甜瓜果实长度主基因+多基因遗传分析
2022-05-07李文龙梁晓雪胡建斌
黄 松,李文龙,梁晓雪,李 琼,胡建斌
(1.信阳农林学院 河南 信阳 464001; 2.河南农业大学园艺学院 郑州 450002)
甜瓜(L.)为葫芦科一年生蔓生植物,其多样性仅次于同科植物南瓜。近期进化研究表明,栽培甜瓜历史上经历了三次独立驯化,第一次发生在非洲,随后的两次发生在亚洲,整个亚洲及地中海沿岸的北非和南欧等广大地区都有可能是栽培甜瓜的驯化地。甜瓜丰富的多样性主要表现在果实性状上,其变异类型众多,包括果实形状、果皮底色、果面覆盖物(网纹、覆纹、果皮毛)、果面特征(沟、棱、皱纹)、果肉厚度、果肉颜色、果肉品质(糖、酸、维生素C 含量及香气和质地等)等性状的变异。据此,Pitrat建议将栽培甜瓜分为长毛亚种(ssp.)和短毛亚种(ssp.)及其下属的16个变种。其中,长毛亚种和短毛亚种分别对应生产上的厚皮甜瓜类型和薄皮甜瓜类型。果实形状(或果形指数)是甜瓜重要的外观性状,也是甜瓜变异最明显的性状之一,主要有圆形、苹果形、卵形、椭圆形、圆柱形、棒形等6 种类型。生产上推广应用的厚皮甜瓜品种多以圆形和椭圆形为主,薄皮甜瓜品种以苹果形和卵形为主。相关性研究表明,决定果实形状的主要因素是细胞的纵向分裂速率,即果实长度。因此,探明甜瓜果实长度的遗传基础,对果实外观性状的遗传改良具有重要意义。
早期Wall认为,甜瓜长果形对圆果形是显性遗传。林碧英等对薄皮×厚皮甜瓜杂交后代进行分析发现,长果形或椭圆形对圆球形均为显性遗传。然而更多的研究表明,甜瓜果实长度是典型的数量性状,无论是在自然群体还是在特定分离群体中,果实长度都会出现连续变异。目前,在甜瓜的12 条染色体上均发现有控制果实长度或果形指数的QTL,大约有130 个,但在不同的遗传背景下,其定位结果不尽相同。其中,报道的第8 染色体QTL最多,达26 个,而第5 染色体仅有2 个QTL。这些研究结果表明,调控甜瓜果实长度的基因遍布整个基因组,且其效应大小不一,遗传基础十分复杂。主基因+多基因混合遗传模型是解析复杂数量性状遗传基础的有效方法,该方法通过分析特定群体的性状表型,以检测影响目的性状的主基因和多基因数目,并评估其遗传效应,进而揭示目的性状的遗传规律,已经用于茄子、西葫芦、黄瓜等果实形状的遗传分析。
笔者以果实长度差异明显的2 份甜瓜材料构建6 个世代群体,采用主基因+多基因混合遗传模型分析方法,对春秋两季甜瓜果实长度进行遗传分析,以期明确果实长度所属的遗传模型及其基因效应,为甜瓜果实形状遗传改良提供理论依据。
1 材料与方法
1.1 材料
试验材料为圆果形自交系H581(P)和长果形H906(P),其果形指数分别为1.0 和12.0,花期子房长度差异也十分明显(图1),两份材料均选自前期构建的甜瓜核心种质。2017 年秋季杂交配制F,2018 年春季自交获得F,秋季回交获得回交1 代B(F×P)和B(F×P)。为了准确统计分离群体单株的果实表型,将B、B和F群体各单株分别自交构建其家系(B、B和F)。

图1 亲本材料H906 和H581 的果实和开花当天的子房表型
1.2 方法
2019 年春季(3—7 月)和秋季(7—10 月)在河南农业大学毛庄科教园区试验基地进行甜瓜各世代群体的育苗和田间种植。春季于日光温室中育苗,秋季于塑料大棚中遮阳育苗。幼苗长至2 叶1心时定植,非分离群体P、P和F分别种植30 株,分离群体B、B和F分别种植96 个、110 个和140 个家系,每个家系包括7~10 个单株。每株在11~15 节选留1 个瓜,除去其他雌花及侧枝,其他田间管理同常规。待果实成熟时采用直尺(最小刻度1 mm)测量果实长度。
1.3 数据分析
非分离群体(P、P和F)各单株的果长实测值为其表型值,分离群体(B、B和F)各单株的果实表型值以其对应的家系单株实测值的平均值代替。

2 结果与分析
2.1 果实长度的变异分析
春秋两季对甜瓜6 个世代群体的果实长度进行统计,结果见表1。春季非分离群体P、P和F果实长度均值分别为10.2、110.9、43.4 cm,秋季果实长度均值分别为和9.7、98.8、39.4 cm,且群体内变异较小(<17%)。春季果实长度平均值普遍高于秋季,可能是春季果实生长发育时期比秋季长的缘故。F均值处于P和P之间,但无论是在春季还是秋季,其均值明显偏向圆果亲本P。分离群体(B、B和F)的果实长度变异较大,春季>25%,秋季>40%,秋季果实长度变异更明显。无论是春季还是秋季,B果实长度均值最小,B果实长度均值最大,F果实长度均值介于二者之间。各分离群体的峰度均为负值,说明存在主效基因的作用。果实长度在3 个分离群体中呈现连续分布,春秋两季B和F果实长度偏向圆果亲本P,B偏向长果亲本P2,且均呈现单偏峰分布(B和F偏度均为正值,B偏度为负值)(图2),符合主基因+多基因表型变异的基本特征。

图2 甜瓜果实长度在分离群体中的频次分布

表1 甜瓜果实长度表型统计
2.2 遗传模型的选择和检验
采用主基因+多基因混合遗传模型对6 个世代群体果实长度表型值进行遗传分析,分别计算24种模型MLV 值和AIC 值,结果见表2。根据AIC值最小原则,选取最小AIC 值及与之差异不大的遗传模型作为备选模型。春季3 个最小AIC 值分别为2 188.243、2 192.339 和2 184.243,所对应的模型分别为C-0、D-0 和E-1,秋季3 个最小AIC 值分别为867.288、859.539 和860.129,所对应的模型也是C-0、D-0 和E-1,即这3 种模型为春秋两季所共有,均可作为果实长度的备选模型。C-0 对应PG-ADI,为加性-显性-上位多基因模型,无主基因效应;D-0和E-1 分别对应MX1-AD-ADI 和MX2-ADI-AD,受1 对或2 对主效基因控制,也受多种效应的多基因影响。由此可见,备选的3 种遗传模型存在一定的差异,特别是C-0 与D-0、E-1 的差异较大,需要进一步检验,以确定最适遗传模型。
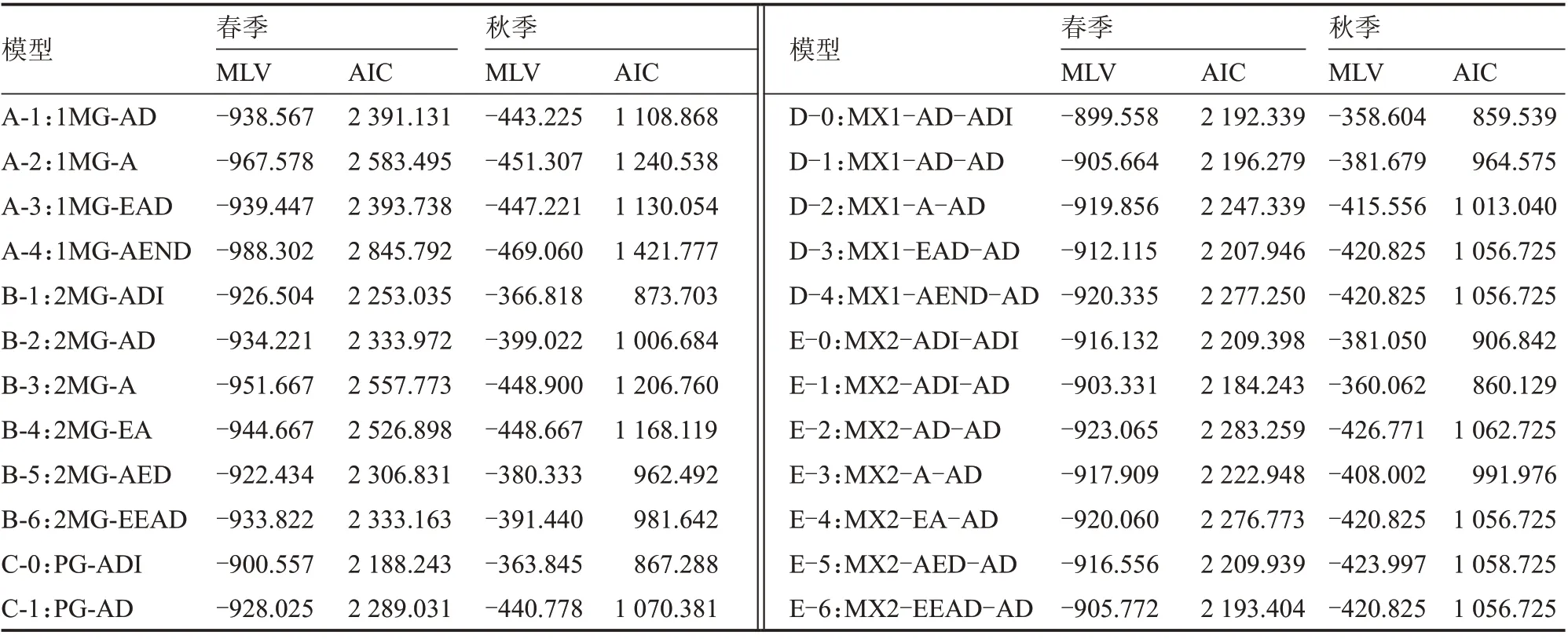
表2 各遗传模型的MLV 及AIC 值
对3 个备选模型进行适合性检验,结果见表3。C-0 模型在春秋两季共出现7 个显著性统计量,D-0模型在两季共有3 个显著性统计量,而E-1 模型在两季未出现显著性统计量。根据显著性统计量数最少原则,E-1 模型可认为是最适遗传模型,即2 对加性-显性-上位性主基因+加性-显性多基因模型。因此,甜瓜果实长度由主基因和多基因联合调控,主基因的遗传效应包括加性、显性和上位性效应。

表3 备选模型C-0、D-0 和E-1 的适合性检验
2.3 E-1模型遗传参数的估测
基于最适遗传模型E-1,各类参数估算结果见表4。一阶参数估算表明,控制果实长度的主基因有2 对(、),其加性效应()在春秋两季均为正值,但效应值的差异较大。第1 对主基因()的效应值较大(春季12.22,秋季9.74),第2 对主基因()的效应值相对较小(春季5.06,秋季1.69),均能增加果实的长度。两对主基因的显性作用()均为负向,能减少果实长度,但效应值差异明显。基因春季显性效应最强(-12.08),秋季则较弱(-3.67),基因在春秋两季的显性效应不同,但效应值均较低(||<2),说明显性效应受环境影响较大。由此可知,基因的加性和显性效应明显强于基因,且在春季的作用更为明显,说明基因在春季能更好地发挥其育种价值。

表4 最适模型E-1 的一阶和二阶参数估算
注::群体均方;、:第一、二主基因的加性效应;、:第一、二主基因的显性效应;:两主基因间的加性×加性互作效应;:两主基因之间的加性×显性互作效应;:两主基因之间的显性×加性互作效应;:两主基因之间的显性×显性互作效应;:多基因方差;:主基因方差;:主基因遗传率;:多基因遗传率。
2 对主基因存在多种互作效应,其中加性×加性互作效应()较强,秋季(14.11)高于春季(5.98),可用于增加果实长度。尽管2 对主基因的加性-显性互作效应(和)和显性-显性互作效应()均能被检测到,但其效应值均很低(||、||和||均小于5),且不同季节的作用方向不同,说明互作效应对表型的贡献甚微,且易受环境影响,难以用于果实长度的遗传改良。
二阶参数的估算结果表明,春季主基因在B、B和F群体中的遗传率()分别为42.67%、69.59%和81.05%,秋季3 个群体的分别为61.03%、70.42%和83.33%,主基因在B和F中表现出高遗传率,说明B和F世代有利于果实长度的选择与改良。多基因遗传率()相对较低,且仅在部分群体中被检出。如春季B的最大,也仅为26.57%,其他群体很低(<14%),秋季B和春秋两季F的h均为0。由此可见,甜瓜果实长度主要受主基因控制,主基因遗传率在各世代的比重较高,早期选择能取得较好的效果。
3 讨论与结论
果实长度是决定果实形状的重要因素,也是果实重要的外观性状。作为遗传多样性最丰富的葫芦科植物之一,甜瓜果实形状变化多样,不同果形的商业品种也十分常见,如长果形的羊角蜜,椭圆形的西州密25,圆果形的玉菇。因此,果实长度也是甜瓜重要的商品性状。甜瓜果实长度除了受遗传因素调控外,还受光照强度、花期温度、土壤营养状况等环境因素影响。尽管前人已经对甜瓜果实形状(果实长度、果形指数等)进行了大量遗传研究,如通过配制各种作图群体(BC、F、RIL 等),并在可控的条件(如温室、大棚等)下观测果实表型性状以减少环境影响,但至今仍然未克隆到调控甜瓜果长的关键基因,这从侧面反映了甜瓜果实长度遗传基础的复杂性。
作为剖析复杂数量性状遗传结构的一种有效方法,主基因+多基因混合遗传模型已经应用于果实形状的遗传基础分析。如甜瓜同属植物黄瓜,其果实长度符合2 对加性-显主基因模型(B-2),主基因遗传率最高达88.09%。茄子和辣椒的果实长度均符合2 对加性-显性-上位性主基因+加性-显性上位性多基因模型(E-1),主基因遗传率较高(>50%)。西葫芦果实长度受1 对加性主基因+加性-显性多基因(D-2),但主基因遗传率较低(<30%)。不同种类植物的果实长度遗传模型不尽一致,但大多受少数主基因和多基因联合调控,为挖掘果实长度调控基因提供了理论依据。笔者从前期构建的甜瓜核心种质中鉴定了一份果实极长的种质材料H906,该种质为甜瓜长毛亚种(ssp.)下的蛇瓜变种(var.),是目前所知的甜瓜中果实最长的类型,可能包含了控制果长的关键基因。笔者通过主基因+多基因混合遗传模型分析发现,甜瓜果实长度符合2 对加性-显性-上位性主基因+加性-显性多基因模型(E-1),受主基因和多基因的联合调控,且包含多种遗传效应,与前人研究结果基本一致。
在本试验中,E-1 模型中主基因的遗传效应显得更为复杂。其中,加性和显性效应对果实长度起主要调控作用,尤其是春季的作用更明显,且以第1对主基因()为主,互作效应影响较小(除加性互作效应外)。由于加性和显性效应在育种实践中有较大的利用价值,因此甜瓜果实长度的遗传改良可从关键的基因入手,在特定季节(如春季)加强对基因的选择。由于主基因的遗传率较高(除春季B外均高于60%),多基因遗传率较低(除春季B外均低于14%),因此,甜瓜果实长度的遗传改良可考虑在F、B及其自交后代中开展,在早期世代加强对主基因(特别是基因)的选择,以便快速获得所需的单株。同时,考虑到主基因的加性和显性效应均发挥重要作用,而显性效应是不可固定的遗传变量(随世代推移可能会消失),遗传操控难度较大,因而在实际选择育种过程中,应尽量扩大群体规模,选择同时具备加性和显性效应的基因型单株,从而获得理想的育种材料。
尽管本试验探明了甜瓜果实长度所属的遗传模型,证实了果实长度具备遗传改良的潜力,但仅仅通过传统的田间性状重组来改良果实长度不可避免的存在盲目性。因此,在明确其遗传方式的基础上,借助现代高通量基因分型技术,采用正向遗传学方法鉴定控制果实长度的主效基因或QTL,通过分子标记辅助选择可实现果实长度的精准改良。
