美洲大蠊多肽PAE2 基于AMPK 调控自噬逆转BEL-7402/5-FU 多药耐药的体外研究
2022-06-01李彩琳李银蕊单婉歆吴定宇
欧 玲,李彩琳,李银蕊,周 杰,单婉歆,吴定宇,彭 芳*
(1. 大理大学药学院,云南大理 671000;2.云南省昆虫生物医药研发重点实验室,云南大理 671000;3.大理大学资产与实验室管理处,云南大理 671000)
肝细胞癌(hepatocellular carcinoma,HCC)是全球第五大恶性肿瘤,每年约有70 万人死于HCC〔1-2〕。在我国,HCC 占肝癌85%~90%〔3〕。目前,临床除化疗外,常采用手术治疗、放疗、生物治疗等多种方法联合治疗肝癌,其中化疗是临床治疗肝癌的重要手段,但肿瘤细胞在接受常规的化疗药物治疗后通常会产生耐药,甚至具有多药耐药性(multi-drug resistance,MDR)〔4〕。因此,寻找治疗性生物靶标成为近年来肿瘤治疗的热点。腺苷-磷酸活化蛋白激酶(adenosine monophosphate-activated protein kinase,AMPK)是真核细胞中重要的能量感受器之一,对细胞内的AMP/ATP 比值非常敏感,任何导致该比值升高的因素均可激活AMPK,活化的AMPK 可参与调节自噬过程〔5〕。细胞自噬是一种溶酶体依赖的自我消化过程,多项研究表明,自噬可双向调节MDR,抑制自噬可能增强耐药肿瘤细胞对化疗药物的敏感性〔6-7〕。
中药具有多靶点、多层面的特点,在逆转肿瘤MDR 方面具有独特的优势〔8〕。美洲大蠊Periplaneta americana L.为昆虫纲蜚蠊目蜚蠊科大蠊属昆虫,具有抗炎、抗氧化、抗肿瘤等作用〔9-10〕。课题组前期以美洲大蠊为原料提取分离得到脱脂膏和CⅡ-3,进一步对CⅡ-3 进行分离得到多肽PAE2,研究表明,PAE2具有抗肝癌和逆转肝癌多药耐药作用,其相关机制与影响耐药相关蛋白及酶表达从而减少药物排出、促进细胞凋亡以及抑制血管生成有关〔11-13〕。然而,PAE2调节自噬的机制尚不清楚。因此,本研究采用人肝癌细胞株BEL-7402 及其耐药株BEL-7402/5-FU为试验对象,以索拉非尼为阳性对照药,以AMPK为靶点探讨美洲大蠊多肽PAE2对自噬的影响。
1 材料
1.1 仪器TCS SP8 型激光共聚焦显微镜(德国Leica 公司);TD3 型台式低速离心机(湖南湘仪实验仪器开发有限公司);CKX41SF 倒置相差显微镜(日本Olympus 公司);SN255939 型全自动酶标仪、3111 型CO2培养箱(美国Thermo Fisher Scientific公司);785BR15145 型PCR 仪(美国Bio-Rad)。
1.2 药品与试剂美洲大蠊多肽PAE2(白色冻干粉末,批号:P18267,纯度≥98%)由大理大学药学院张成桂博士提供;索拉非尼(批号:819C021,纯度≥99%)、噻唑蓝(MTT,批号:C0009-2)、青链霉素混合液(100X,批号:20190905)、DAB 浓缩液(批号:20181120)均购自北京索莱宝科技有限公司;甘油(批号:20200414)购自昆明远方生物制品有限公司;五氟尿嘧啶(5-FU,批号:#WXBC0190V)、二甲基亚砜(DMSO,批号:RNBD6084)均购自Sigma 公司;RPMI-1640 培养基(批号:8521447)、胎牛血清(FBS,批号:18050302)、牛血清白蛋白(BSA,批号:15260-037)均购自美国Gibco 公司;DAPI(批号:MAO128-Jul-14F)购自美仑生物;0.25% 胰蛋白酶-EDTA(批号:MSTE1905X)购自迈基生物技术有限公司;AMPK 试剂盒(批号:201890901)购自江苏晶美生物科技有限公司;AICAR(批号:A818441337769)购自爱普拜生物技术有限公司;BeyoIIcDNA 第一链合成试剂盒(批号:081220200922/0761819083)、BeyoFastTMSYBR GREEN qPCR Mix(批号:121919200527)均购自碧云天生物技术有限公司。
1.3 细胞人肝癌细胞株BEL-7402 (批号:MXC050)、人肝癌耐药细胞株BEL-7402/5-FU(批号:MXC049)均购于上海美轩生物科技有限公司。
2 方法
2.1 细胞培养BEL-7402、BEL-7402/5-FU 细胞用含10% FBS 和1%青链霉素的RPMI-1640 培养基在37 ℃、5% CO2的细胞培养箱中培养,另在BEL-7402/5-FU 细胞的培养基中加入20 μg/mL 的5-FU 以维持其耐药性。2 种细胞均要1~2 d 更换培养基,长满培养皿80%以上进行传代,选择对数生长期细胞进行试验。
2.2 药液制备美洲大蠊多肽PAE2用磷酸盐缓冲液(PBS)制成50 mg/mL 的储备液,索拉非尼用DMSO 配制成10 mg/mL 的储备液,用0.22 μm 医用微孔滤膜过滤除菌后分装,4 ℃冰箱保存备用,试验时用0.9%氯化钠溶液稀释至相应浓度后进行给药。
2.3 分组与给药试验分为敏感组、耐药组、索拉非尼组、PAE2低剂量组、PAE2中剂量组、PAE2高剂量组、PAE2低剂量联合AICAR 组、PAE2中剂量联合AICAR 组、PAE2高剂量联合AICAR 组。索拉非尼及美洲大蠊多肽PAE2剂量参照文献〔14〕,AICAR 作用浓度由MTT 试验确定。敏感组加入BEL-7402 细胞,其余各组加入BEL-7402/5-FU 细胞。给药时,敏感组和耐药组加入常规培养基,给药组则加入含相应药物的培养基。
2.4 MTT 比色法检测AICAR 的作用时间及浓度取对数生长期的BEL-7402/5-FU 细胞接种于96 孔板中,于37 ℃、5% CO2培养过夜后,加入含不同浓度的AICAR 培养基,每个浓度设10 个复孔,每孔100 μL,同时设置试剂对照组和细胞空白对照组。培养48 h 后每孔加入20 μL MTT,继续培养4 h,终止培养。每孔加入100 μL DMSO,振荡30 min,待紫色结晶完全溶解后,酶标仪测570 nm 处OD 值(OD570)。计算AICAR 对BEL-7402/5-FU 细胞的生长抑制率,处理数据得20%抑制浓度(IC20),以IC20作为试验剂量与美洲大蠊多肽PAE2不同剂量联合用于进行后续试验。
2.5 酶联免疫吸附试验(enzyme linked immunosorbent assay,ELISA)检测AMPK 的含量检测敏感组,耐药组,索拉非尼组,PAE2低、中、高剂量组AMPK 含量。敏感组取对数生长期的BEL-7402 细胞,其余各组取对数生长期的BEL-7402/5-FU 细胞,接种于96 孔板中,待细胞生长融合至70%左右,除敏感组和耐药组外,其余各组分别加入相应药物。于37 ℃、5% CO2培养箱中常规培养48 h,培养结束后取细胞培养液2 500 r/min 离心10 min,取上清,按照试剂盒说明书进行检测。
2.6 实时聚合酶链式反应(real-time polymerase chain reaction,real-time PCR)检测AMPK mRNA 的表达Trizol 试剂提取总RNA,测定其浓度与纯度,按照cDNA 第一链合成试剂盒说明合成cDNA,以cDNA 为模板采用两步法进行扩增。反应程序:预变性(95 ℃2 min);PCR 反应(40 个循环;95 ℃15 s;60 ℃30 s;72 ℃30 s)。增加溶解曲线步骤:95 ℃15 s,60 ℃30 s。选择GAPDH 为内参,2-△△Ct法计算AMPK mRNA 表达水平。
2.7 real-time PCR 检测细胞自噬相关因子mRNA的表达以GAPDH 为内参,两步法进行扩增,2-△△Ct法计算细胞中自噬相关蛋白5(autophagy related protein 5, ATG5)、自噬相关蛋白7(autophagy related protein 7, ATG7)、磷酸肌醇-3-激酶3(phosphoinositide-3-kinase class 3, PIK3C3)、Beclin 1、微管相关蛋白1 亲链3-Ⅱ(microtubuler-associated protein 1 light chain 3-Ⅱ, LC3-Ⅱ)、P62、Zeste 同源物增强子2(enhancer of Zeste homolog 2, EZH2)的mRNA 相对表达水平。引物序列见表1。

表1 real-time PCR 引物序列
2.8 激光共聚焦显微镜观察LC3-Ⅱ蛋白的表达取对数生长期的BEL-7402 和BEL-7402/5-FU 细胞,接种至预先放有细胞爬片的24 孔板中,常规培养24 h 后加入含相应受试药物的培养基继续培养48 h。每孔加入冰丙酮300 μL,室温固定15 min;5% BSA 封闭60 min。一抗孵育过夜,二抗孵育2 h。DAPI 避光染色10~15 min。抗荧光淬灭封片剂封片,独立重复试验3 次,避光保存。第2 天于激光共聚焦显微镜下拍照观察,每组6 个平行。
2.9 免疫细胞化学法(immunocytochemistry,ICC)检测P62 蛋白的表达取对数生长期的BEL-7402 和BEL-7402/5-FU 细胞,接种至预先放有细胞爬片的12 孔板中,常规培养24 h 后加入含相应受试药物的培养基继续培养48 h。3% H2O2与甲醇按1∶4 的体积比均匀混合后滴入各爬片上,室温孵育30 min;5% BSA 室温封闭10 min;一抗、二抗孵育;DAB 显色试剂盒显色至所需棕色后加入蒸馏水终止反应;苏木素复染30 s,将标本放入饱和Na2HPO4溶液中浸泡2 min,取出后立即用蒸馏水清洗;乙醇梯度脱水,甘油封片,倒置显微镜下观察拍照。使用Image J 6.0 软件计算平均光密度值。
2.10 统计分析采用SPSS 17.0 统计软件进行数据处理,所得数据均采用(±s)表示,多组间比较采用单因素方差分析,两组间比较采用t 检验,检验水准α=0.05,P<0.05 为差异有统计学意义。
3 结果
3.1 AICAR 的作用时间及浓度采用MTT 法对AMPK 激活剂AICAR 的作用时间及浓度进行检测,结果发现,AICAR 对细胞的杀伤呈时间和浓度依赖,后续细胞试验中与美洲大蠊多肽PAE2不同剂量组联合使用的浓度为IC20,作用时间为48 h。经计算,激活剂AICAR 作用浓度为0.006 mmol/L,即1.55 mg/mL(分子量为258.23)。见封三图1。
3.2 美洲大蠊多肽PAE2 对细胞中AMPK 含量的影响与敏感组相比,耐药组细胞中AMPK 含量明显增多(P<0.05),PAE2高剂量组细胞中AMPK 含量明显降低(P<0.05);与耐药组相比,索拉非尼组、PAE2各剂量组细胞中AMPK 含量均显著降低(P<0.05);与索拉非尼组相比,PAE2各剂量组中AMPK含量均降低,但差异无统计学意义(P>0.05)。见表2。
表2 美洲大蠊多肽PAE2 对细胞中AMPK 含量的影响(±s,n=8,pg/mL)

表2 美洲大蠊多肽PAE2 对细胞中AMPK 含量的影响(±s,n=8,pg/mL)
注:与敏感组比较*P <0.05;与耐药组比较#P<0.05。
组别 AMPK 含量敏感组 135.82±9.27耐药组 147.40±5.67*索拉非尼组 132.29±7.55#PAE2 低剂量组 128.41±8.08#PAE2 中剂量组 128.72±11.50#PAE2 高剂量组 126.10±8.12*#
3.3 美洲大蠊多肽PAE2 对细胞中AMPK mRNA表达水平的影响与敏感组相比,耐药组细胞中AMPK mRNA 表达水平显著升高(P<0.05),索拉非尼组细胞中AMPK mRNA 表达水平显著降低(P<0.05);与耐药组相比,索拉非尼组细胞中AMPK mRNA 表达水平显著降低(P<0.05),PAE2各剂量组细胞中AMPK mRNA 表达水平也有不同程度降低,以PAE2高剂量组效果最佳(P<0.05),但效果不及索拉非尼。见表3。
表3 美洲大蠊多肽PAE2 对细胞中AMPK mRNA表达水平的影响(±s,n=4)
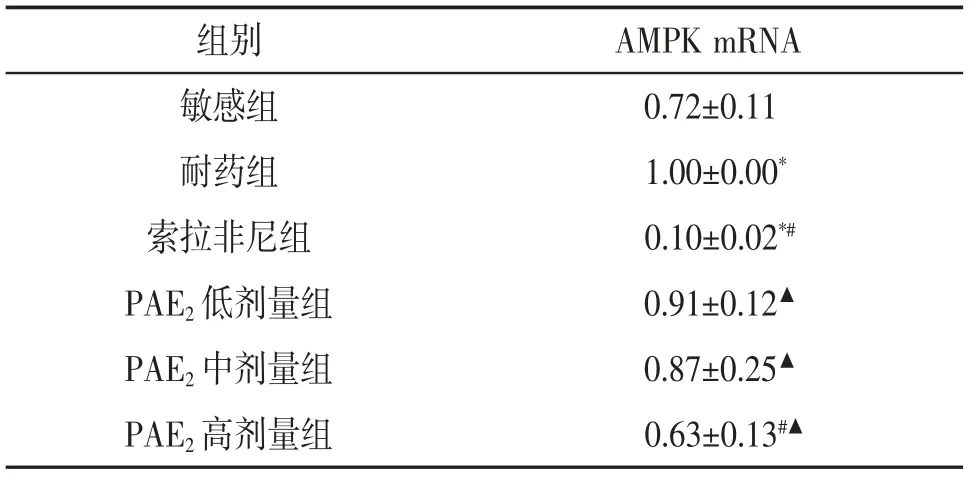
表3 美洲大蠊多肽PAE2 对细胞中AMPK mRNA表达水平的影响(±s,n=4)
注:与敏感组比较*P<0.05;与耐药组比较#P<0.05;与索拉非尼组比较▲P<0.05。
组别 AMPK mRNA敏感组 0.72±0.11耐药组 1.00±0.00*索拉非尼组 0.10±0.02*#PAE2 低剂量组 0.91±0.12▲PAE2 中剂量组 0.87±0.25▲PAE2 高剂量组 0.63±0.13#▲
3.4 美洲大蠊多肽PAE2 对细胞自噬相关因子mRNA 表达水平的影响与敏感组相比,耐药组ATG5 mRNA、ATG7 mRNA 和PIK3C3 mRNA 表达水平均升高,且PIK3C3 mRNA 的表达水平差异有统计学意义(P <0.05)。与耐药组相比,PAE2各剂量组ATG5 mRNA 和PIK3C3 mRNA 表达水平均明显降低(P<0.05),ATG7 mRNA 表达水平也降低,但仅PAE2中剂量组的差异有统计学意义(P<0.05)。PAE2联合AICAR 给药后,ATG5 mRNA、ATG7 mRNA、PIK3C3 mRNA 表达水平见表4。
表4 美洲大蠊多肽PAE2 对细胞自噬相关因子ATG5 mRNA、ATG7 mRNA、PIK3C3 mRNA表达水平的影响(±s,n=8)

表4 美洲大蠊多肽PAE2 对细胞自噬相关因子ATG5 mRNA、ATG7 mRNA、PIK3C3 mRNA表达水平的影响(±s,n=8)
注:与敏感组比较*P<0.05;与耐药组比较#P<0.05;与索拉非尼组比较▲P<0.05。
组别 ATG5 mRNA ATG7 mRNA PIK3C3 mRNA敏感组 0.85±0.11 0.80±0.07 0.49±0.09耐药组 1.00±0.00 1.00±0.00 1.00±0.00*索拉非尼组 0.25±0.05*# 1.12±0.21* 0.29±0.05*#PAE2 低剂量组 0.78±0.22#▲ 0.91±0.26 0.53±0.07#▲PAE2 中剂量组 0.47±0.08*#▲ 0.53±0.06#▲ 0.20±0.07*#PAE2 高剂量组 0.48±0.05*#▲ 0.82±0.07▲ 0.67±0.14*#▲PAE2 低剂量联合AICAR 组 1.04±0.10*▲ 0.70±0.14#▲ 0.14±0.03*#▲PAE2 中剂量联合AICAR 组 1.11±0.19*▲ 1.14±0.08* 0.28±0.05*#PAE2 高剂量联合AICAR 组 0.36±0.09*# 1.26±0.17*# 0.21±0.04*#
与敏感组相比,耐药组Beclin 1 mRNA 和LC3-ⅡmRNA 表达水平明显升高(P<0.05),P62 mRNA和EZH2 mRNA 表达水平明显降低(P<0.05)。与耐药组相比,PAE2各剂量组Beclin 1 mRNA 和LC3-ⅡmRNA 表达水平显著降低(P<0.05),P62 mRNA和EZH2 mRNA 水平显著升高(P<0.05)。与PAE2单用相比,PAE2低、高剂量联合AICAR 组P62 mRNA的表达水平显著降低(P<0.05),PAE2低、中、高剂量联合AICAR 组EZH2 mRNA 的表达水平均显著降低(P<0.05),PAE2低、中剂量联合AICAR 组LC3-II mRNA 的表达水平显著升高(P<0.05)。见表5。
表5 美洲大蠊多肽PAE2 对细胞自噬相关因子Beclin 1 mRNA、LC3-ⅡmRNA、P62 mRNA、EZH2 mRNA表达水平的影响(±s,n=8)

表5 美洲大蠊多肽PAE2 对细胞自噬相关因子Beclin 1 mRNA、LC3-ⅡmRNA、P62 mRNA、EZH2 mRNA表达水平的影响(±s,n=8)
注:与敏感组比较*P<0.05;与耐药组比较#P<0.05;与索拉非尼组比较▲P<0.05;与PAE2 单用比较△P<0.05。
组别 Beclin 1 mRNA EZH2 mRNA敏感组 0.25±0.05 4.53±0.39耐药组 1.00±0.00* 1.00±0.00*索拉非尼组 0.13±0.01*# 0.72±0.38*PAE2 低剂量组 0.03±0.00*#▲ 9.04±0.22*#▲PAE2 中剂量组 0.06±0.01*#▲ 3.55±0.62*#▲LC3-ⅡmRNA 0.59±0.06 1.00±0.00*0.30±0.03*#0.15±0.07*#▲0.29±0.07*#P62 mRNA 1.86±0.25 1.00±0.00*0.14±0.02*#8.09±1.15*#▲1.35±0.15#0.38±0.08*# 8.67±0.70*#▲PAE2 低剂量联合AICAR 组 0.05±0.01*#▲ 0.99±0.05*▲△ 0.52±0.05*△ 0.72±0.10*△PAE2 中剂量联合AICAR 组 0.05±0.01*#▲ 0.55±0.03#▲△ 0.86±0.13*▲ 1.47±0.29*#▲△PAE2 高剂量组 0.16±0.04*# 9.29±0.32*#▲PAE2 高剂量联合AICAR 组 0.07±0.01*#▲ 0.69±0.15*△0.29±0.03*#0.41±0.08*#△
3.5 美洲大蠊多肽PAE2 对LC3-Ⅱ蛋白的影响为了探究美洲大蠊多肽PAE2对自噬相关蛋白LC3-Ⅱ的影响,运用激光共聚焦显微镜观察LC3-Ⅱ蛋白在各组细胞中的表达。两种荧光融合后,与敏感组相比,耐药组细胞自噬作用降低,PAE2各剂量组给药后自噬作用明显降低;PAE2联合AICAR给药后,细胞中绿色荧光明显增多,自噬作用增强。
3.6 美洲大蠊多肽PAE2 对细胞自噬蛋白P62 的影响ICC 结果显示,敏感组细胞大量表达P62,其余各组细胞P62 表达减少,索拉非尼和PAE2均对P62有促进作用,且PAE2作用优于索拉非尼。PAE2联合AICAR 给药后,P62 表达减少。见封三图2。与敏感组相比,耐药组细胞中P62 表达明显降低(P<0.05),提示耐药细胞自噬作用增强;与耐药组相比,索拉非尼组和PAE2各剂量组P62 表达均显著增加(P<0.05),表明药物对细胞自噬起到了抑制作用。PAE2联合AICAR 给药后,P62 表达明显降低(P<0.05),即AICAR 激活AMPK 使细胞自噬作用增强。见表6。
表6 美洲大蠊多肽PAE2 对细胞自噬蛋白P62 的影响(±s,n=8)

表6 美洲大蠊多肽PAE2 对细胞自噬蛋白P62 的影响(±s,n=8)
注:与敏感组比较*P<0.05;与耐药组比较#P<0.05;与索拉非尼组比较▲P<0.05;与PAE2 单用比较△P<0.05。
组别 P62敏感组 0.16±0.01耐药组 0.09±0.01*索拉非尼组 0.11±0.01*#PAE2 低剂量组 0.12±0.00*#PAE2 中剂量组 0.13±0.01*#▲PAE2 高剂量组 0.13±0.01*#▲PAE2 低剂量联合AICAR 组 0.09±0.01*▲△PAE2 中剂量联合AICAR 组 0.10±0.01*▲△PAE2 高剂量联合AICAR 组 0.10±0.01*▲△
4 讨论
肝癌患者病死率较高,且因临床诊断发现晚,手术治疗效果并不理想。目前临床上的治疗手段以化疗为主。索拉非尼是一种口服多靶点激酶抑制剂,具有较好的抑制肿瘤细胞生长、增殖作用,是首个被美国食品药品监督管理局(Food and Drug Administration,FDA)批准的用于治疗晚期HCC 的分子抑制剂,因此本研究选择索拉非尼作为阳性对照药〔15〕。由于在化疗过程中易出现耐药现象,疗效不佳,因此,愈来愈多的研究者对肿瘤耐药机制及如何逆转耐药进行深入探索。目前临床上使用的逆转剂种类较多,但由于其逆转效率不高且机制单一,应用受到限制。而中药具有多靶点、多阶段性作用的优势,可以针对肿瘤MDR 的多种机制进行有效的逆转。课题组前期研究〔11-13〕表明美洲大蠊多肽PAE2具有逆转肝癌MDR 作用,但对自噬的影响未知。AMPK 是将多种细胞功能和生理过程及能量利用联系起来的主要传感器之一,而自噬的主要诱导因素为营养缺乏和(或)能量缺乏,在生物进化过程中,这种内在特性从未改变,所以,AMPK 是自噬的重要上游调节因子〔4〕,且可从多层面促进自噬。因此,本研究基于AMPK 影响自噬来探讨PAE2逆转肿瘤多药耐药作用机制。
肿瘤细胞在营养缺乏、缺氧、饥饿等压力环境下,AMPK 被激活,从而保护细胞免于死亡。ELISA结果显示,BEL-7402/5-FU 细胞株中AMPK 含量明显高于BEL-7402 细胞株,差异具有统计学意义,推测这是耐药细胞的一种保护机制,肿瘤耐药细胞能量代谢异常,需要更多的能量。为了验证这一结果,采用real-time PCR 检测细胞中AMPK mRNA 的表达水平,结果显示出与ELISA 同样的趋势。基于上述结果可知,耐药细胞中AMPK 高表达,美洲大蠊多肽PAE2可抑制AMPK 的表达,且以PAE2高剂量组效果较优。
LC3-Ⅱ定位于自噬体内外膜之间,自噬体与溶酶体结合后,LC3-Ⅱ被溶酶体的水解酶降解,因此可作为自噬标记物。激光共聚焦显微镜观察LC3-Ⅱ的表达,结果显示美洲大蠊多肽PAE2可降低耐药细胞株中LC3-Ⅱ蛋白的表达,即抑制耐药细胞自噬。P62 是一种重要的选择性自噬的调节剂,也是自噬底物。P62 可以和LC3-Ⅱ及自噬体内膜上的泛素化底物结合,形成自噬溶酶体被降解,自噬抑制可导致P62 积累,在HCC 的发生过程中扮演重要角色〔16〕。ICC 结果表明美洲大蠊多肽PAE2可提高耐药细胞株中P62 蛋白的表达。哺乳动物自噬体形成涉及2 个泛素化样修饰,即ATG12 修饰和LC3修饰〔17-18〕。ATG12 被泛素活化酶E1 样酶ATG7 激活后转移到泛素结合酶E2 样酶ATG10,并与ATG5结合形成自噬体前体。Beclin 1 是哺乳动物中最先被确定的特异性自噬调控基因之一,AMPK 可直接在Thr388 处磷酸化Beclin 1,Beclin 1 中Thr388 处的磷酸化对于诱导自噬是必不可少的,同时AMPK 促进Beclin 1 与PIK3C3 的直接结合以激活自噬〔19〕。AMPK 还可以特异性磷酸化EZH2,破坏EZH2 与Zeste 12 同源物1 抑制因子2(suppressor of zeste 12 homolog,SUZ12)的相互作用,导致多梳抑制复合物2(polycomb repressive complex 2,PRC2)靶基因被上调,从而抑制肿瘤细胞的生长〔20〕。real-time PCR 结果显示,美洲大蠊多肽PAE2能够使自噬相关因子ATG5 mRNA、ATG7 mRNA、PIK3C3 mRNA、Beclin 1 mRNA、LC3-ⅡmRNA 的表达水平降低及P62 mRNA 和EZH2 mRNA 的表达水平升高。AICAR 是AMPK 的激活剂,PAE2联合AICAR 用药后,AMPK被激活,细胞自噬有一定程度的增强。上述结果进一步证实了美洲大蠊多肽PAE2通过下调AMPK 的表达而抑制肿瘤细胞自噬。
虽然美洲大蠊多肽PAE2抑制了耐药细胞的自噬,但PAE2各剂量组对细胞自噬的影响并未呈现出剂量依赖性,这可能与PAE2多靶点、多层面、多途径的作用特点、自噬过程及肿瘤多药耐药的复杂性有关,也可能与PAE2未达到有效治疗剂量或者超出有效浓度而出现拮抗作用有关,中药的量效关系仍处于探索阶段。此外,PAE2对P62 及其mRNA的表达均体现出上调的作用,但各剂量组之间的增强作用并不一致,在real-time PCR 结果中甚至出现PAE2低、高剂量组效果较优而中剂量组效果不明显的情况,这可能与药物在体外细胞中的代谢过程及肿瘤微环境的不稳定性,以及P62 在转录水平和翻译水平对细胞自噬的调节作用不同有关,后续可以通过克隆、突变P62,然后进行质粒构建与重组以及细胞转染等操作,以此来探究P62 在蛋白水平和基因水平表达不一致的原因。
综上所述,美洲大蠊多肽PAE2可能通过抑制AMPK 而抑制自噬以达到逆转肝癌多药耐药的作用。在后续研究中,需要进一步探索AMPK 对自噬是起到一般调节作用还是主导作用,PAE2作用为何未呈现剂量依赖性的原因,体外和体内试验是否呈现相同的效果等。后续课题组也将对PAE2对自噬作用的影响进行体内研究,以进一步完善PAE2逆转肝癌MDR 的机制,为PAE2的临床研究奠定基础。
