DNA和蛋白质修复等[47-48]。
细胞膜是嗜酸硫杆菌应对酸环境的屏障,嗜酸硫杆菌能够合成高度不透性的细胞膜以应对胞外高浓度的质子。这些膜由大量的支链脂肪酸、饱和和单不饱和脂肪酸、β-羟基、ω-环己基和环丙烷脂肪酸组成[49]。位于细胞膜上的蛋白通道在酸环境下其通道大小和离子选择性受到调节,以应对外界质子胁迫。其次,嗜酸硫杆菌进化出高效的质子外排能力,质子外排是涉及细胞能量的复杂系统。目前在嗜酸硫杆菌如A. ferrooxidans、A. thiooxidans和A. caldus等细菌基因组中鉴定出了几种质子外排蛋白[50]。在酸胁迫下,嗜酸硫杆菌中的H+-ATPase活性和NAD+/NADH比率上调[20,48]。呼吸链复合体蛋白的表达增加,这些复合物可以将质子泵出细胞[20]。当胞质中质子浓度过高时,细菌可以利用氨基酸脱羧酶催化脱羧反应消耗胞内质子,在一定程度上维持细胞内pH值稳态。此外,嗜酸硫杆菌能够利用阳离子转运蛋白(如Na+/K+转运蛋白)产生反向跨膜电位(Δψ)以抑制质子向胞内流动[20,48]。研究数据表明,A. thiooxidans的基因组中含有大量的阳离子转运蛋白基因,这些转运蛋白可能参与反向跨膜电位的形成以抑制质子流入[20,50]。当细胞暴露于高浓度的质子环境或质子流入细胞时,大量的DNA修复蛋白和分子伴侣会修复受损的DNA和蛋白。
酸环境是嗜酸硫杆菌在生物冶金工业应用中的重要条件,是矿石中金属溶解浸出的关键因素。在过去几十年中,嗜酸硫杆菌的分子和生化分析与基因组、转录组、蛋白质组等多组学数据相结合,为嗜酸硫杆菌的耐酸机制提供了见解。然而,目前一些pH稳态机制和耐酸模型仍缺乏严格的实验证据,需要深入的研究和挖掘。
3 嗜酸硫杆菌在工农业中的应用
我国“十四五”规划明确指出的“绿色发展”理念,“绿水青山就是金山银山”的科学论断也体现出人与环境和谐共存的内在需求。微生物是联系人类与环境的重要纽带,嗜酸硫杆菌在重金属矿产资源开采、有色金属回收、重金属污染物治理、生物脱硫、土壤环境改善等领域具有重要作用(图2),是实现资源高效转化和环境污染治理不可或缺的微生物资源。

图2 嗜酸硫杆菌在工农业中的应用Fig. 2 Application of Acdithiobacillus spp. in industry and agriculture
3.1 嗜酸硫杆菌在工业中的应用
生物冶金是嗜酸硫杆菌在工业中的重要应用,该技术是利用微生物的直接或间接作用将矿石中的各种金属溶出,并加以回收利用[16,51]。嗜酸硫杆菌是生物冶金中的优势菌株,通过其独特的硫和亚铁代谢能力,利用矿石中的亚铁和硫化物生长,产生Fe3+和硫酸,Fe3+是参与生物冶金的关键因素,可以作为氧化剂攻击金属,自身被还原为Fe2+,产生的酸性环境有利于矿石的溶解和金属离子的浸 出[15,51]。嗜酸硫杆菌能够从低品位矿石中浸出铜、锌、镍、钴等金属[16,51]。据报道,在伊朗51.0%的锌被A. ferrooxidans细菌从复杂的低品位锌铅矿石中浸出[52],此外,A. ferrooxidans也被用于从低品位的硫化矿中提取钴,浸出效率达62%[53]。由A. ferrooxidans、A. thiooxidans以及其他几种浸矿微生物组成的菌种能够从低品位矿中回收70%的铜[54]。此外嗜酸硫杆菌可以利用生物氧化作用将金矿石中的硫化物等组分氧化分解从而使金裸露而易于提取。越来越多的研究证实与传统的冶炼技术相比,嗜酸硫杆菌参与的生物冶金过程在处理低品位矿、尾矿中具有明显的优势。
近年来,嗜酸硫杆菌的应用已从重金属矿产资源开采拓展到含金属废弃物的资源回收[55]。随着科技的进步,电气和电子设备(electrical and electronic equipment,EEE)已成为生活中必不可少的一部分。电子产品更新换代导致废旧手机、电脑、电视、电池等电子废弃物的大幅增加,2020年全球电子垃圾监测报告指出上一年度全球产生了53.6 Mt电子垃圾[56]。这些电路板、电池等废弃物中含有金、银、锂、钴、铜、铅等多种稀有金属和贵重金属[15],是放错了地方的资源。嗜酸硫杆菌能够有效回收利用这些电子废弃物中的各种金属,避免电子垃圾带来的环境污染,提高资源利用效率。在最佳条件下A. thiooxidans在72 h浸出时间内能从废旧手机印刷电路板中回收98%的铜和82%的镍[57]。通过逐步间接生物浸出法,在A. ferrooxidans的作用下,废旧发光二极管中镓、铜和镍浸出率分别提高到83%、97%和84%[58]。
随着社会的进步,皮革厂、电镀厂、畜禽粪便等污水、污泥废弃物排放量与电子垃圾同样急剧增加,第二次全国污染源普查公报显示,2017年我国工业固体危险废弃物产生量达6 581.45万t[59],这些废弃物中通常含有铬、铅等重金属,直接排放到环境中会造成重金属在食物链中积累,破坏生态环境,危害人类健康。嗜酸硫杆菌由于自身的生理代谢特点和较高的重金属离子耐受能力而被广泛应用于含重金属的废弃物治理中。利用A. ferrooxidans处理焚烧后的垃圾残渣,在同时含有亚铁和元素硫的介质中培养时,锰、钴、铜和锌的浸出效率高达100%[60]。来自天津当地市政污泥处理厂的污泥,在经过10 d的亚铁氧化细菌生物浸出反应后其锌、铜、镉、铬、锰、镍和铅得到一定程度的浸出和去除[36]。A. ferrooxidans和A. thiooxidans被报道能够从炼油厂废弃物回收铝、钴、钼和镍[61]。
生物脱硫(biodesulfurization)是嗜酸硫杆菌在污染物治理中的另一项重要应用,生物脱硫是利用微生物自身的生理代谢实现硫化物脱除的技术,广泛应用在石油脱硫、煤炭脱硫、沼气脱硫等领域,以减少SO2的排放和对环境的影响[62-64]。据报道A. ferrooxidans和A.thiooxidans细菌能够有效去除煤炭中黄铁矿硫和总硫的含量[63,65]。Cheng等[62]将A. ferrooxidans细菌固定化,对高浓度的H2S气体[(15 000±100)ppmv]的脱除率达到80%。
3.2 嗜酸硫杆菌在农业中的应用
嗜酸硫杆菌在农业上的应用与其在工业中的应用相比研究的较少,但是嗜酸硫杆菌与农业应用的联系由来已久。嗜酸硫杆菌中的A. thiooxidans细菌是20世纪20年代从堆肥中分离得到的[66]。20世纪60、70年代,针对我国北方土壤缺磷的现状,通过对A. thiooxidans的深入研究,利用A. thiooxidans和硫磺、磷矿粉、硫酸铵、麦糠或碎秸秆制成的堆肥可以提高磷矿粉的速效性,促进小麦和玉米的生长,提高农作物的产量。基于嗜酸硫杆菌能够产酸的特点,降低土壤pH值,改善盐碱地的土壤环境[10]。随着土壤pH值降低,土壤酸化,有利于土壤中一些不溶性矿质盐的溶解,土壤中的硫酸盐和可溶性的钾、钙、磷等元素含量增加,从而改善了作物的矿质营养[9]。此外,嗜酸硫杆菌能够有效浸出重金属污染土壤的铬、镉、铅、铜等重金属实现土壤修复的目的[67]。
4 总结与展望
嗜酸硫杆菌是资源环境微生物研究与微生物工业应用有机结合的典范,其独特的能量代谢和极端环境适应机制不仅是微生物学研究中的重要科学问题,也是它们具有重要工农业应用价值的核心。基于前期的研究成果,为更好的解决我国资源与环境问题,服务国家需求,嗜酸硫杆菌的研究在以下四个方面需要重点布局。
第一、能量代谢机制研究。嗜酸硫杆菌具有硫、亚铁、氢多种能量代谢途径,能量代谢是细菌改造和应用研究的基石。目前嗜酸硫杆菌的代谢途径大多是基于序列信息和生物信息学分析的模型,缺乏实验证据。通过完善微生物细胞的分子代谢网络、关键酶及代谢途径中代谢物的调控作用,加深理论研究解答嗜酸硫杆菌生存之谜。
第二、极端环境抗逆和适应机制研究。嗜酸硫杆菌能够在酸、重金属离子、盐等各种极端环境下生长,研究酸、重金属离子等极端环境因素对微生物分子、遗传和群落演化的作用特征,揭示嗜酸硫杆菌适应极端环境的分子机制有利于针对特殊环境设计极端环境抗逆菌株,提高菌株应用价值。
第三、嗜酸硫杆菌种群信号交流和调控机制研究。嗜酸硫杆菌应用价值的发挥是微生物整体作用的结果。研究嗜酸硫杆菌种间和种内的信号交流传递和外界环境感应机制,分析不同信号分子的调控功能及其引起的基因表达和细胞行为,开展微生物种群之间调控作用研究,有利于从群体角度指导嗜酸硫杆菌的应用,为种群调控策略提供理论基础。
第四、嗜酸硫杆菌研究技术的开发。相比于大肠杆菌等模式微生物,嗜酸硫杆菌分子操作困难、效率低,制约了嗜酸硫杆菌的理论研究和分子遗传改造,深入探索嗜酸硫杆菌分子遗传操作手段以及蛋白酶学、细胞表型检测技术等,为高效菌株设计改造和开发极端环境微生物底盘细胞奠定基础。