非小细胞肺癌预后标志物和预测相关性microRNA的研究
2022-05-27孙珊珊毕莎杉
孙珊珊,冯 虎,毕莎杉
(1.山东大学附属威海市立医院肿瘤 综合治疗科,山东 威海264200;2.山东大学附属威海市立医院 病理科,山东 威海264200)
2019年肺癌(LC)死亡比例占癌症相关死亡的23.5%[1],而非小细胞肺癌(NSCLC)占LC发病率和死亡率的80%以上[2]。随着高通量测序和芯片技术的快速发展,发现了许多与肿瘤预后有关的致病基因,推动了分子革命时代的浪潮。靶向及免疫药物的发现也为NSCLC治疗做出重大贡献,提高了NSCLC患者的无进展生存期(progression free survival,PFS),但其总生存(OS)仍不理想,尤其是晚期患者[3-4]。因此迫切需要发现针对NSCLC的更有效的治疗靶点。
本研究利用GEO(Gene Expression Omnibus)平台上的5个mRNA表达数据集分析了NSCLC与正常组织的差异表达基因(DEGs)。利用Cytoscape软件中的ClueGO 插件进行通路富集分析并利用基于String 11.0数据库的蛋白质-蛋白质相互作用网络(PPI)进行分析。并对差异表达基因进行了靶向miRNA的预测,构建了mRNA-miRNA调控网络。最终筛选出6个与NSCLC预后相关的分子。本研究旨在寻找与NSCLC发生发展和预后相关的潜在靶分子,为临床治疗提供依据和方向。
1 资料和方法
1.1 数据集筛选
从GEO平台(http://www.ncbi.nlm.nih.gov/geo)上获取5个mRNA数据集(GSE18842,GSE19188,GSE21933,GSE33356和GSE102287)和miRNA数据集(GSE53882)。RNA数据集分别基于GPL570(Affymetrix Human Genome U113 Plus 2.0 Array)(GSE18842,GSE19188,GSE33356 和GSE102287);GPL6254(Phalanx Human OneArray)(GSE21933)和GPL18130(State Key Laboratory Human microRNA array 1888)测序。各个数据集包含的NSCLC样本数及正常肺组织样本数分别为:GSE18842为46和45;GSE19188为94和62;GSE33356为60和60;GSE102287为32和34;GSE21933为21和21;GSE53882为397和103。
1.2 数据标准化与差异基因筛选
通过均值、log2转换和插值缺失的表达数据对数据进行标准化处理。利用基于R 3.5.2的limma软件包对NSCLC和正常肺组织的DEGs和差异miRNA(DEMs)进行鉴定,LogFC>2 和校正P值<0.05被认为具有统计学意义。
1.3 差异表达基因的PPI网络与通路富集分析
利用String11.0数据库(https://string-db.org)获取DEGs的PPI信息,富集阈值P值<0.001,利用Cytoscape软件(v.3.7.1)对PPI网络进行可视化。我们在Cytoscape软件中使用了ClueGO插件,进一步分析研究DEGs的生物学过程、细胞组成、分子功能以及KEGG通路信息,P值<0.05被认为具有统计学意义。
1.4 mRNA靶miRNAs的鉴定及mRNA-miRNA相互作用网络
利用ENCORI(Encyclopedia of RNA Interactomes,http://starbase.sysu.edu.cn)进行了差异表达基因靶miRNAs的预测分析,然后筛选预测miRNA和DEMs的重叠miRNA。并在GENEMANIA(http://genemania.org)平台上获取DEGs可能的mRNA相互作用信息。构建mRNA-miRNA调控网络,揭示miRNA对mRNA的调控参与NSCLC的发生发展。
1.5 差异表达基因的临床相关性分析
我们使用公共开放数据库GEPIA(Gene Expression Profile Interactive Analysis,http://gepia.cancer-pku.cn/)来评估DEGs的表达与不同病理类型之间的相关性,分别基于肺腺癌(LUAD)和肺鳞癌(LUSC)资料分析DEGs与TNM分期、PFS和OS的相关性。
2 结果
2.1 DEGs和DEMs的筛选
数据预处理后,通过限制logFC>2和P值<0.05,我们分别得出在NSCLC组织中DEGs有466、309、715、111和650个。DEMs有184个,其中48个miRNA表达下调,49个miRNA表达上调,在5个mRNA数据集中,共筛选出51个重叠基因(上调13个,下调38个),见表1,图1。

表1 51个差异表达基因的情况

图1 5个mRNA数据集差异基因交集的韦恩图(A),以及差异基因表达谱的热图(B、C)。
2.2 DEGs的通路富集分析
利用ClueGo软件进行了GO和KEGG富集分析,发现DEGs与调节细胞分化,组织重塑和蛋白激酶A的生物功能调节密切相关,且与构成多囊泡小体相关。KEGG富集分析表明,DEGs主要聚集在PPAR信号通路中(P值均<0.05)(见图2)。

图2 差异表达基因的GO和KEGG富集分析结果图
2.3 DEGs相互作用与蛋白质-蛋白质调控网络分析
利用GENMANIA网站获取和生成DEGs及它们相互作用基因之间的网络,见图3A。基于STRING平台获取DEGs编码的蛋白质信息,并使用Cytoscape软件生成PPI网络,见图3B,发现 IL6位于调控网络的中心位置,提示DEGs可能通过IL6调节细胞功能。
2.4 DEGs与临床特征的相关性研究
基于TCGA中肺腺癌(LUAD)和肺鳞癌(LUSC)数据集的分析也证实了ANKDR29、IGSF10和PPBP在NSCLC样本中表达下调,相反,THBS2、 GREM1和SPP1在肿瘤组织中表达上调,见图4A。此外,IGSF10与病理分期相关(P=0.039),但其他基因没有发现明显的差异,见图4B。在GEPIA平台中评估分析,我们发现ANKRD29、GREM1、IGSF10、THBS2、PPBP和SPP1对NSCLC患者的OS有显著影响(P值分别为0.012、0.013、0.008、0.042、0.017和0.01),见图5A,但只有IGSF10高表达患者的PFS显著优于低表达患者P=0.019,见图5B。

图4 DEGs在LUAD和LUSC中的表达情况(A)以及在对不同分期的影响(B)

图5 DEGs在NSCLC中OS(A)和PFS(B)中的生存曲线
2.5 mRNA-miRNA互作网络的构建
在51个DEGs中,我们在ENCORI平台上发现了3761个可能相关的miRNAs。与我们获得的DEMs比较后,认为有22个miRNAs参与调控DEGs,见表2。值得注意的是miR-143-3p与IGSF10和GERM1密切相关,结合肺癌组织中miR-143-3p表达上调与IGSF10表达下调趋势,提示miR-143-3p有可能抑制IGSF10转录,影响肺癌细胞分化增殖。
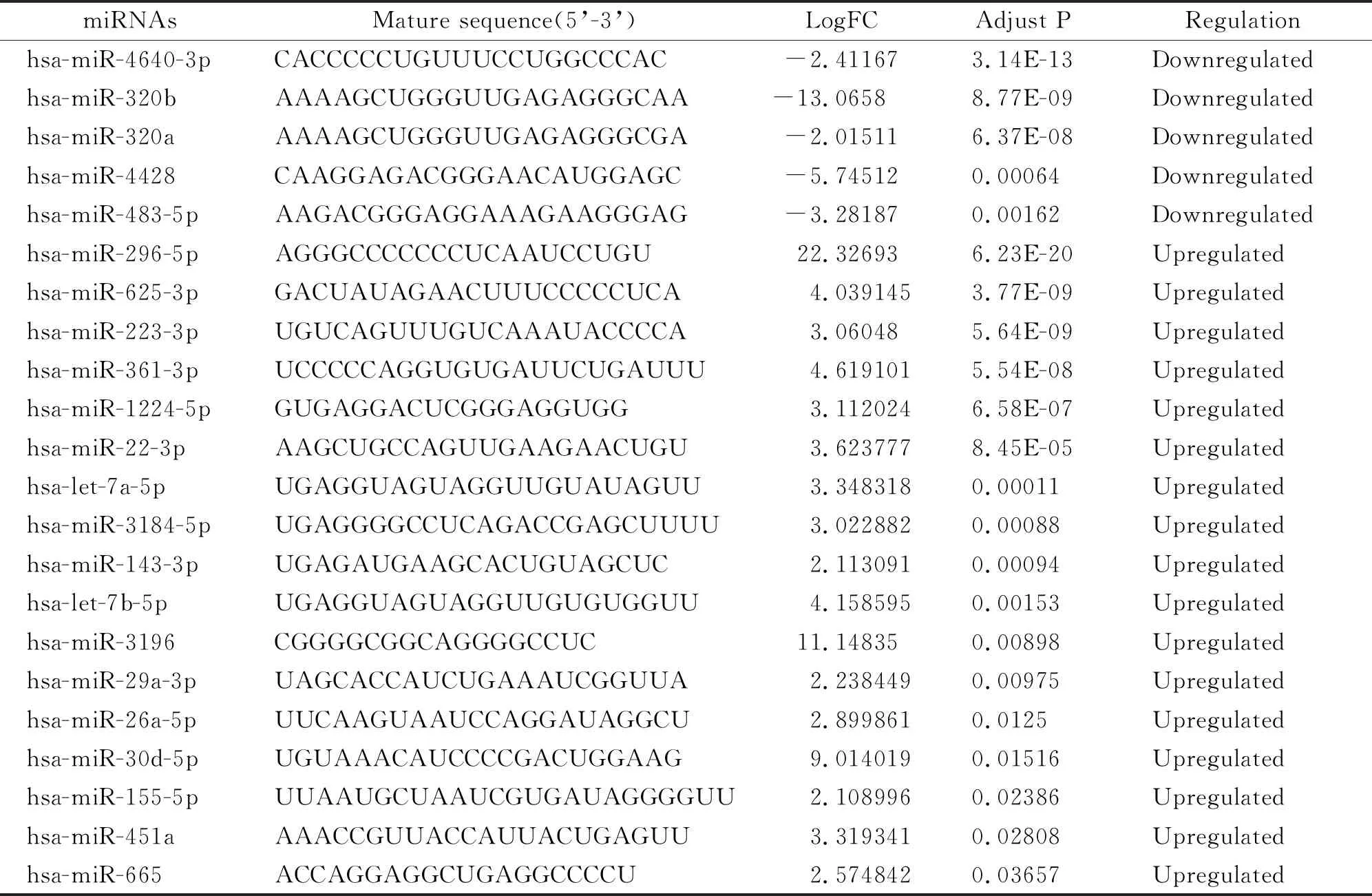
表2 NSCLC中22个重叠差异表达的miRNA的信息
3 讨论
随着信息数字化的发展,包括高通量测序和DNA芯片技术在内的生物信息学分析技术在医学研究领域得到了广泛的应用,挖掘新的生物标志物可能成为诊断和治疗的靶标[5-6]。本研究从GEO平台筛选出5个基因芯片数据集(GSE18842、GSE19188、GSE21933、GSE33356和GSE102287),将每个数据集中的DEGs交互,得到51个与NSCLC的发生发展密切相关的DEGs,应用ClueGo软件对51个DEGs进行GO和KEGG富集分析,我们发现DEGs主要富集于组织重塑、细胞分化和蛋白激酶A(PKA)的调控,并主要构成细胞成分的多囊泡小体。KEGG富集分析证实了PPAR信号通路与NSCLC相关。以往研究表明,PKA通过不同亚基的作用参与细胞分化、肿瘤转化和预后[7-8],靶向PAK可抑制耐selumentinb(一种MEK抑制剂)肺癌细胞的生长[9]。PPAR通路失调会激活肿瘤免疫和缺氧,最终导致代谢紊乱[10]。上述发现间接说明我们研究结果的准确性。此外我们发现有22个miRNAs与DEGs之间存在mRNA-miRNA相互作用网络。有6个候选基因(ANKRD29、GREM1、IGSF10、THBS2、PPBP和SPP1)对NSCLC患者的OS有显著影响。
miRNA通过3’端非编码区与特定mRNA结合来抑制相关基因的蛋白表达,但这一过程不是绝对的[11]。我们的结果显示只有3个基因(IGSF10、GREM1、THBS2)与22个miRNAs相关,miR-143-3p可能在调控IGSF10和GREM1表达方面起关键作用。先前报道表明miR-143-3p在NSCLC中下调导致自噬潮增加,降低对药物的敏感性[12]。IGSF10是定位于细胞膜上的免疫球蛋白超家族成员,在多种肿瘤中检测到突变,而沉默IGSF10可诱导整合素-β1/FAK信号的激活促进肿瘤增殖和黏附[13]。GREM1是编码骨形态发生蛋白拮抗剂家族成员,在细胞外基质合成、胶原合成和生物结构修饰等方面中起关键作用[14]。文献报道,GREM1高表达的患者在乳腺癌、结肠癌、基底细胞癌等疾病中预后较差,这可能与不同的内部机制激活有关,包括激活TGF-β/smad通路、BMP通路或EMT通路[15-16]。这与我们的研究结果相一致。但这些候选基因在NSCLC背景下的认识还不够深入,考虑到分子调控的复杂性和多样性,我们不能给出一个潜在的特定调控环路,我们提出它是否也可以调节IGSF10或GREM1参与NSCLC的发生发展,还需要更多的研究来证实。
前期也有一些生物信息学分析来揭示NSCLC的分子机制[17-18]。与已有研究相比,我们的研究具有更大的数据量(5个微阵列数据集)和更严格的DEGs筛选条件(logFC>2),从而保证了结果的准确性。我们利用预测平台和GEO数据库的结合,建立了mRNA-miRNA相互作用网络,更好地探索候选基因在NSCLC中的意义。但仍有许多不足之处,还需要蛋白和RNA水平上的分子实验来探索这些候选基因在NSCLC中的生物学机制。
综上所述,我们通过综合生物信息学分析确定了6个与NSCLC患者OS相关的DEGs,其中IGSF10与PFS相关。我们发现ANKRD29 和 IGSF10可能与NSCLC发生发展有关,这在以往是鲜有报道的。综合预测和mRNA-miRNA互作网络给出的22个miRNAs可以为下一步的研究指明方向。总之,本研究为NSCLC个体化治疗提供了潜在靶点。
