低温胁迫对柳杉不同无性系的影响及抗寒性评价
2022-05-10崔洁冰张萌张莹婷徐进
崔洁冰 张萌 张莹婷 徐进
(南京林业大学林木遗传与生物技术省部共建教育部重点实验室 南京林业大学南方现代林业协同创新中心 南京林业大学林学院,南京 210037)
柳杉(Cryptomeria fortunei)为杉科(Taxodiaceae)柳杉属(Cryptomeria)常绿高大乔木,喜山地气候及条件良好的酸性土壤,具有侧根发达、生长迅速、适应性强、树形优美且能净化空气、改善环境等特性[1-2],是中国南方优良的用材和园林树种。天然种群集中分布于中国东南的天目山、武夷山、日本本州岛及九州岛等地,长江以南省区也多有栽培[3]。但其生长和分布常受到干旱、寒冷等极端环境的制约,随着全球气候变化以及极端天气频繁发生,柳杉的生长发育受到了极大的影响。
低温是影响自然界植物生存的主要环境因子之一,也是限制植物地理分布的关键因素[4]。一般来说,温度降低时会导致植物的各种生理活动减慢,生长发育变缓,当低温造成的伤害超过一定程度时,环境胁迫就会对植株造成不可逆的伤害,甚至死亡[5-6]。植物为了抵御低温胁迫,其根茎叶在形态上存在着相应的响应机制,如叶形、叶片厚度、叶色和解剖结构等方面的变化[7-8]。其次,植物体内多种渗透调节物以及保护酶等在维持细胞结构及生理功能、抵抗恶劣气候条件等方面具有重要作用[9],并通过一系列生理生化代谢反应提高对环境的适应能力。
柳杉的抗寒性研究可为其品种选育提供理论依据,对于提高林木生产力与地区经济效益、促进我国生态环境的建设具有重要意义[10]。但是目前柳杉的研究主要集中在育苗、良种选育、病虫害防治与生理生态等方面,关于柳杉不同无性系的抗寒性研究相对较少。本研究测定了10个无性系在低温胁迫下的丙二醛(malondialdehyde,MDA)含量、超氧化物歧化酶(superoxide dismutase,SOD)活性及叶绿素荧光参数(Fv/Fm、Y(II)、qP)等生理生化指标,采用主成分分析结合隶属函数计算对柳杉各无性系抗寒性差异进行综合评价,以期进一步探寻柳杉无性系抗寒能力评价体系,为柳杉抗寒无性系的评价筛选及造林栽培提供理论依据。
1 材料与方法
1.1 材料
10个柳杉无性系主要来源于我国东部柳杉分布区(表1),穗条均采自南京林业大学白马实验基地柳杉优良无性系收集区,2018年6月进行扦插,2020年用于本研究。

表1 材料及来源Table 1 Materials and sources
1.2 方法
1.2.1 低温处理 取生长一致的10个柳杉无性系的2年生幼苗枝条,将每个无性系的枝条分为7组,分别放入海尔程控冰箱中进行4℃、0℃、-4℃、-8℃、-12℃、-16℃和-20℃低温处理,从4℃开始,以4℃/2 h降温,到达每个目标温度时处理12 h。零上温度处理后直接进行生理指标的测定,零下温度处理结束后放入4℃中解冻12 h后进行测定,每个处理设置3个重复。
1.2.2 测定方法 取针叶0.5 g,通过0.05 mol/L磷酸缓冲液(pH7.8)获得粗酶提取液。采用硫代巴比妥酸法[11]和氮蓝四唑比色法[11]分别测定MDA含量和SOD活性。柳杉枝条经低温处理后,使用便携式光纤型双通道调制叶绿素荧光仪DUAL-PAM/F(德国Walz公司)测定叶绿素荧光参数PS II最大光化学效率(Fv/Fm)、PS II的实际光合量子产量(Y(II))和光化学淬灭系数(qP),每个处理3次重复。
1.2.3 数据分析 (1)各项指标数据采用Excel 2010和SPSS 22进行统计与分析,使用Duncan检验法进行差异显著性分析。
(2)权重计算:本课题组Zhang等[12]在分析半致死温度时发现-4℃是一个关键的转折温度,因此以-4℃下各指标的值作主成分分析。综合相对电导率(relative conductivity,REC)、MDA含量、SOD活性及Fv/Fm 4个指标(REC数据见Zhang等[12]),使用SPSS 22软件将数据进行标准化处理,通过主成分分析确定其特征值、负荷量与贡献值,计算权重(weight,W)。
(3)隶属函数值计算:根据各无性系测定的指标数据,计算其隶属函数值并进行排序,对负向指标采用反隶属函数计算[13-14]。
隶属函数 :f(xij)=(xij-xjmin)/(xjmax-xjmin)
反隶属函数 :f(xij)=(xjmax-xij)/(xjmax-xjmin)
式中,f(xij)表示i无性系j指标的抗寒隶属函数值;xij表示i无性系j指标的测定值;xjmax表示所有无性系j指标的最大值;xjmin表示所有无性系j指标的最小值。
(4)抗寒性评价:根据各指标权重及隶属函数值,通过公式Y=Wij×f(xij)计算各无性系抗寒性综合指数Y(Wij为各抗寒指标权重;f(xij)为各指标隶属函数值)[13-14]。再结合相对电导率(REC)、MDA含量、SOD活性以及叶绿素荧光参数(Fv/Fm)等指标进行综合分析,建立抗寒性评价体系。
2 结果
2.1 低温胁迫对柳杉各无性系生理生化指标的影响
2.1.1 低温胁迫对柳杉各无性系中MDA含量的影响 由图1和附表1可知,柳杉不同无性系的MDA含量总体呈上升-下降-上升的趋势。但柳杉各无性系在不同温度时,MDA含量的变化不太一致,在一定温度范围内具有波动性。在0℃、-8℃、-16℃出现了峰值,在0℃和-16℃后呈明显的下降趋势,其中在-4℃、-8℃、-12℃时,MDA含量相较于0℃呈现下降趋势,但各无性系变化无明显规律,说明柳杉不同无性系对低温的应答反应不同,且MDA含量受多种因素影响。在整个低温处理过程中,66#无性系的MDA含量偏低,74#无性系的MDA含量偏高,说明66#无性系在低温胁迫下膜功能受损害的程度较低,抗性较强。

图1 不同低温胁迫对柳杉各无性系MDA含量的影响Fig.1 Effects of different low temperature stress on MDA content in different clones of C. fortunei
2.1.2 低温胁迫对柳杉各无性系超氧化物歧化酶(SOD)含量的影响 由图2和附表2可知,在低温胁迫下,多数柳杉无性系的SOD活性变化呈下降-上升-下降的趋势。在4℃降到0℃时,由于植株对低温的不适应,SOD活性呈现下降趋势,随后植株经过短暂的抗寒锻炼,抗寒能力逐渐增强,SOD活性出现短暂的上升,-8℃后,SOD活性逐渐下降,说明过低的温度超过了SOD酶作用的范围,植株自身抗氧化系统受到破坏,抗寒能力减弱。特别的是25#和74#无性系,呈现先上升后下降的趋势。在整个低温处理过程中,68#无性系的SOD活性值偏高,而且波动较小,抗性较强。42#无性系的SOD活性值下降较快且降幅较大,说明在低温处理过程中,42#无性系的超氧化物歧化酶受温度的影响比较大,抗性较弱。

图2 不同低温胁迫对柳杉各无性系SOD含量的影响Fig.2 Effects of different low temperature stress on SOD content in different clones of C. fortunei
2.1.3 低温胁迫对柳杉各无性系叶绿素荧光参数的影响
2.1.3.1 低温胁迫对柳杉各无性系Fv/Fm的影响 由图3和附表3可知,随着处理温度的降低,柳杉各无性系的Fv/Fm值呈上升-下降的变化趋势。在0℃时,Fv/Fm开始下降,-4℃时急剧降低,到-20℃逐渐平缓,说明柳杉在经历低温初期时有抵御能力,能够在一段时间内维持光合效率,超过抵御的临界点,其光合效率大幅度降低。其中,42#无性系的Fv/Fm变化幅度较大,且显著低于其他无性系,说明其PS II反应中心光能转化效率较低,对低温胁迫较为敏感,抗寒性较差。68#无性系的Fv/Fm值在4℃至-4℃时相对较高,而57#无性系在低温初期变化平缓且随温度的降低整体水平降幅较小,说明它们在低温胁迫下PS II损伤较小,光能转化效率较高。

图3 不同低温胁迫对柳杉各无性系Fv/Fm的影响Fig.3 Effects of different low temperature stress on Fv/Fm in different clones of C. fortunei
2.1.3.2 低温胁迫对柳杉各无性系Y(II)的影响 由图4和附表4可知,在低温处理下柳杉各无性系Y(II)与Fv/Fm的变化趋势相似,随着温度的降低整体呈上升-下降的变化趋势。Y(II)在低温初期出现小幅上升,在0℃后呈下降趋势,随着温度的降低,在-4℃以后急剧下降,-8℃后变化逐渐平缓,-20℃时各无性系Y(II)值趋近于0。其中,68#无性系在0℃达到峰值,在-4℃之后急剧下降,且变化幅度较大,说明68#在低温初期抗性较强,却对温度较为敏感。57#无性系的Y(II)值随温度降低也明显下降,但整体上比其他无性系下降幅度小。由图可以看出,42#无性系在-8℃和-16℃时Y(II)值趋近于0,各无性系在-20℃时的Y(II)值也几乎为0。Y(II)是PS II实际光合量子产量,一段时间内持续地对光能的有效转换能力[15]。Y(II)为0,说明柳杉PS II反应中心的实际光能转换能力在低温胁迫下受到抑制,柳杉的光合结构受到破坏。

图4 不同低温胁迫对柳杉各无性系Y(II)的影响Fig.4 Effects of different low temperature stress on Y(II)in different clones of C. fortunei
2.1.3.3 低温胁迫对柳杉各无性系qP的影响 由图5和附表5可知,柳杉各无性系qP值在4℃-0℃无显著变化,从0℃开始缓慢下降,-4℃时qP值急剧降低,在-8℃后变化平缓,直至-20℃ qP值趋近于0,这与Y(II)值的变化相似。由图5可以看出,qP值随温度的降低呈现下降趋势。其中,74#无性系的qP值在低温初期变化不明显且处于较高水平,57#和32#无性系随温度的变化qP值降幅较小,在低温后期,qP值高于其他无性系,说明74#、57#和32#无性系光合电子传递能力较强,PS II受害程度较轻,抗性较好。42#无性系qP值变化与Y(II)值变化相似,显著低于其他无性系且趋近于0,说明对光能的利用能力较弱,抗性较差。

图5 不同低温胁迫对柳杉各无性系qP的影响Fig.5 Effects of different low temperature stress on qP in different clones of C. fortunei
2.2 主成分分析
结果如表2所示,遵循特征值大于1的原则,从4项生理指标中共提取出2个主成分,特征值分别为2.217和1.056,贡献率分别为55.419%和26.389%,累计贡献率达81.809%,因此,结果理想,提取的2个主成分概括了4个生理指标的绝大部分信息。运用2个主成分对4个生理指标进行负荷量分析和权重的计算,由表3可以看出,各指标对柳杉抗寒性影响由强到弱分别是相对电导率(REC)、MDA、Fv/Fm、SOD。

表2 主成分分析结果Table 2 Results by principal component analysis
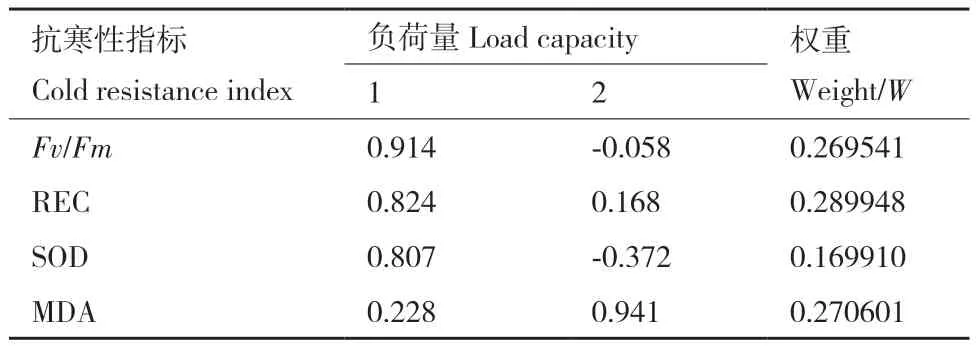
表3 柳杉各无性系抗寒性的负荷量及权重Table 3 Load capacities and weights of cold resistance indicators of C. fortunei clones
2.3 抗寒指标隶属函数值
以相对电导率(REC)、MDA含量、SOD活性及叶绿素荧光参数(Fv/Fm)为评级指标,计算隶属函数值并进行排序。结果(表4)表明,无性系57#、68#和32#的隶属函数值较大,表明其抗寒评分较高,抗寒性较强,3#、25#和42#无性系隶属函数值较小,表明其抗寒评分较低,抗寒性相对较弱。
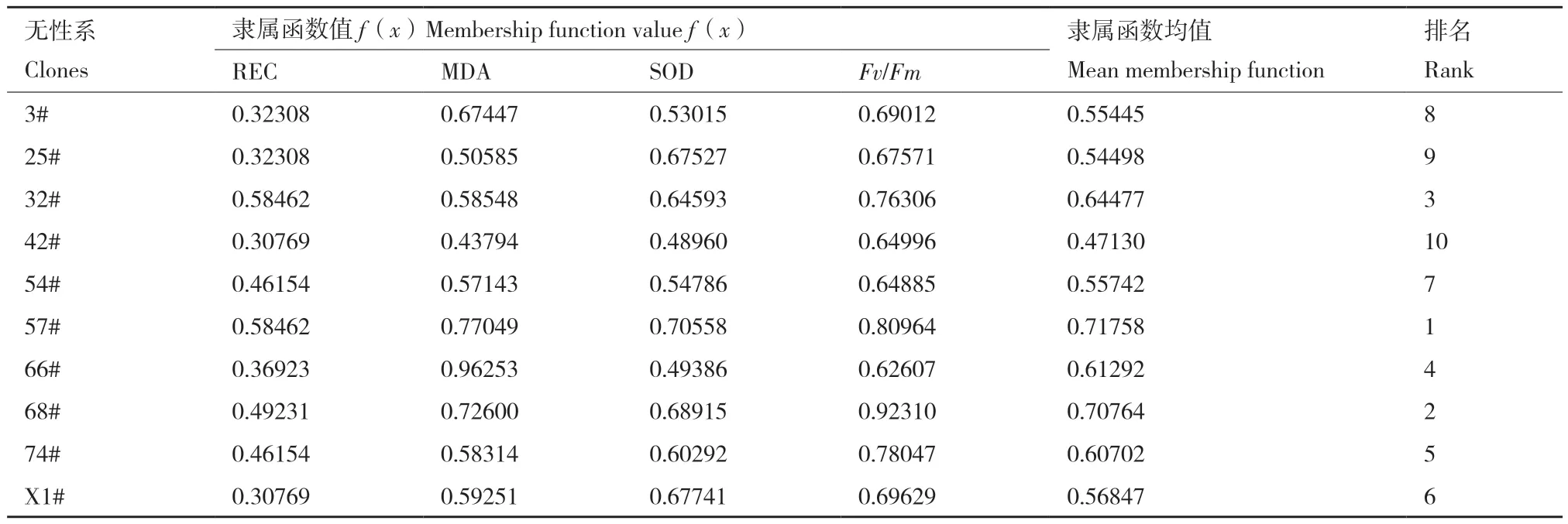
表4 柳杉各无性系抗寒性指标的隶属函数值Table 4 Membership function values of cold resistance index of C. fortunei clones
2.4 柳杉各无性系抗寒性综合评价
以相对电导率(REC)、MDA含量、SOD活性及叶绿素荧光参数(Fv/Fm)为评级指标,依据权重与隶属函数值,计算柳杉各无性系抗寒性综合指数Y,进而评价柳杉各无性系的抗寒性(表5)。结果表明,57#、68#和32#为抗寒性较强的无性系,3#、25#和42#无性系抗寒性较弱。各无性系的综合指数有一定的差异,其综合指数为0.46-0.72,抗寒性最好的无性系为57#,其次为68#、32#、66#和74#,抗寒性最弱的无性系为42#,其余几个无性系的综合指数相对来说差距不大,为0.52-0.56。

表5 柳杉各无性系抗寒性综合评价Table 5 Comprehensive evaluation of cold resistance in C. fortunei clones
3 讨论
细胞膜是细胞感受环境胁迫最敏感的部位,当植物受到低温胁迫时,细胞膜首先受到伤害,透性增大,使细胞内的电解质大量外渗。外渗液电导值的变化,可反映材料的细胞膜伤害程度,进而判断植物抗寒性的大小[16-18]。Zhang等[12]在研究柳杉不同无性系低温胁迫时相对电导率发现,电导率整体呈“S”型曲线,呈现先上升后基本不变的趋势。10个柳杉无性系的半致死温度在-4.7℃--8℃。其中57#和32#在整个过程中相对电导率相对于其他无性系较低,抗寒能力较强,42#相对电导率增幅最大,说明受低温的影响较大,抗寒能力较弱。
此外,植物在逆境条件下往往发生膜脂过氧化作用,其产物MDA会严重损伤生物膜,通常利用它作为膜脂过氧化指标,表示细胞膜脂过氧化程度及对逆境反映的强弱[19]。本研究表明在低温胁迫下,MDA含量随着温度的降低在一定温度范围内存在波动性,但总体来说呈现先上升后下降再上升的趋势,表明在低温胁迫下,柳杉细胞膜功能受到抑制,随着温度的降低MDA在细胞内积累,膜功能受阻或丧失,造成细胞代谢紊乱,从而对细胞产生伤害。柳杉各无性系MDA的含量没有明显的变化规律,其原因可能有两个方面原因,一是MDA含量与某些植物的抗寒性并无太大关系,如王微[20]对紫花苜蓿几个抗寒生理指标的研究中也出现MDA含量无变化规律的现象,她认为抗寒反应的生理机制比较复杂,单从某一个生理指标来鉴定抗寒性是比较困难的;二是试验中采用离体枝条针叶进行测定,可能与整株植株生理指标变化有差异;三是整个低温处理过程在黑暗的冰箱中进行,没有考虑光照对生理指标的影响。
过氧化氢(H2O2)和超氧阴离子(O2-)是逆境情况下产生的活性氧(ROS)物质,低温胁迫下植物会增加细胞内活性氧物质的含量[21]。超氧化物歧化酶(SOD)是生物防御系统的关键酶,在植物体内主要是用于清除超氧阴离子自由基等活性氧物质,维持植物体内活性氧代谢平衡,从而对机体起到保护作用[22-23]。因此,SOD酶活性的强弱能直接反映出植物在低温胁迫下的自我调节能力,常被作为评价植物抗寒性的重要指标[7,24]。低温胁迫下柳杉各无性系SOD酶活性整体上表现出先短暂的下降后逐渐上升而后又下降的趋势。低温初期SOD活性的降低与陈政等[19]对日本无刺花椒的研究结果一致。章锦涛等[25]在研究山茶的抗寒能力时发现随着胁迫温度的降低,植株体内SOD活性先升高后降低。在低温胁迫下为了维持植物体内活性氧和自由基的动态平衡,植物体内的保护酶体系会发挥作用清除这些有害物质,使SOD等保护酶的活性增强,但超过保护酶清除作用范围时其酶活性会下降[26-27]。郭祥泉[28]认为随着低温胁迫时温度的降低,SOD酶活性有一个温度拐点,在拐点前其活性急剧上升,在拐点后又下降,以此来定量研究不同树种及不同个体的低温临界值。
光合作用是植物对逆境较为敏感的过程,低温胁迫会抑制PS II的活性,使其光能捕捉能力与光化学转化效率降低,反应中心的开放程度降低,从而引起叶绿素荧光参数的变化[29-30]。植物可变荧光Fv与最大荧光Fm的比值Fv/Fm能够良好地反映光合色素把捕获的光能转化为化学能的速度和效率[31-32]。Fv/Fm下降越多表明PS II受损伤越大,Fv/Fm的变化可反映低温下PS II受伤害的程度[33]。柳杉各无性系Fv/Fm值在4℃-0℃出现短暂的上升,0℃后随着低温胁迫的加剧而逐渐降低。Fv/Fm的降低说明PS II反应中心受到伤害,抑制了光合作用的原初反应,不利于光能的转换。Y(II)值代表光系统 II的实际量子产量,它反映了植物的实际光合效率[29]。qP是衡量开放的光系统 II反应中心光抑制的程度,一定程度上表示PS II电子传递活性[34]。高qP值可以加速推动光合电子传递能力,促进光合效率增长[35]。Y(II)的变化趋势与Fv/Fm的变化大致相同,随着温度的降低柳杉各无性系Y(II)值呈现先升高再降低的趋势,而qP在低温初期变化不明显,0℃后与Y(II)值变化相同,随着温度的变化逐渐降低,甚至趋近于0,说明PS II反应中心的开放受到影响,实际光合效率和光合活性均受到低温的抑制。Fv/Fm、Y(II)和qP值在4℃-0℃的回升及不变说明柳杉在经历低温初期有抵御能力,能够在一定时间内维持光合效率,当超过临界值其光合效率会大幅降低,严重时导致反应中心受到损伤甚至失活。
植物是一个统一的有机整体,逆境胁迫下引起植物生理生化指标变化的因素错综复杂,因此,单一指标很难准确评价植物的抗逆性[36-37]。主成分分析能有效地简化数据,分析各指标间的关系,起到了浓缩数据的作用[14,38]。利用主成分分析及隶属函数法,可以综合评价柳杉各无性系的抗寒能力,在一定程度上可以真实的反映其抗寒能力强弱[39-40]。陈新华[41]运用综合评价的方法,对不同品种甜樱桃的抗寒性进行了评价,苏李维等[14]用主成分分析法结合隶属函数值建立了葡萄抗寒性综合评价方法。本研究以相对电导率(REC)、MDA含量、SOD活性及叶绿素荧光参数(Fv/Fm)为评级指标,依据权重与隶属函数计算柳杉各无性系抗寒性综合指数,最终结果表明57#、68#和32#为抗寒性较强的无性系,3#、25#和42#无性系抗寒性较弱。
4 结论
低温胁迫下,测定10个柳杉无性系针叶的丙二醛含量、超氧化物歧化酶活性及叶绿素荧光参数,其变化均具有显著差异。结合主成分分析与隶属函数法对10个柳杉无性系的抗寒性进行综合评价,其中57#、68#和32#无性系对低温胁迫具有较强的适应性,抗寒能力强,而3#、25#和42#无性系抗寒性较弱。
文章所有附图附表数据请到本刊官网下载(http://biotech.aiijournal.com/CN/1002-5464/home.shtml)。
