中国‘苦水’玫瑰杂交育性的初步研究
2022-05-19张玉洁焦子惠李雪丽达晓伟
汪 颖 张玉洁 焦子惠 李雪丽 王 凯 达晓伟 孙 坤 张 辉
(西北师范大学生命科学学院,兰州 730070)
‘苦水’玫瑰(×Yu. et.Ku.)是玫瑰精油主产区兰州市永登县的主栽品种,以迷人的芳香而闻名于世界。‘苦水’玫瑰的栽培区每年可见“花开时繁团锦簇,秋季里不见一果”的景象,即便是在花期无人采摘的大片玫瑰园地。由于‘苦水’玫瑰在生产上主要受关注的是花,研究上主要关注其花部的各种经济性状,如花色、花香、精油及其生物活性成分等;同时,‘苦水’玫瑰多采用压条扦插等营养扩繁的生产方式,使人们一直忽视这个问题:芬芳的玫瑰谷从来不缺少蜜蜂等辛勤的传粉者,‘苦水’玫瑰为什么不会像月季一样结出丰硕的果实呢?
‘苦水’玫瑰所在的蔷薇科(Rosaceae)有多种果树,如杏()、李()和苹果()等都具有配子体型自交不亲和性,即自花授粉不能结实,在生产上必须配置授粉品种或进行人工辅助授粉才能保证坐果。蔷薇属不同植物间能进行杂交产生可育后代,且杂交和多倍化是蔷薇属植物进化的主要因素。现代月季多由品种间杂交和远缘杂交而来,亲缘关系越近杂交亲和性越好。李玉舒通过对‘堇蝶’(‘Jindie’)、‘婉粉’(‘Wanfen’)、‘玉蕊霞光’(‘Yurui Xiaguang’)等栽培玫瑰品种的花瓣数目与结实性的比较研究,发现玫瑰的结实性与花瓣的数目有关,也与雄蕊瓣化的程度有关。蔷薇属大部分植物的染色体倍性都已经陆续报道,‘苦水’玫瑰染色体为14条,属1A型,但关于其小孢子母细胞减数分裂相关内容却鲜见报道。染色体的数目和结构决定植物的生殖过程,减数分裂过程的稳定保证了后代遗传的稳定性和连续性,其中任何异常均会影响花粉育性。另外,雄性不育也是植物种内生殖障碍的重要机制之一。为此,本研究通过遗传杂交实验研究了‘苦水’玫瑰的有性生殖水平,对‘苦水’玫瑰的花部特征及花粉的形态与功能进行了观察研究,同时细胞学层面观察小孢子的发生过程,以揭示‘苦水’玫瑰千花无果这一现象的内在原因。
1 材料与方法
1.1 试验材料
选择‘苦水’玫瑰以及4 个其他蔷薇属的种(品种)为试验材料(见表1),各供试材料都经过形态特征和cpDNA分子水平的鉴定。

表1 玫瑰材料Table 1 A list of rose materials studied
1.2 试验方法
‘苦水’玫瑰花期在4 月底至6 月初,5 月为盛花期,与‘丰花’玫瑰、‘四季’玫瑰和‘艳霞’玫瑰的花期基本重合,与重瓣黄刺玫的末花期相遇,授粉材料在花蕾成熟至外层花被微开裂时标记待用。自交材料直接套袋隔离,次日(上午10:00前)人工辅助自交授粉后套袋挂牌。异交双亲材料用镊子去雄,收集花药于1.5 mL 离心管,母本材料避免伤及柱头,用硫酸纸袋保护隔离;花药散粉后用棉球蘸取授予柱头,套袋标记。‘苦水’玫瑰自交授粉50个,杂交每个组合不少于10个。精细田间管理,记录落果情况,于8月果实着色后采摘。阴干后解剖果实,种子计数。
对供试的5个材料各取15朵盛开花朵,分离观察花瓣、雄蕊及雌蕊等三轮花器官的形态并计数,其中瓣化雄蕊表现为雄蕊畸形,瓣化有翅状结构,边缘或顶端残存黄色花药。定义:雄蕊瓣化率=瓣化雄蕊数量/(瓣化雄蕊数量+正常雄蕊数量),利用SPSS 21 检验数据的方差齐性,完成雄蕊瓣化率的单因素方差分析,比较材料之间的均值差异性。
除黄刺玫外,供试的其余4 种材料随机选取3朵花上的各10 个将裂花药,置于离心管中干燥。完全放粉后,在离心管中加入1 mL 2.5%的蔗糖溶液,吸取0.5 μL 的花粉液在100 倍显微镜下选取3个视野进行观察。根据花粉颗粒的饱满程度,区分大花粉和小花粉,分别计数。参考1.2.2 观察结果,推算花粉胚珠比(P/O 值)以及大花粉与胚珠之比(BP/O值)。
参考王琼等提供的栽培玫瑰花粉萌发的适宜条件,在0.3 g·L的蔗糖溶液中对上述4 种供试材料进行花粉悬滴培养,分别做3个重复对照。将花粉置于垫有湿滤纸的培养皿中,于人工气候箱中25 ℃培养。1、2、3、15、24 h,各时间段选取3 个400倍视野统计花粉萌发数量,拍照。
于4月下旬现蕾时开始采集处于不同发育阶段的幼嫩花蕾若干。用卡诺试剂((无水乙醇)∶(冰醋酸)=3∶1)固定12~24 h,固定好的材料用75%的乙醇清洗后再置于75%的乙醇于4 ℃保存。
参考李懋学的常规花粉染色观察,改良石炭酸品红溶液染色。将保存的花蕾于解剖镜下剖离花药,花药在10%HCL 中室温解离15 min,蒸馏水漂洗后石炭酸品红染液染色20 min。切碎花药盖上盖玻片并轻敲盖玻片使细胞分散,最后封片用于镜检。光学显微镜拍照记录减数分裂的整个过程及其中出现的异常分裂现象。
2 结果与分析
2.1 ‘苦水’玫瑰的授粉结实性
‘苦水’玫瑰的自然授粉与人工辅助自交授粉均无法成功坐果,具有严格的不结实性(见表2)。授粉初期可见子房微微膨大,数天后萎蔫从果柄处脱落。

表2 ‘苦水’玫瑰的遗传杂交Table 2 Genetic hybridizations involving R. sertata × R.rugosa
收集‘丰花’玫瑰、‘四季’玫瑰、‘艳霞’玫瑰及重瓣黄刺玫的花粉,授在去雄‘苦水’玫瑰的柱头上,7~10 d 可见明显肉质浆果,授粉2 月后果皮着色,解剖后可见颗粒饱满的种子(见图1A)。4 种供试材料中,‘苦水’玫瑰与‘艳霞’玫瑰的杂交结籽数最高,平均每个浆果含16.33 个种子。‘苦水’玫瑰与重瓣黄刺玫初步完成的8次杂交试验,其结果与其他材料结果相似,即‘苦水’玫瑰做母本有结实,果实大小如图1B所示。

图1 ‘苦水’玫瑰为母本的异交结实情况A.KSRR×YXRR的果实及种子;B.KSRR×CBRX的果实Fig.1 Production of cross pollination for style of‘Kushui’rose A.Fruits and seeds from KSRR×YXRR;B.Fruits from KSRR×CBRX
‘苦水’玫瑰的花粉授到‘丰花’玫瑰、‘四季’玫瑰、‘艳霞’及重瓣黄刺玫的柱头上,子房膨大后可留存多日,内无正常发育的种子,均无法获得异交果实。
2.2 花部特征观察
在4 种玫瑰材料的花部发现均有雄蕊花瓣化现象存在,图2A 为‘苦水’玫瑰的三轮花器官形态,第3行为花瓣化的雄蕊结构。Levene统计检验瓣化百分率的方差齐性,在未假定方差齐性的情况下利用Dunnett T3进行两两比较,发现‘四季’玫瑰雄蕊的瓣化程度最低,雄蕊瓣化百分比均值为1.54%,其余3 种品种玫瑰都具有较高的雄蕊花瓣化现象,‘艳霞’玫瑰达到了6.58%,‘丰花’玫瑰和‘苦水’玫瑰分别为5.06%和5.74%,三者之间差异不显著(见图2B)。
4种玫瑰的雌蕊群均由多个离生心皮组成,每个心皮含有1个球形胚珠,图2B中的“雌蕊数”即雌蕊群的心皮数目就等于胚珠数,而“雄蕊数”并不包含花瓣化的雄蕊,因为瓣化雄蕊中并无成熟的花粉粒释放出来。根据花粉计数和雌蕊数目计算4种玫瑰每朵花的花粉胚珠比(pollen-ovule ratio,P/O),结果均为2×10左右。由于供试材料的花粉饱满度百分率和P/O为方差齐性,在假定方差齐性的情况下利用Gabriel进行两两比较,发现4种玫瑰材料的P/O 均值之间没有显著的差异,‘苦水’玫瑰为2.17×10,‘艳霞’玫瑰、‘丰花’玫瑰和‘四季’玫瑰分别为2.53×10、2.17×10和1.91×10(见图3B)。
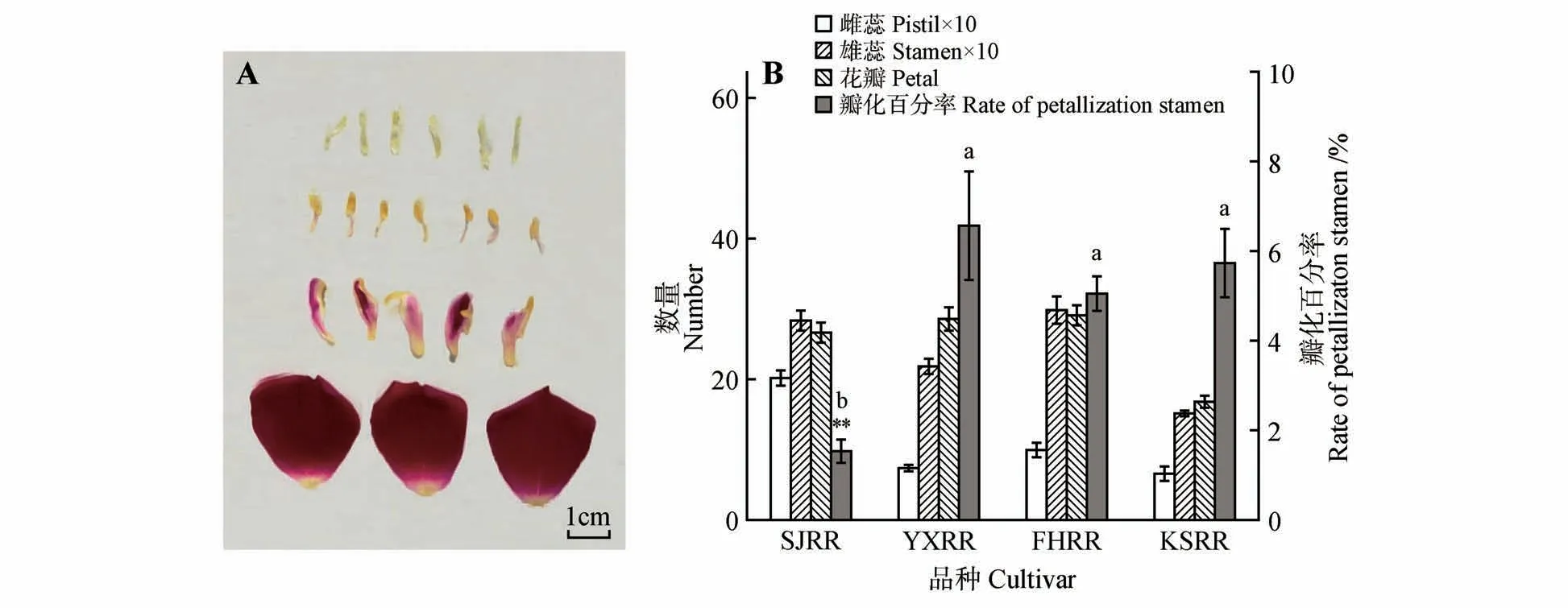
图2 ‘苦水’玫瑰的花器官形态(A)及不同材料的雄蕊瓣化状况(B)不同小写字母表示差异显著(P<0.05);**表示差异极显著(P<0.01)Fig.2 Morphology of floral organ of‘Kushui’rose and petallization of stamens in four materials Different lowercase letters indicate significant difference(sP<0.05);**means extremely significant difference(P<0.01)
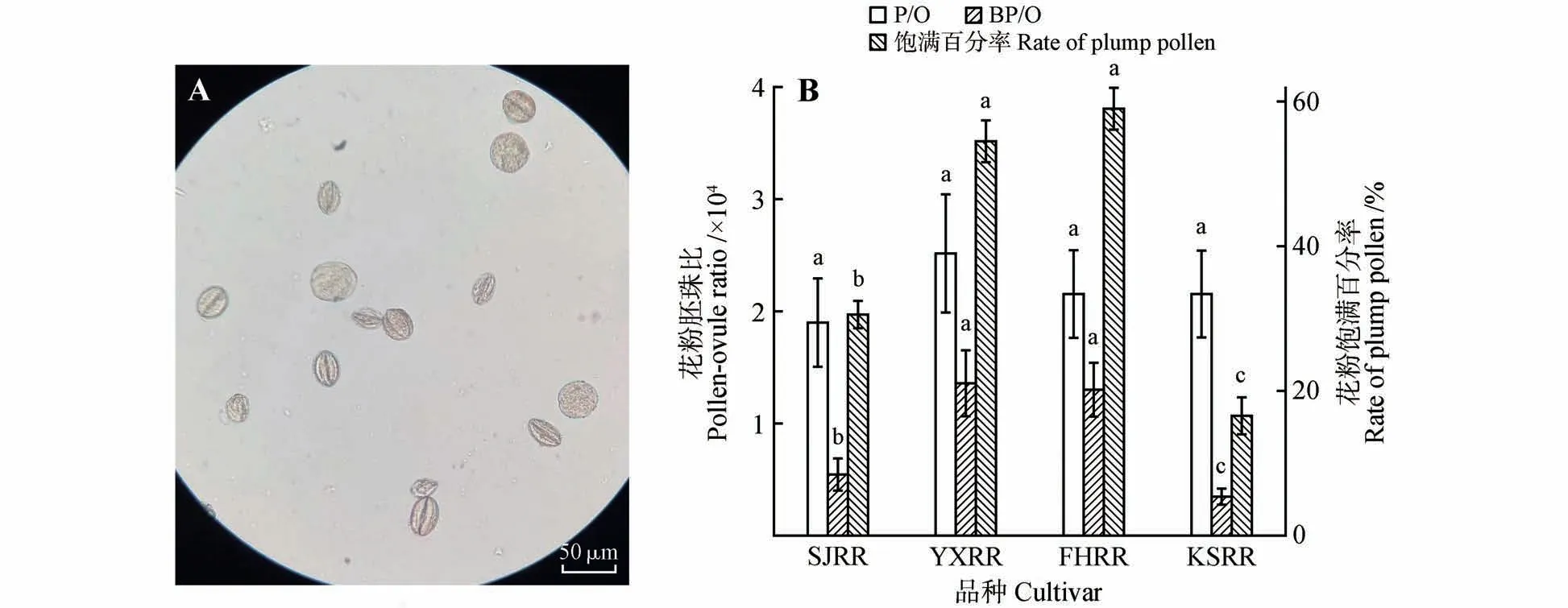
图3 ‘苦水’玫瑰花粉异型性和不同玫瑰花粉败育情况A‘.苦水’玫瑰花粉异型性(400×);B.不同玫瑰材料的花粉败育状况Fig.3 Pollen heteromorphorsis of‘Kushui’rose and pollen abortion of different materialsA.Pollen heteromorphorsis of KSRR(400×);B.Pollen abortion of different materials
2.3 花粉异型性及萌发率
4种玫瑰的花粉都有败育现象,花粉异型现象明显。图3A 显示‘苦水’玫瑰除了颗粒饱满的“大花粉”之外,还存在一系列大小不等的“小花粉”。比较而言,其余3种玫瑰的花粉都可以分为差异非常明显的“大花粉”和“小花粉”,‘艳霞’玫瑰的花粉形态如图4A 所示。统计花粉饱满百分率显示‘苦水’玫瑰的数值在4 种玫瑰中为最小,仅为16.73%,‘四季’玫瑰次之为30.86%,‘艳霞’玫瑰和‘丰花’玫瑰分别达到了54.91%和59.43%(图3B)。仅把大花粉计算在内的大花粉胚珠比(Big pollen/ovules ratio,BP/O ratio)在4种材料中从小到大分别为0.36×10、0.55×10、1.31×10和1.37×10(见图3B)。在未假定方差齐性的情况下各材料两两比较花粉饱满百分率和BP/O 值,两个统计参数除‘丰花’玫瑰与‘艳霞’玫瑰差异不显著外,‘四季’玫瑰和‘苦水’玫瑰与其他材料都存在极显著的差异(图3B)。
除‘苦水’玫瑰外,其余3 种材料在0.3 g·L蔗糖悬滴中培养的花粉,1 h 后就可观察到萌发现象,凸起处花粉壁变薄;2 h 后可见透明状花粉管形态,图4A 为‘艳霞’玫瑰在2 h 时于400×视野下的镜检结果;15 h 后3 种玫瑰花粉萌发率达到最大,‘四季’玫瑰单个视野统计结果可以达到44.44%。‘苦水’玫瑰花粉的各时段检测结果显示都没有萌发,仅在15 h 时于一个视野下观察到一个花粉粒吸胀的情形(见图4B)。‘苦水’玫瑰的花粉吸胀处花粉壁界线不清晰,破裂后有内容物流出,这与图4A 所示‘艳霞’玫瑰萌发的花粉管具有明显的不同。如图5所示,统计玫瑰材料在15 h和24 h的萌发率得到数据均值,在未假定方差齐性的条件下检测到‘四季’玫瑰具有显著高于‘丰花’玫瑰和‘艳霞’玫瑰的萌发率,分别为20.44%、11.71%和11.01%,而‘苦水’玫瑰的萌发率近似为0。
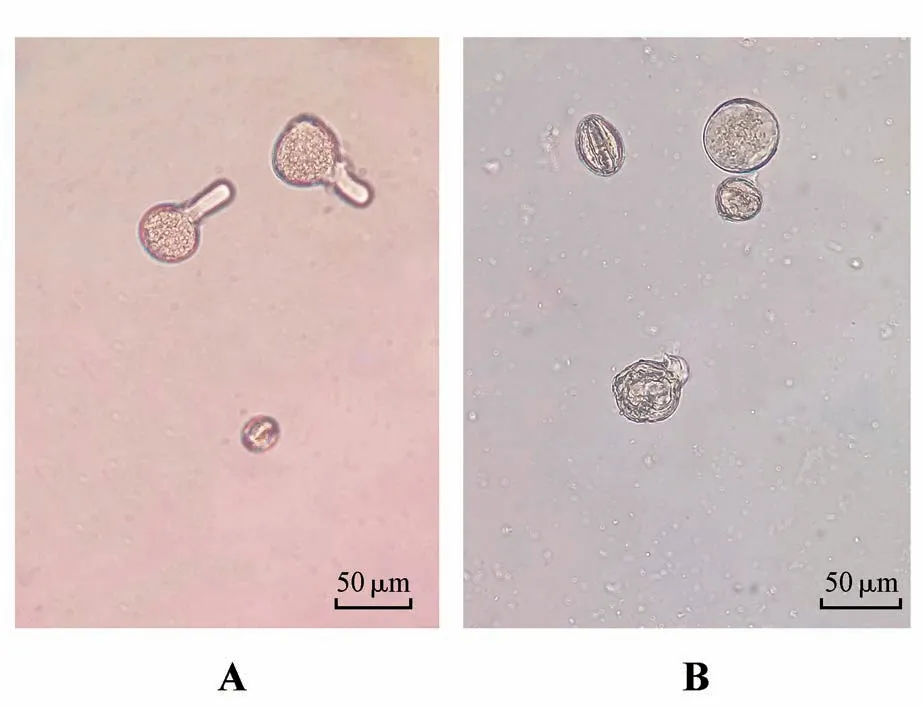
图4 玫瑰花粉在3%蔗糖溶液中的萌发情况A.萌发的‘艳霞’玫瑰花粉(2 h,400×);B‘.苦水’玫瑰花粉的吸胀现象(15 h,400×)Fig.4 Pollen germination of rose in 3%sucrose solution A.Germination of YXRR pollens(2 h,400×);B.Bulges of KSRR pol⁃len(s15 h,400×)

图5 玫瑰材料萌发率统计Fig.5 Germination rates of rose materials
2.4 小孢子母细胞减数分裂行为观察
通过对‘苦水’玫瑰小孢子母细胞减数分裂后期Ⅰ(见图6:Ac)和‘四季’玫瑰减数分裂前期Ⅰ(见图6:Bj)的染色体观察,发现两种玫瑰染色体均为14 条(2n=2x=14)。两种栽培玫瑰小孢子母细胞发育过程中均存在减数分裂各时相不同步的现象,同一花蕾不同花药的发育程度不同,同一个花药中的不同小孢子母细胞减数分裂进程不一致(见图6:Bo),减数分裂的不同步程度严重。
本研究观察到了‘苦水’玫瑰和‘四季’玫瑰几乎所有的小孢子母细胞减数分裂时相(图6箭头所示)。‘四季’玫瑰小孢子母细胞外形圆润,减数分裂过程中染色体协调分配,胞质均匀分裂,属正常减数分裂行为(见图6B)。与‘四季’玫瑰不同的是,‘苦水’玫瑰的减数分裂过程中观察到细胞核与细胞质均有异常分裂现象。‘苦水’玫瑰中期Ⅰ(见图6:Ab)、后期Ⅰ(见图6:Ac)、中期Ⅱ(见图6:Af)等多个时相中都可观察到减数分裂细胞外形有一定程度的不规则形变。后期Ⅰ(见图6:Ac)、末期Ⅱ(见图6:Ag)、三分体(见图6:Ai)等多个时相染色体的着色程度深浅不一,范围不等,染色体分配行为具有明显的不协调性。‘苦水’玫瑰中期Ⅱ(见图6:Af)和四分体时期(见图6:Ah)中可观察到成对的次级小孢子母细胞及四分体的细胞质着色范围大小不一,表明存在胞质不均匀分裂。这种染色体和细胞质分裂的不均等会直接导致‘苦水’玫瑰减数分裂的产物中存在大量异常的四分体,甚至有明显的三分体(见图6:Ai)。

图6 小孢子母细胞减数分裂过程(1 000×)A‘.苦水’玫瑰减数分裂过程(a.前期Ⅰ;b.中期Ⅰ;c.后期Ⅰ;d.末期Ⅰ;e.前期Ⅱ;f.中期Ⅱ;g.末期Ⅱ;h.四分体;i.三分体);B‘.四季’玫瑰减数分裂过程(j.前期Ⅰ;k.中期Ⅰ;l.后期Ⅰ;m.末期Ⅰ;n.末期Ⅱ;o.减数分裂的不同步性)Fig.6 Meiosis process of pollen mother cell(s1 000×)A.Meiosis process of KSRR pollen mother cell(sa.Prophase Ⅰ;b.Metaphase Ⅰ;c.Anaphase Ⅰ;d.Telophase Ⅰ;e.Prophase Ⅱ;f.Metaphase Ⅱ;g.Telophase Ⅱ;h.Tetrad;I.Triad);B.Meiosis process of SJRR pollen mother cell(sj.Prophase Ⅰ;k.Metaphase Ⅰ;l.Anaphase Ⅰ;m.Telophase Ⅰ;n.Telophase Ⅱ;o.Non−synchronism of meiosis)
3 讨论
3.1 苦水玫瑰的雄性不育导致千花无果现象
单瓣玫瑰、复瓣玫瑰和多数重瓣玫瑰都具有很好的结实能力,但李玉舒对于自己命名的“紫缎”和“洒金紫缎”这两个‘苦水’玫瑰品种都记载为“基本不结实”,她推测玫瑰的雄蕊瓣化程度与其结实性存在负相关性,本研究显示‘苦水’玫瑰与其他油用玫瑰的雄蕊花瓣化程度没有显著差异,处于同一水平,推测对其结实性能的影响不大。
‘苦水’玫瑰所在的蔷薇科具有配子体型自交不亲和性,同属蔷薇亚科的草莓属植物更是具有两个控制自交不亲和的S-位点。本研究发现‘苦水’玫瑰作为母本能够正常授粉结实,而自交或作为父本与其他任何玫瑰品种间杂交都无法正常受精结实,表明存在明显的花粉异常或雄性不育现象,同时,与重瓣黄刺玫的远缘种间杂交的初步结果也支持该结论。同时,据柏斌等报道‘苦水’玫瑰的杂交育种实践中也把其确定为雄性不育系。所以,雄性不育造成了’苦水‘玫瑰园区千花无果的独特景象,而并非是由蔷薇科常见的自交不亲和性导致。当然,‘苦水’玫瑰雄性不育的这种上位效应一定程度会掩盖其自交不亲和性的表现,‘苦水’玫瑰是否具有完整的S-位点以及正常的S-基因并不清楚,还需进一步在基因层面做深入研究。
3.2 ‘苦水’玫瑰的小孢子减数分裂异常造成了花粉败育
‘苦水’玫瑰具有与‘丰花’、‘四季’及‘艳霞’等玫瑰品种相似的花粉胚珠比(P/O 值),这说明在提供花粉数量方面它们具有相近的供粉效率,而受粉结实性能的差异主要取决于花粉质量。李玉舒等发现玫瑰品种中都不同程度地存在花粉败育现象,败育的花粉粒较正常的花粉粒小,内凹,空瘪呈畸形,这与在本研究各种玫瑰材料中观察到的现象相一致。然而,与其他材料仅具有大花粉和小花粉两种差别明显的类型不同,‘苦水’玫瑰的花粉镜检结果显示除了正常的大花粉之外,还存在从大到小连续变异的异常花粉,这种花粉败育极大地降低了‘苦水’玫瑰作为父本在杂交授粉时的贡献率。
染色体压片试验中观察到‘苦水’玫瑰和‘四季’玫瑰的染色体均为14条,与招雪晴等的结果相一致,且两种玫瑰的减数分裂各时相进程有严重的不同步性,该特性是蔷薇属植物普遍存在的减数分裂现象。陈瑶等认为植物小孢子母细胞减数分裂异常是导致花粉败育的主要原因,在减数分裂的前期、后期、末期等任何一时期发生异常都会导致小孢子不能正常发育,降低了花粉育性。本研究发现小孢子母细胞减数分裂时染色体分配不协调,胞质不均等分裂等因素都影响了‘苦水’玫瑰正常的四分体发生和发育,甚至变为三分体,推测种种异常的减数分裂行为是造成‘苦水’玫瑰花粉败育的主要原因,而且这种花粉败育严重妨碍了其在杂交育种中做父本时的结实率,从而导致其具有雄性不育的特性。当然,植物花粉形成和发育过程中绒毡层也起着十分重要的作用,如果绒毡层功能失常,出现液泡化,过度肥大等现象,均会导致小孢子母细胞减数分裂不能正常进行。同时,异常的纺锤体方向被认为是三分体形成的原因,细胞不均等分配也可能形成三分体或多分体。然而,‘苦水’玫瑰减数分裂过程中绒毡层细胞如何变化?其小孢子母细胞异常受哪一种雄性不育机制控制?仍需要进一步的深入研究。
3.3 ‘苦水’玫瑰的雄性不育与遗传杂交育种
生产上,‘苦水’玫瑰主要依靠压条扦插等方式营养扩繁,长期的无性繁殖也导致‘苦水’玫瑰出现不同的自然变异类型,而且在外观形态和内在香气成分方面都具有明显的差异,这种自然分化的性状变异在‘苦水’玫瑰育种实践中的应用尚需进一步探索。本研究发现花粉败育导致‘苦水’玫瑰具有雄性不育现象,但其柱头具有正常可授性,花柱能够正常引导花粉管生长,特别是胚珠中具有正常可受精的卵细胞。所以,‘苦水’玫瑰可以作为母本进行杂交育种改良种质,这也是柏斌和牛元等能够通过常规的杂交育种技术获得大花型‘苦水’玫瑰新品种的原因。
