水稻铁吸收与转运机理
2022-05-27邱冠凯倪大虎韩忠民王婧莹方军
邱冠凯,倪大虎,韩忠民,王婧莹,方军
(1.中国科学院 东北地理与农业生态研究所,黑龙江 哈尔滨 150081; 2.中国科学院大学,北京 100049;3.安徽省农业科学院,安徽 合肥 230031)
0 引 言
铁作为植物生长发育过程中最重要的微量元素之一,其参与了植物的多种细胞活动。铁参与多种氧化还原反应,尤其是在光合作用和呼吸作用的电子传递链中起到关键性的作用。铁在土壤中的溶解度很低,溶解的铁离子主要以三价的形式存在,并随着pH值的升高溶解度显著降低。因此,植物体对土壤中对于铁离子的利用率很低[1],尤其在碱性土壤中容易产生缺铁胁迫,缺铁会直接对植物体的生长发育、农作物的产量与质量造成影响[2]。在铁元素严重缺乏或者高pH土壤地区种植水稻,就会面临缺铁胁迫。因此,研究水稻在碱性条件下对铁元素吸收和转运分子机制,可以为改善水稻在碱性土壤中的缺铁胁迫耐受性提供理论基础[3]。
1 植物的铁吸收机制
植物体对于铁元素的吸收主要通过两种不同的途径[1]。双子叶植物以及非禾本科植物均采用机理Ⅰ(strategyⅠ),该途径吸收的铁离子为二价铁离子。而对多数禾本科植物而言,主要通过机理Ⅱ(strategyⅡ)吸收三价铁离子。通过机理Ⅰ吸收铁的植物体,分泌次生代谢产物尼克酰胺(Nicotianamine,NA),这种次生代谢产物可以直接与土壤中的二价铁离子结合,植物体通过自身的一些转运蛋白将其运输到根部细胞内[4]。通过机理Ⅱ吸收铁的植物体,其根部细胞分泌的次生代谢产物麦根酸(Mugineic acids,MAs)直接结合土壤中的三价铁离子,再由植物体转运到细胞内[4]。水稻吸收铁元素同时并存这两种吸收方式[5]。
1.1 铁吸收机理Ⅰ
某些植物包括部分非禾本科植物和双子叶植物吸收铁离子时采用机理Ⅰ。这些植物的根部在吸收铁离子的过程中具有极强的还原能力,可将一些重要的还原剂排出体外以还原土壤中的三价铁离子[3],增强根部对于铁离子的吸收效果(图1)。采用机理Ⅰ的植物在缺铁环境下,通过向土壤中释放正电荷酸化土壤,增加土壤中二价铁离子的浓度,再由铁离子转运蛋白将其转运至细胞内[6]。

图1 高等植物铁吸收机制
转运蛋白IRT(The iron-regulated transporter)能够将二价铁离子从根部附近的土壤中转运至植物体细胞内[7]。有研究发现水稻中存在两个IRT基因(OsIRT1和OsIRT2),它们编码ZIP(Zinc-regulated transporter,IRT-like protein)转运蛋白家族。拟南芥中相应缺铁信号基因AtIRT1,不仅在植株的铁转运过程中起到重要的作用,还与植物体的光合作用有关。irt1敲除突变体呈现出叶片黄化的表型,生长发育受到了严重的影响[8]。研究发现,水稻体内有多种Fe2+转运蛋白,包括OsIRT1、OsIRT2、OsNRAMP1和OsNRAMP5[4,9-10]。OsIRT1就是二价铁离子的重要转运蛋白[4],在缺铁条件下,三种基因OsIRT1、OsIRT2和OsNRAMP1表达量升高,而OsNRAMP5表达量基本不变,对铁吸收的调控作用相对较小[10-11]。
1.2 铁吸收机理Ⅱ
作为禾本科植物主要的铁吸收方式,机理Ⅱ与机理Ⅰ大不相同,采用机理Ⅱ的植物体能够合成MAs并释放到根际土壤中[12]。MAs首先结合土壤周围的三价铁离子,形成Fe(Ⅲ)-MA,再由 YS1(Yellow stripe 1)转运蛋白运送至细胞内[13]。
S-腺苷甲硫氨酸(S-adenosylmethionine,SAM)是MAs合成过程中的重要前体(图2)。MAs的合成通过以下三种酶完成:尼克酰胺合成酶NAS、尼克酰胺氨基转移酶NAAT和脱氧麦根酸合成酶DMAS[14-16]。由NAS将三分子的SAM结合成一个分子的NA,NA可以被NAAT、DMAS还原酶作用合成2-DMA。囊泡中所合成的MAs主要在根部细胞中发现,也正是因为这些囊泡在昼夜间收缩,所以MAs可能是在囊泡中合成并进行储存。水稻中虽然存在类似的囊泡活动,但相比于大麦合成的MAs要减少许多[17]。DMA转运蛋白在缺铁情况下可被诱导表达,通过诱导表达OsTOM1可增加水稻对缺铁的耐受性[18],OsTOM1在水稻叶根中都有所表达,它还参与铁离子的转运过程。OsTOM2是OsTOM1的同源基因,在细胞质中表达并参与转运水稻中的DMA[19]。DMA可以直接与Fe3+结合形成Fe(Ⅲ)-DMA复合物,转运蛋白YS/YSL(Yellow stripe-like),将其直接运输至植物体根部细胞内[13]。该转运蛋白首先是在玉米和大麦中发现,之后在水稻中克隆出18个相关的同源基因[20]。OsYSL15编码三价铁离子与DMA螯合物转运蛋白[21]。OsYSL16虽然也编码转运蛋白,它主要是在根表皮中表达[22-23]。水稻在缺铁条件下,OsNAS1和OsNAS2的表达量升高。缺铁条件下OsNAS3的表达量却很低,但在过量铁的环境下却能够强烈表达OsNAS3。OsNAS3主要在根中表达,在维管束、表皮及外表皮细胞中也有所表达[24]。osnas3敲除突变体在铁过量条件下表现出生长不良、干重降低、叶片黄化严重及铁在叶片中的积累量增多等现象[25]。在铁过剩条件下,OsNAS3能够合成NA以缓解铁的过剩所带来的不利影响。而在正常铁含量条件下,OsNAS1和OsNAS2合成NA可增强铁的转运[26]。

图2 禾本科植物中甲硫氨酸循环及麦根酸类物质的合成过程[28]
在正常生理环境下,水稻通过机理I方式吸收铁元素,并在吸收过程中发生强烈的氧化还原反应。而在缺铁环境条件下会通过机理Ⅱ吸收铁元素,吸收进入植物体内的铁元素从始至终都以螯合物的形式存在,并且不会发生氧化还原反应[27]。
2 水稻铁离子吸收和转运的分子机制
2.1 IDEF和HRZ调控铁离子的吸收和转运
转录因子OsIDEF1作为水稻在缺铁环境下表达的重要基因[29-31],能够识别下游顺式作用元件IDE1(Iron deficiency responsive cis-actingelements 1),该顺式作用元件的核心序列为5′-CATGC-3′。OsIDEF1可以与多种二价金属离子进行结合,如二价铁离子、锰离子等[32],并能够在缺铁的条件下发挥信号传导的作用[33]。OsIDEF1作为最上游的转录因子,能够正向调控多种铁运输相关载体基因的表达,如OsYSL15、OsYSL2和OsIRT1等。过表达OsIDEF1能够增加水稻的缺铁耐受性并增加其耐盐碱性。参与多种泛素蛋白酶降解的CSN6(COP9 signalosome 6)在铁缺乏早期含量下降,以确保OsIDEF1的快速积累,从而上调铁吸收相关基因,帮助水稻克服缺铁协迫[34]。OsIDEF1能够诱导OsIBP1.1和OsIBP1.2的表达,而OsIBP1.1再调控OsYSL2的表达影响水稻对于铁离子的吸收[35]。转录因子OsIDEF2能够识别顺式作用元件IDE2,该顺式作用元件的核心序列为5′-CA(A/C)G(T/C)(T/C/A)(T/C/A)-3′。研究发现,OsIDEF2在维持水稻植物体铁平衡中起到至关重要的作用[30]。
HRZ(Hemerythrin motif-containing really interesting new gene and zinc-finger protein)作为一种含有血红蛋白结构域的锌指蛋白,能够负向调控植物体对于铁离子的吸收[36]。HRZ可直接与金属离子铁和锌发生结合。含有HRZ的RNAi植株对缺铁具有较强的耐受性,并能够在植株和种子中积累更多的铁离子[37]。OsHRZ1和OsHRZ2同样作为泛素连接酶,能够直接作用于铁信号传导的相关蛋白。OsHRZ1能够直接泛素化OsPRI1、OsPRI2和OsPRI3[38-39]。OsPRI(Psitive regulator of iron homeostasis)是一类bHLH转录因子,水稻中的OsPRI1、OsPRI2和OsPRI3与拟南芥中的bHLH105和bHLH115为同源基因。ospri1基因敲除突变体对铁缺乏敏感性增加,而ospri2和ospri3基因敲除突变体中的铁无法从根部转运至地上部,其过表达植株能够促进植物体铁的积累并上调缺铁应答基因。OsPRI1、OsPRI2和OsPRI3均能够结合OsIRO2和OsIRO3的启动子。其中,OsPRI2和OsPRI3通过结合靶基因启动子还能够激活OsYSL2的表达[39]。因此,OsHRZ1能够通过泛素化OsPRI1、OsPRI2和OsPRI3进而调控OsIRO2和OsIRO3转录,依次生成级联反应来参与植物对于铁离子的吸收。
2.2 bHLH转录因子调控铁离子的吸收和转运
目前,在水稻中发现与铁离子吸收相关的bHLH(basic helix-loop-helix)转录因子,包括OsIRO2[40-42]、OsIRO3[43]和OsbHLH133[44]等。其中,OsIRO2在缺铁条件下被诱导表达并且促进铁离子的吸收,而OsIRO3和OsbHLH133的表达却是抑制铁离子的吸收。顺式作用元件IDE的核心序列为5′-CACGTGG-3′,在缺铁条件下被上游转录因子OsIRO2所识别。缺铁条件下,该基因在根部和地上部的表达量升高[42]。对直接参与铁吸收机理Ⅱ的相关基因的启动子区域中均发现了顺式作用元件IDE,这也说明OsIRO2能够参与水稻中MAs的合成,并调控下游基因的表达[42]。在缺铁条件下,过表达OsIRO3使得植物体地上部铁离子的积累量变少,而与铁离子吸收直接相关的基因表达量却发生下调,由此可见,OsIRO3是水稻对于铁离子吸收和转运的负调控因子[43]。OsbHLH156在铁吸收机理II中起到关键性作用,该基因主要在根中表达,在缺铁条件下该基因功能降低,导致水稻呈现黄化表型。OsbHLH156能够与转录因子OsIRO2协同作用来增强水稻对于缺铁的耐受力[45-46]。通过对缺铁响应基因的转录变化与DNA甲基化变化关联分析,研究人员进一步发现,在缺铁条件下,调控铁响应基因的铁信号通路核心转录因子OsbHLH156和OsIRO2甲基化发生了显著的变化[47]。
缺铁时,植物通过bHLHIVc家族成员(bHLH34,bHLH104,bHLH105,bHLH115)激活下游转录因子FIT和bHLHIb(bHLH38,bHLH39,bHLH100,bHLH101)的表达,后者形成蛋白复合物促进铁吸收基因如IRT1和FRO2的表达,从而促进铁吸收。铁充足或过量时,植物能抑制FIT和bHLHIb转录因子的表达,从而阻止铁过量摄入以避免遭受铁毒害。缺铁条件下,bHLH IVc蛋白激活下游转录因子bHLH156和bHLHIb,进而促进铁吸收基因如IRT1和FRO2的表达以增加铁吸收。随着植物体内铁的增多,bHLH11蛋白也不断积累,并在bHLH IVc蛋白的帮助下进入细胞核,通过招募转录抑制因子[48],从而减少植物体对于铁的吸收。
在植物体中,研究者发现一种特异表达的小肽IRON MAN(IMA),其可以激活缺铁响应基因(bHLHIb、IRT1和FRO2)的表达[49]。在拟南芥中,通过筛选与IMA的互作蛋白,发现其可以与BST蛋白相互作用。IMA的C端含有一个保守的BID(BTS interaction domain)结构域,其负责与BTS蛋白互作。作为一个E3连接酶,BTS可以通过泛素化途径降解IMA。在铁充足情况下,BTS促进bHLH105/bHLH115降解,阻止了铁吸收基因的表达,减少了铁吸收,从而避免植物遭受铁毒害;在铁匮乏情况下,IMA被诱导后与BTS互作,从而阻止了bHLH105/bHLH115的降解,而被释放的bHLH105/bHLH115则激活缺铁响应基因,促进铁吸收,以满足植物的铁需求[50]。OsIMA1和OsIMA2在缺铁条件下受转录因子OsIDEF1、OsbHLH058、OsbHLH059以及两个基因自身的正向调控,而受HRZ泛素连接酶的负向调控。过表达OsIMA1和OsIMA2可以使水稻在正常条件下呈现缺铁表型,而下调两种基因只会产生轻微的影响[51]。
2.3 IRT和NRAMP参与游离态二价铁离子的转运
水稻除了通过两种不同的途径来吸收铁元素外,还能够直接利用铁转运蛋白将Fe2+直接转运至细胞内,这样的铁转运蛋白包括OsIRT1[52-53]、OsIRT2[3]、OsNRAMP1[54]和OsNRAMP5[10-11]。其中对于Fe2+的转运起到关键性作用的为OsIRT1、OsIRT2,缺铁环境可使两种基因表达量升高[3]。在水稻中,过表达OsIRT1使其对缺铁胁迫的耐受性增加。OsIRT2是OsIRT1的同源基因,具有73%的相似度,也在水稻的根部表达。NRAMP家族成员同样也作为水稻吸收铁元素不可或缺的一部分,OsNRAMP1和OsNRAMP5作为7个家族成员中最重要的两个参与其Fe2+的转运,其中的OsNRAMP1同样也可以参与Cd2+的转运[54]。在缺铁环境条件下OsNRAMP1的表达量显著升高,而OsNRAMP5的表达量没有明显的变化[55]。OsNRAMP5的主要功能是转运Cd2+、Mn2+两种金属元素[11,56]。OsVIT2是一个液泡膜定位的二价铁转运蛋白,在叶鞘的束内输导组织和籽粒糊粉层中表达,作用是负责将铁隔离到这些组织的液泡中。敲除OsVIT2后产量不受影响,但会增加铁从节到种子以及从糊粉层到胚乳的分配,从而提高了精米中铁的累积[57]。木薯中过量表达拟南芥液泡铁转运蛋白OsVIT1,在温室条件下可以使木薯块根中铁增加3~4倍[58]。田间试验中,在木薯中共表达突变的拟南芥铁转运蛋白IRT1和蛋白FER1,其铁含量比非转基因对照的铁水平高7~18倍,锌水平高3~10倍,而生长参数和块根产量不受转基因强化的影响[59]。
2.4 YSL家族在铁离子转运中的作用
目前,水稻中已经发现18个YSL家族成员能够参与铁离子的转运[60-62](图3)。二价铁离子的螯合物就是通过OsYSL2转运蛋白将其转运至细胞内部。转运蛋白OsYSL2除了能够转运铁元素外,还能够转运锰元素。研究发现OsYSL2除了在根部表达,外还在植物体其他部位表达,该基因除了参与植物体铁元素的吸收外,还同样参与铁元素在植物体内的运输[62]。OsYSL15[61]、OsYSL16[23]通过机理II吸收三价铁离子和麦根酸螯合物时起到关键性作用。在缺铁的环境条件下,OsYSL15表达量显著升高。osysl15基因敲除突变体呈现发黄枯萎的表型,并且在其植物体中各个部位铁含量均显著降低[62]。OsYSL15在大麦中的同源基因HvYSL1专一性运输Fe3+,在水稻中过表达HvYSL1可增加其耐受性[32]。
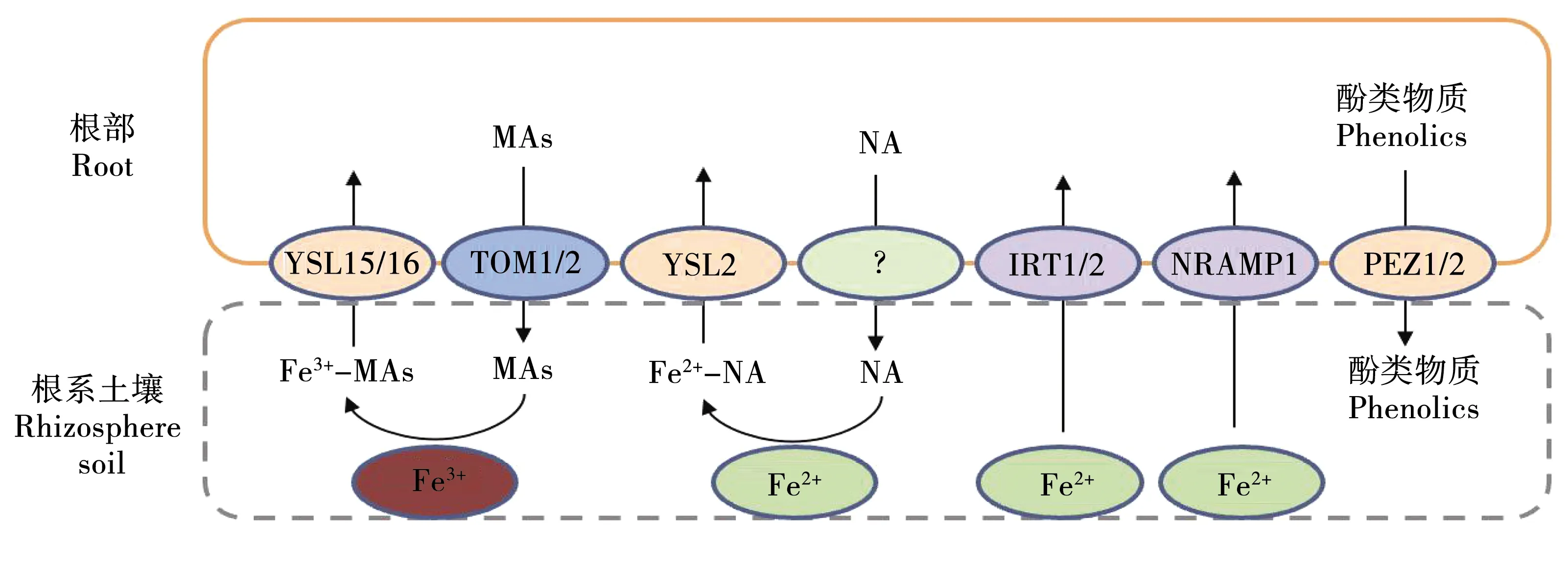
图3 铁离子从根际土壤进入水稻根部细胞过程
3 展望
当前关于水稻吸收铁离子的分子机制的一部分已被阐明(图4),但仍然能有许多领域需要研究。水稻中最上游的两个基因OsIDEF1和OsHRZ1被认为最有可能是铁的感受器,但到目前为止却没有直接证据证明,而且目前仍然没有将水稻中铁的吸收和转运分子机制运用到实际生产中。铁离子在细胞器中的运输方式相关研究很少,如铁离子在叶绿体和线粒体的电子传递中是如何发挥作用的。解析铁离子在信号传导过程中的分子机制,将更有利于对其进行应用。此外,影响植物体铁元素吸收的因素并不局限于这些调控机制。例如,植物体分泌的香豆素可以直接影响到植物根际的微生物群落结构[63-64],从而活化土壤铁营养,促进植物铁吸收等[65]。所以,亟需更深入且从不同角度探究水稻的铁吸收机制,以应用在遗传育种与品种改良中。

图4 缺铁响应信号通路
