光照对小麦的春化效应及其机制初探
2022-05-23张洪伟焦义然陈文烨董福双杜进民王海波
张洪伟,焦义然,陈文烨,董福双,杜进民,王海波,周 硕
(1.河北农业大学农学院,河北保定 071001;2.河北省农林科学院生物技术与食品科学研究所/河北省植物转基因中心,河北石家庄 050051;3.河北科技大学食品与生物学院,河北石家庄 050051)
禾谷类作物的抽穗期是其发育完全的幼穗从旗叶中抽出的时间,是决定其产量的重要农艺性状之一。在六倍体小麦中,该性状主要受到三类因素的影响,即春化需求、光周期敏感性以及自身早熟性。春化需求是小麦需要经历长时间的低温才能开花的性质;光周期敏感性是小麦只有在合适的光周期条件下才能开花的性质;满足春化需求和光周期条件后,不同小麦品种间的抽穗期仍有差异,这些差异由自身早熟性(earliness,Eps)决定。
在小麦中,关键的春化调控基因包括、以及,其中和是开花促进基因,是开花抑制基因。小麦基因主要在顶端分生组织和叶片中表达,随春化时间的持续而逐渐上调,是拟南芥定的同源基因;VRN1蛋白可以直接结合基因的启动子,在长日照条件下促进基因在叶片维管束细胞中的大量表达。作为拟南芥Flowering Locus T(FT)的同源蛋白,小麦VRN3被运输到顶端分生组织(shoot apical meristem,SAM),同Flowering Locus D-like 2(FDL2)和14-3-3蛋白互作,进一步促进表达。小麦VRN1在促进表达的同时,还可以结合的启动子,直接抑制其表达,从而解除该基因对小麦开花的抑制作用。小麦及均可被长日照诱导,而且,VRN2可与CONSTANS 2(CO2)竞争结合Heme Activator Protein 3(HAP3)蛋白,形成VRN2/HAP3/HAP5三元复合物,解除CO2对表达的促进作用,只有当被春化所抑制时,CO2才可以结合启动子而促进开花。
除、及基因外,还有其他基因和小麦春化有关。小麦的是拟南芥关键春化基因()的同源基因,是春化相关的开花抑制基因,可被抑制。拟南芥中Vernalization-Insensitive 3(VIN3)所介导的染色质甲基化状态改变是导致拟南芥FLC表达变化的主要原因之一,在小麦中发现,的同源基因()、和的表达均被春化上调。另外,小麦Vernalization-related 2(VER2)可与Glycine-rich RNA Binding Protein 2(GRP2)结合,解除GRP2对小麦表达的抑制,促进开花。
植物可以通过预期周期性的(约24 h)环境变化协调自身生物进程,以更适应外界环境,这个过程依赖于植物精确的昼夜节律(Circadian rhythm),这种节律主要由依次表达的5个Pseudo-Response Regulator(PRR)基因及部分冗余基因Circadian Clock-Associated 1(CCA1)和Late Elongated Hypocotyl(LHY)共同维持,这些基因与GIGANTEA(GI)、LUX ARRHYTHMO(LUX)、Early Flowering 3(ELF3)、ELF4等基因相互作用,构成了极其复杂的基因表达反馈网络,进而调节()表达。光照可以使CO蛋白更稳定,在黑暗条件下CO蛋白被泛素蛋白酶体所降解,因此只有在白天表达才能促进和()表达,并导致提前开花。在小麦中,基因沉默导致小麦不能开花;表达受光周期和生理节律调控,其在拟南芥中的过表达导致拟南芥提早开花,且在缺失突变体(与野生型拟南芥相比明显延迟开花)中恢复了其野生表型;小麦()在拟南芥中的过表达导致其在短日照条件下延迟开花,而沉默导致小麦在长日照和短日照条件下均提早开花;和拟南芥中基因是开花过程中关键光周期响应基因不同,小麦(,拟南芥的同源基因)决定了小麦开花过程中的光周期响应;长日照条件下双突变体(为野生型或显性突变体)比野生型提前3 d开花,说明TaPPD1蛋白存在时,CO1和CO2甚至轻微抑制开花。
为进一步了解温度和光照对小麦开花的影响机制,分别在光照及黑暗条件下对小麦进行春化,分析两种条件下部分开花相关基因的表达情况,旨在明确光照对小麦的春化效应及分子机制。
1 材料与方法
1.1 试验材料及其处理
以小麦中麦175为试验材料,选取大小一致、籽粒饱满的种子,用75%酒精处理1 min,2%次氯酸钠处理10 min,用无菌水冲洗3次;于 22 ℃、黑暗条件下催芽24 h,将催芽的种子播种于营养土和蛭石1∶1混合的基质中,于22 ℃、光照条件下(16 h光照/8 h黑暗光周期,光照强度约4 000 lx)培养7 d(二叶期);分别于上述光照条件和黑暗条件下,于4 ℃春化28 d。分别在春化0、7、14、21及28 d取10株材料,重复3次。将全部叶片经过液氮速冻后,于-80 ℃保存备用。将剩余的小麦植株移栽到温室中,于22 ℃、上述光照条件下继续生长,统计抽穗时间,并于抽穗后统计植株主茎叶片数及株高。
1.2 RNA提取及cDNA合成
使用EasyPure Plant RNA Kit(北京全式金生物技术有限公司)提取1.1中-80 ℃保存叶片的总RNA,琼脂糖凝胶电泳检测其完整性,Nano Drop 2000(Thermo Fisher Scientific)检测/值及RNA浓度。将总RNA稀释至500 ng·μL,使用Prime Script RT reagent Kit with gDNA Eraser (Perfect Real Time) 试剂盒[宝生物工程(大连)有限公司]合成cDNA,将其稀释10倍作为Real-time PCR模板。
1.3 Real-time PCR
在NCBI上下载各相关参考序列,并在Ensembl Plants网站上(http://plants.ensembl.org/Multi/Tools/Blast)进行blastn分析,获得在小麦中国春3个亚基因组上的对应序列;根据这些序列,利用软件Primer 5.0设计对应的Real-time PCR引物(表1)。利用作为内参基因,在 ABI 7500 Real-Time PCR System(Thermo Scientific)上进行Real-time PCR。使用天根生化科技(北京)有限公司的SuperReal PreMix Plus (SYBR Green)试剂盒。扩增体系20 μL:2×SuperReal PreMix Plus 10 μL,50×ROX Reference Dye 0.4 μL,正反向引物(10 μmol·L)各0.6 μL,cDNA模板5 μL,RNase-Free ddHO 3.4 μL;扩增程序为:94 ℃预变性 3 min;94 ℃变性30 s,按各自退火温度退火 30 s,72 ℃延伸30 s,40个循环;溶解曲线分析程序: 95 ℃ 15 s,60 ℃ 1 min,缓慢升温至95 ℃并持续30 s,60 ℃ 15 s。数据分析采用2-△△C方法。

表1 供试引物及其序列Table 1 Primers used in this study and their sequences
2 结果与分析
2.1 光照春化与暗春化对小麦表型的影响
表型分析结果表明,春化结束时,小麦处于三叶期,暗春化及光照春化的小麦表型没有明显差异。光照春化植株的平均抽穗期为100.53 d,而暗春化为120.27 d,比光照春化晚19.73 d,二者差异极显著(< 0.01)。完成抽穗后,光照春化植株的平均主茎叶片数为6.5片,而暗春化植株的平均主茎叶片数为8.0片,二者差异极显著 (<0.01);光照春化植株的平均株高为32.43 cm,而暗春化植株的平均株高为28.86 cm,二者差异显著(<0.05)。
2.2 光照春化及暗春化对小麦春化相关基因表达的影响
在春化前表达量很低,随着春化的进行,其表达量迅速上调;在光照条件下,其上升幅度明显高于黑暗条件(图1A)。在春化前表达量相对较高,春化显著抑制该基因的表达,其在暗春化下的表达量更低(图1B)。随着暗春化的持续而逐渐上调表达,并在春化21 d时达到峰值,随后表达下调;在光照条件下,该基因在春化7 d时即达到表达峰值,至春化28 d时明显较低;该基因在暗春化条件下的表达量比光照春化更高(图1C)。在供试材料中均未被检测出。
在光照春化条件下表达水平显著高于春化前,但在暗春化条件下,仅在春化7 d时表达量显著上调,春化更长时间与春化前相比没有显著差异(图1E)。在光照春化7 d时表达水平较春化前有所下降,随春化继续进行,其表达水平恢复至春化前水平;暗春化导致其表达量较春化前显著降低(图1F)。在光照春化条件下总体表现为上调表达(仅在春化21 d时表达轻微下调),其在暗春化条件下的表达没有明显的规律(图1G)。整体而言,、、在光照春化时表达量显著高于暗春化(图1E~图1G)。

VL:光照春化;VD:暗春化。图柱上不同字母表示差异显著(P<0.05)。下同。
随着春化的持续,表达水平逐渐升高,其在光照条件下的上升幅度明显高于黑暗条件,且黑暗条件下春化14 d后其表达已趋于稳定(图1D)。
2.3 光照春化及暗春化对小麦光周期基因表达的影响
较春化前而言,在光照春化条件下,表达水平显著下降(图2A);表达仅在光照春化7 d时明显降低,其他时间与春化前无明显变化(图2B);光照春化导致表达水平显著降低,但春化28 d时其表达水平恢复到春化前水平(图2C)。在暗春化条件下,、以及表达较之春化前均显著降低,且其表达量显著低于光照春化(图2A~图2C)。
春化导致表达显著上调,其表达水平达到峰值后逐渐下降,但光照春化和暗春化下该基因表达达到峰值的时间不同(光照春化21 d,暗春化14 d),且光照春化的表达峰值高于暗春化(图2D);在光照春化及暗春化条件下表达量均显著降低,但在暗春化条件下降低程度更高(图2E);在光照春化及暗春化条件下表达量上升,且在光照春化条件下表达量上升幅度更高(图2F)。
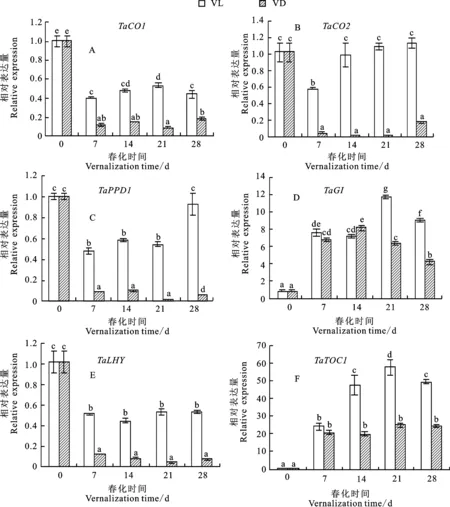
图2 光周期基因在光照春化及暗春化过程中的表达
3 讨 论
表达在光照春化和暗春化条件下均逐渐上升,但光照春化导致其上升幅度更明显(春化28 d时,光照春化表达量是暗春化条件下的4.5倍);受到光照春化及暗春化的强烈抑制,但暗春化条件下的抑制效果更强;光照春化和暗春化过程中均未检测到表达;由于抑制表达,在光照春化过程中的迅速表达受到、及低温以外因素的影响。在小麦中,TaVRN3、14-3-3和TaFDL2蛋白组成六聚体FAC复合物(florigen activation complex),结合启动子并调控其表达。在六倍体小麦中至少已经发现了46个情况基因,17个基因,以及5个基因;这些基因家族成员广泛互作,产生了大量的FAC复合体,在不同组织及发育阶段响应不同的FT-like信号;在春化过程中,可能存在这样的FT-like信号,迅速诱导的表达。另一类开花相关蛋白(MADS-box)形成同源或异源四聚体,结合靶基因启动子CArG-box元件并调控其表达。如MADS-box蛋白Vegetative to Reproductive Transition 2(TaVRT2)结合启动子,并抑制的表达;另一种MADS-box蛋白HvVRN1的染色质免疫共沉淀测序(ChIP-seq,chromatin immunoprecipitation sequencing)在大麦基因组中获得了514个含CArG-box元件的结合位点,37个包含这些位点的基因受HvVRN1的调控;鉴于启动子包含CArG-box元件,可能在春化过程中受到其他基因的诱导。还需要更多的试验,来明确光照春化过程中导致显著上调的基因。
中麦175中,被光照春化和暗春化上调,不符合其抑制开花的功能。在小麦和短柄草中,有些品种的被春化上调,有些品种被春化下调;该基因可能参与了某种反馈调节,灵活微调植物对春化作用的响应能力。CO1/CO2/PPD1可能存在相似的反馈调节:长日照、突变体背景下,突变导致小麦明显晚花;过表达导致长日照和短日照下大麦提前开花,说明是开花促进基因。长日照条件下及的同时缺失导致大麦开花显著延迟,说明是开花促进基因;本试验中、以及在光照春化条件下表达量明显高于暗春化,也说明了这三个基因促进开花。长日照条件下、和的表型贡献率说明是主要的开花促进基因,长日照条件下小麦突变体比双突变体开花明显延迟进一步证明了这个结果。然而PPD1蛋白存在时,和同时突变导致小麦在长日照和短日照条件下均提早开花,不符合它们促进开花的功能,暗示了CO1和CO2可能通过灵活微调参与的长日照促进开花过程。
在小麦中发现了3个同源基因,其中和在光照春化条件下上调表达,可能调控春化过程中组蛋白甲基化状态,促进开花。是、的同源基因;和参与了拟南芥的染色质组蛋白甲基化修饰,抑制春化过程中的表达,促进开花。在春化过程中逐渐上调,但不受春化过程调控,而被短日照条件诱导,抑制()基因表达,不依赖于促进拟南芥在短日照条件下开花。虽然和亲缘关系较近,但仅有在暗春化条件下显著下调,而与前人研究不同,在光照春化条件下其总体表达差异不明显,因而可能参与了类似于拟南芥基因不依赖于的短日照促进开花过程。
4 结 论
光照春化比暗春化更有利于小麦开花;光照春化条件下,、、、、、、、、、、和的表达水平高于暗春化。推测光照和低温通过影响生理节律基因的表达来调控下游基因的表达,其中一些基因(如、及一些未知基因)通过调控表达调控小麦开花;和表观调控有关的基因同样受到光照和低温的调控。
