泡核桃凋落叶对生菜光合特性的影响
2022-04-25郑崇兰李志超祝铭谦巫玲琳刘永安
郑崇兰,龙 丽,黄 庆,李 培,李志超,祝铭谦,巫玲琳,刘永安*
(1.凉山彝族自治州林业草原科学研究院,四川西昌 615000;2.凉山彝族自治州张林国有林场,四川西昌 615000)
植物化感作用是指植物通过向外界环境中释放有效的化合物,对自身和相邻的其他植物(包括微生物)产生的促进或抑制作用[1]。泡核桃(Juglans sigillataDode)广泛分布于我国西南地区,是分布区重要的果用、油料、材用和绿化的经济林树种,被许多地区列入经济建设的主导产业进行重点开发。开展泡核桃林下种植可充分利用土地资源,但已有研究表明核桃叶片中含有大量的醌类、黄酮类、二芳基庚烷类、萜类、多酚类等化感物质[2-3],林下凋落叶势必对林地环境及林下植物产生影响。研究发现,核桃凋落叶在土壤中分解,显著抑制萝卜、白菜、芥菜叶片中的叶绿素合成、降低三种作物的光合能力[4],且抑制作用随着凋落叶添加量的增加而增强[5]。而将核桃凋落叶化感作用耐受性强的植物用于开展林下套作,能有效提高土地利用率[5]。有研究表明生菜对麻风树叶子、种子壳的水浸提液[6]及橡胶叶水浸提液[7]的化感作用较不敏感,受银杏凋落叶[8]和银杏外种皮[9]的化感抑制作用却很显著。
本研究以生菜(Spinaciao leracea)为受体植物,采用盆栽试验,向土壤中添加不同量的泡核桃凋落叶,模拟泡核桃凋落叶自然分解,探讨泡核桃凋落叶对生菜光合作用的影响,为泡核桃林下种植提供理论依据。
1 材料与方法
1.1 试验材料
采用四川省凉山州木里县15 年生泡核桃人工林中采集的新鲜凋落叶,粉碎后,混合均匀,备用。
选择大小均匀、饱满的生菜种子,播种前用0.5%K2MnO4溶液消毒20 min,用蒸溜水冲洗5 次,备用。
栽种土壤选择芬兰进口泥炭土,基本理化性质见表1。栽种器具选用上口径24 cm、下口径17 cm、高14 cm的聚乙烯塑料盆。
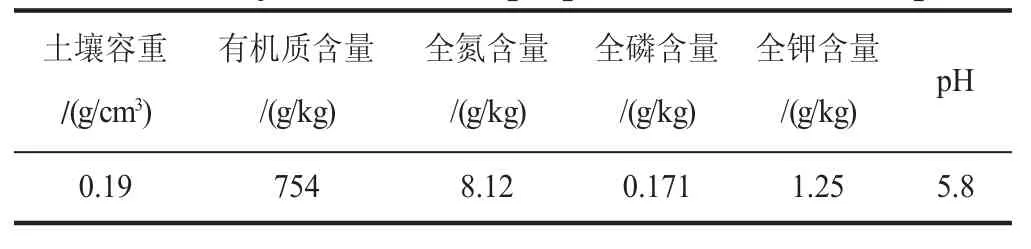
表1 供试土壤基本理化性质Table 1 Physico-chemical properties of the soil samples
1.2 试验设计
本试验于2021 年5—9 月在凉山州林业草原科学研究院塑料大棚内进行。试验采取单因素随机试验设计,依据调查数据,人工泡核桃林每年平均凋落叶量约为8 000 kg/hm2,按盆口径22 cm 折合量为30 g/盆,故本试验设置凋落叶添加量分别为T15(15 g/ 盆,约4 000 kg/hm2)、T30(30 g/ 盆,约8 000 kg/hm2)以 及T60(60 g/盆,约16 000 kg/hm2)3 个处理组,以不添加凋落叶为对照组。每个组设置10 个重复,共计40 盆。
按处理要求,将定量的凋落叶与试验用土充分混匀后浇透水,1 d 后播种生菜。播种时覆盖0.5 cm 试验用土,浇透水以便种子发芽。各处理全部出苗后10 d 进行间苗,然后实施常规田间管理。
1.3 指标测定及方法
叶片光合生理指标采用便携式光合作用仪(ADClci-t,英国)测定,光合仪选择开放气路,人工设置光照强度为1 000 μmol/(m2·s),选取茎中部完全成熟的叶片测定叶片净光合速率(net photosynthetic rate,Pn)、蒸腾速 率(transpiration rate,Tr)、气孔导度(stomatal conductance,Gs)和胞间CO2浓度(intercellular CO2concentration,Ci),每个处理重复测定3 次,每次重复记录5 个数据。日进程于8:00—18:00 进行测定,光合生理特征参数在上午9:00—11:00 进行测定。
1.4 计算方法
化感作用效应敏感指数简称化感指数(RI),计算公式如式(1)(2)所示。

式中,C为对照CK 光合参数测定值,T为处理T15、T30、T60 光合参数测定值。
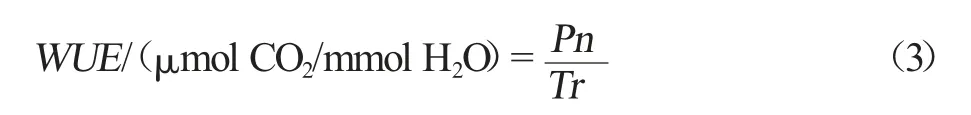
叶片水分利用效率(WUE)计算公式如式(3)所示。式中,Pn 为叶片净光合速率,μmol CO2/(m2·s);Tr为叶片蒸腾速率,mmolH2O/(m2·s)。
1.5 数据处理
采用Excel 2007 进行数据整理,SPSS 19.0 进行单因素方差分析,LSD 法进行多重比较。
2 结果与分析
2.1 泡核桃凋落叶对生菜光合特征参数的影响
由表2 可以看出,T15 处理的生菜净光合速率最大,然后依次为CK、T30,最小为T60,泡核桃凋落叶对生菜净光合速率的影响总体表现为低量促进、高量抑制。低量处理T15 的净光合速率为13.08 μmol CO2/(m2·s),显著高于CK,且其化感指数为0.081,表现为促进效应。高量处理T30、T60 的净光合速率分别为6.32 μmol CO2/(m2·s)和4.65 μmol CO2/(m2·s),化感指数为-0.474 和-0.613,表现为抑制效应,且各处理间净光合速率测定值的差异显著(P<0.05)。由此可见,每盆土添加15 g 泡核桃凋落叶,可显著促进生菜的净光合速率,添加量超过30 g 之后,变为抑制作用。
由表2 可知,胞间CO2浓度表现为随泡核桃叶添加量的增加逐渐增加,CK 最低为271.59 mmol/mol,T15 较CK 稍高,为272.69 mmol/mol,但与CK 无显著差异,化感指数仅为0.004。处理T30 与T60 的胞间CO2浓度显著高于CK的,分别为 308.37 mmol/mol 与 332.16 mmol/mol。化感指数达到了0.119 和0.182,表现为促进作用。这表明,每盆土添加量15 g 泡核桃凋落叶对生菜胞间CO2浓度影响不大,添加量达30 g 之后,胞间CO2浓度表现出上升趋势。
气孔导度变化与净光合速率类似,表现出低促高抑的趋势,T15 气孔导度为0.21 mol/(m2·s),显著高于对照CK,化感指数为0.098,表现为促进作用。当每盆泡核桃凋落叶添加量达到30 g 时,其对生菜叶片气孔导度的影响变为抑制,T30、T60 气孔导度下降为0.11 mol/(m2·s)和0.12 mol/(m2·s),化感指数为-0.401 和-0.350,T30 和T60较CK 相比显著下降,但处理间无较大差异。
泡核桃凋落叶对生菜片蒸腾速率的影响结果见表2。由表知,低量处理T15 的蒸腾速率为4.90 mmol/(m2·s),显著高于对照,化感指数为0.147,表现为促进作用。高量处理(T30、T60)降低了生菜叶片蒸腾速率,化感指数为-0.097 和-0.033,表现为抑制效应。
由表2 可以看出,土壤中添加泡核桃凋落叶降低了生菜叶片的水分利用效率,且降低程度随添加量的增加而增加,且各处理间差异显著(P<0.05),化感指数达到了-0.072、-0.417 和-0.595。

表2 不同泡核桃叶添加量处理下生菜的净光合速率及气体交换参数Table 2 Net photosynthetic rate and gas exchange paramaters of lettuce under different amount of J. sigillate leaf litter
2.2 泡核桃凋落叶对生菜光合特征参数日变化的影响
由图1A(见下页)可看出,CK、T15、T30 和T60 四种处理净光合速率日变化均呈现“双峰型”,在10:00 时和14:00 分别达到峰值,有明显的“光合午休”现象。4 个处理均在10:00 时达到全天的最高峰,T15 最大,为11.63 μmol/(m2·s),之后依次为CK、T30 和T60,且各处理间有显著差异(P<0.05)。在14:00 时达到第二次高峰,各处理均低于第一次高峰,其峰值以CK 为最高,净光合速率达到8.01 μmol/(m2·s),与T15(净光合速率为7.99 μmol/(m2·s)无显著差异,均显著高于T30 和T60。
由图1B(见下页)可知,CK、T30 和T60 处理下生菜苗气孔导度均呈现“双峰型”,在10:00 出现第一个峰值,14:00 出现第二个峰值。CK 处理的最大峰值出现在下午16:00,达到0.188 mol/(m2·s),而T30 和T60 峰值出现在12:00,分别为0.128 mol/(m2·s)和0.154 mol/(m2·s)。T15 的气孔导度在14:00 以前未出现明显的峰值,最大值为0.148 mol/(m2·s),但各时间(除CK 16:00 外)均显著高于其它处理。
由图1C 可知,四个处理的胞间CO2浓度均呈现“W型”,在早上8:00 的胞间CO2浓度较高,然后下降,在中午12:00 出现第二个峰值,随后下降,在下午14:00 出现第二个低谷,随后逐渐上升,在下午18:00 达到全天的峰值。胞间CO2浓度大小依次为T60>T30>CK>T15。
由图1D 可看出,不同泡核桃叶添加量处理的生菜蒸腾速率日变化均呈现为“单峰型”,各处理均从8:00 开始上升,在中午12:00 达到峰值,随后逐渐下降。T15、CK、T60 和T30 各处理的峰值大小分别达到8.809、7.211、6.450、5.971 mmol/(m2·s)。
由图1E 可知,不同泡核桃凋落叶处理生菜的水分利用效率的日变化,总体呈现下降趋势,在中午12:00 时出现一个低谷,在下午14:00 时出现一个小高峰,其峰值远低于早上8:00,然后开始下降。各处理的全天平均水分利用效率由大到小依次为CK、T15、T30 和T60,分别为1.095、1.075、0.909、0.655 μmol CO2/mmol H2O。
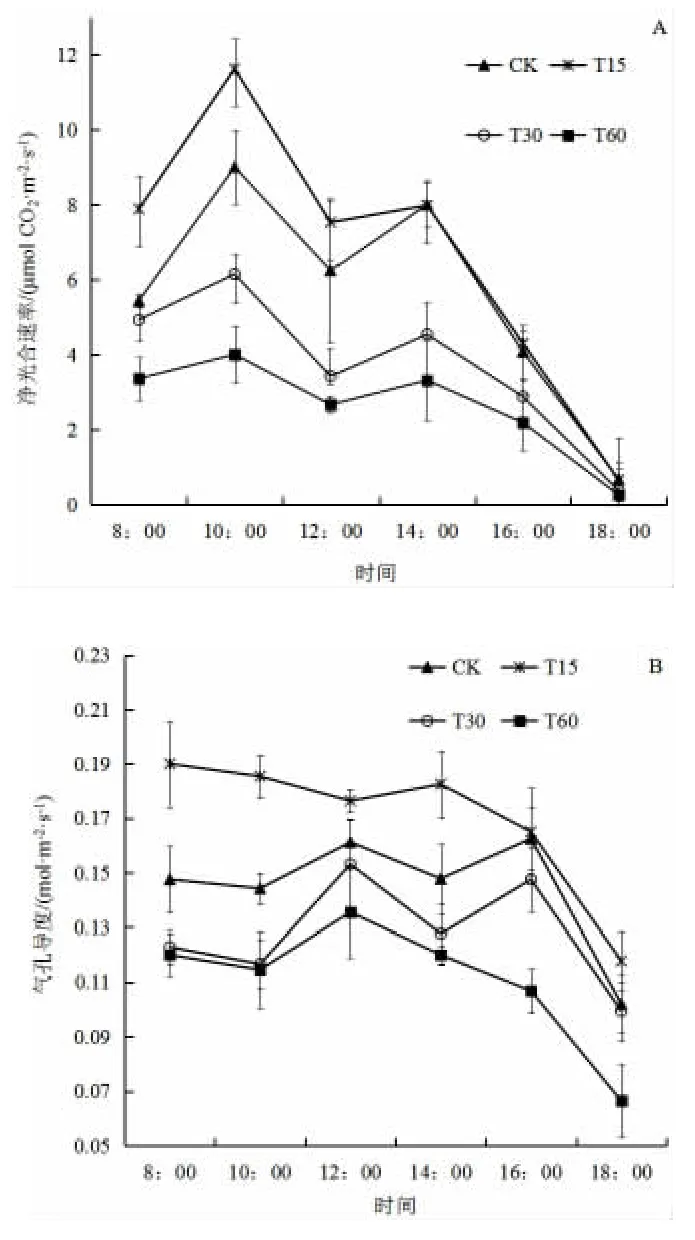
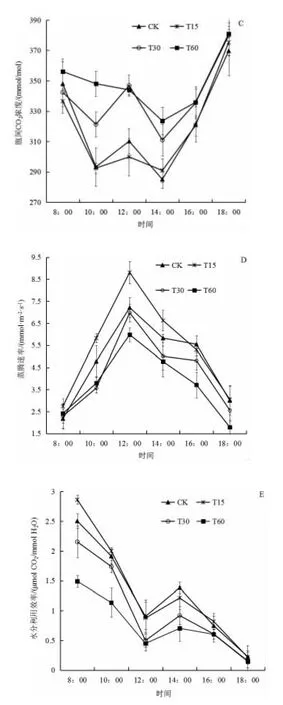
图1 不同泡核桃凋落叶添加量处理下生菜叶片气体交换参数日变化Fig.1 Diurnal variation characteristics of gas exchange parameters of lettuce under different amount of J. sigillate leaf litter
3 讨论与结论
本试验中,生菜的净光合速率随凋落叶添加量增加先升高后降低,胞间CO2浓度随凋落叶添加量的增加而增加,且均高于对照,而气孔导度则表现为先升高后降低。植物叶片光合效率降低有气孔部分关闭导致的气孔限制和叶肉细胞光合活性下降导致的非气孔限制两类[10]。Farquhar 等[11]认为,如果净光合速率的降低伴随着胞间CO2浓度的降低,则净光合速率的下降主要是受部分气孔关闭的气孔限制所致;反之,如果净光合速率下降伴随着胞间CO2浓度升高或不变,则净光合速率的下降主要是由叶肉细胞光合活性下降导致的非气孔限制引起的。初步可以推断,本试验中生菜净光合速率的下降是非气孔限制导所致。低量添加泡核桃凋落叶提高了生菜的净光合速率,而土壤中泡核桃叶凋落叶增多时其净光合速率降低,且显著低于CK,说明添加一定量的泡核桃凋落叶能提高生菜的光合效率,可能是因为低量的凋落叶分解产生的化感物质较少,不足以对生菜生长产生影响,反而通过凋落叶分解产生的营养物质促进了生菜的生长;超过一定量后,过多凋落叶分解产生的化感物质积累,破坏了生菜叶片光合器官的结构,从而导致净光合速率显著下降。
另外,与对照相比,不同处理生菜水分利用效率均显著下降,T15 可能是因为化感物质使生菜叶片的气孔导度增加,从而提高了蒸腾速率;而高添加量处理T30、T60 生菜的水分利用效率下降,可能是因为高浓度的化感物质抑制了生菜的光合作用。光合作用是植物能量的来源,也是植物生长、发育及代谢的物质基础。环境因子的改变会影响植物的光合特性[12],化感物质对植物体光合作用的影响主要表现在使叶绿素含量和光合速率的降低上[13-15]。曾有研究表明,核桃凋落叶的化感物质能显著地降低小麦、燕麦、黑麦草和莴笋等植物的叶绿素含量[16-17]。化感物质可以通过影响气孔导度、胞间CO2浓度、蒸腾速率等气孔因素直接影响植物的光合作用[18]。
本试验中,低浓度泡核桃凋落叶处理的生菜净光合速率在上午12:00 以前显著高于对照,在下午16:00 其净光合速率从“午休”后恢复较慢,其净光合速率和气孔导度提高较小,胞间CO2浓度降低较小,由此暗示经泡核桃凋落叶处理后,因泡核桃叶中的化感物质影响了气孔开张,从而影响了生菜的光合效率和水分利用效率。马红叶等[19]研究也表明化感物质可通过降低叶片中的叶绿素含量、影响蒸腾速率和气孔导度等方式来降低植物的光合速率。
综上,泡核桃凋落叶中的化感物质对生菜植物的光合生理产生显著的影响,盆土中泡核桃凋落叶添加量为15 g,即约4 000 kg/hm2时,能促进生菜光合作用;盆土中泡核桃凋落叶添加量达到30 g,即约8 000 kg/hm2时,能提高胞间CO2浓度,降低气孔导度和蒸腾速率和水分利用效率,从而降低植物吸收、转化和利用光能的能力,导致光合速率的降低,影响植物对碳的固定和同化,从而抑制植物的光合效率。试验结果表明,在去除部分凋落物之后,可在泡核桃林下套种生菜,以提高土地利用率,但具体栽培方式需进行大田试验进一步确定。
