贺兰山低山区土壤种子库空间分布特征
2022-04-06李国旗王雅芳谢博勋
李国旗,王雅芳,刘 星,谢博勋,解 盛,石 云
(1 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,银川 750021;2 宁夏大学西北退化生态系统恢复与重建教育部重点实验室,银川 750021)
贺兰山地处中国西北干旱区,具有重要的地理意义,其作为最后一道生态屏障,阻隔了毛乌素沙地、乌兰布和沙漠、腾格里沙漠这三大沙漠进一步向银川平原扩张,拦截了源自蒙古国、中国阿拉善地区等的沙尘暴;贺兰山的生态安全不仅影响其周边的黄河段、银川平原,还可能进一步影响西北地区、华北地区甚至全国的生态安全[1-2]。贺兰山地处华北平原、蒙古高原、青藏高原的交汇处,其地理位置决定了该地区生物类型复杂多样,且具有交叉性和过渡性[3-4];贺兰山位于温带草原与荒漠的交汇处,山体高耸,生物类型具有明显的空间分异规律[5-6]。植物优势种主要有短花针茅(Stipabreviflora)、蒙古扁桃(Amygdalusmongolica)、沙蒿(Artemisiadesertorum)、青海云杉(Piceacrassifolia)等[7-8]。
土壤种子库是存在于土壤表面和土壤中有生命活力的种子的总量[9-10]。也可以定义为单位面积土壤中的全部有生命活力的种子[11-13]。土壤种子库可作为植物的潜在种群,影响植被自然更新和恢复、生物多样性保护、植被的演替进程和方向[14-16]。大量研究表明,环境空间异质性是制约或影响植物种群、群落乃至生态系统和物种多样性等的重要因素,其中地形是主要来源[17-18]。地形通过其单一的生态作用或受一定的地形复合体影响地表的光、热、水、土、肥等生态因子,从而改变能量的平衡,直接或间接地影响植被格局。而土壤种子库中的种子主要来源于地上植被,地形可通过影响地上植被进一步影响土壤种子库[19-20]。
对贺兰山在不同海拔高度上开展土壤种子库调查研究,主要目的是:① 探明土壤种子库特征的空间差异性;② 比较研究各海拔土壤种子库与相应地上植被的物种组成关系的差异性;③ 评估土壤种子库对植被自然恢复与更新的贡献潜力在海拔间的差异性。贺兰山低山区风蚀严重、植被稀疏,植被自然恢复潜力尚未可知,对不同海拔高度种子库开展研究,探讨地上植被自然恢复潜力在海拔间的差异性,以期为贺兰山低山区植被合理利用、恢复治理与生态重建提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于宁夏回族自治区贺兰山国家自然保护区,地理位置 105°49′~106°41′E,38°19′~39°22′N,东接银川平原,西侧和北侧与阿拉善戈壁荒漠相接,位于温带草原与荒漠的交汇处[1-2]。贺兰山地处中国西北中温带气候区,为典型温带大陆性气候并具有山地气候特征。无霜期229 d,春季风大、沙多,气候变化大,年均温-0.8 ℃。年均降水量420 mm,年均蒸发量2 000 mm,降雨集中在7~9月份,占全年降水量的70%~80%[20]。低山区气温变化剧烈,干旱少雨,为显著大陆性气候。山地植被和土壤具有明显的空间分异规律,海拔由低到高,植被可划分为山前荒漠与荒漠草原带(海拔1 600 m以下)、山麓与低山草原带(海拔1 600~1 800 m)、中山和亚高山针叶林带(海拔1 800~3 100 m)和高山亚高山灌丛、草甸带(海拔3 100 m以上)4个垂直带,土壤划分为棕钙土带——灰褐土带——高山亚高山灌丛、草甸土带这3个带[3-4]。
1.2 样地设计与样品采集
从海拔1 200到2 000 m,每隔200 m设置1个取样点,选取群落生境相对均匀的地方,设置3个5 m×5 m的大样方,进行灌木调查;在每个大样方内沿对角线布置3个1 m×1 m的小样方进行草本调查。调查记录每个样方内各植物物种名称、盖度、高度、多度等,其中盖度采用方格网法直接计算;高度用测高尺测量,做3~5个重复;多度完全记录小样方内每种植物。在植被调查的同时在每个小样方中采用五点取样法采集土样(取样器采用20 cm×20 cm×10 cm规格)。
1.3 实验方法
采用常规分析法测定土壤理化性质(表1)。土壤种子库采用室内萌发鉴定的方法。将土样均匀的平铺在萌发盆里,厚度约为2 cm,盆底还铺有5 cm的无籽毛细沙。将处理好的萌发盆置温室中,采取少浇、勤浇的方式,保持土壤湿润,等待种子萌发。幼苗种属鉴定后去除,未鉴定出的继续生长,直至可以鉴定出,连续4周没有新幼苗萌发结束实验。土壤种子库中种子密度用单位面积(1 m2)土壤中有效种子数量来表示。

表1 土壤理化性质的测定方法
1.4 数据统计与分析
1.4.1 地上植被的物种重要值计算根据对5个海拔地上植被群落特征的调查研究,选取相对多度、相对盖度、相对频度及相对高度4个因素来计算植物各物种重要值,计算公式为:
重要值(IV)=(相对多度+相对盖度+相对频度+相对高度)/4
相对多度=(某一种多度/所有种多度之和)×100
相对盖度=(某一种盖度/所有种盖度之和)×100
相对频度=(某一种频度/所有种频度之和)×100
相对高度=(某一种的平均高度/所有种的平均高度之和)×100
1.4.2 物种多样性指标计算Margalef 丰富度指数:
R=(S-1)/lnN;
Shannon-wiener 指数:
H′=∑PilnPi;
Simpson 优势度指数:
Pielou 均匀度指数:
E=H′/lnS
式中:S为物种总数;Pi=Ni/N,N为样方中总的个体数,Ni为样方中第i物种的个体数。
1.4.3 相似性指数计算不同海拔高度土壤种子库与地上植被的相似性、海拔间土壤种子库的相似性以及地上植被的相似性,均采用Sorensen的相似性系数(similarity coefficient,SC)来表示,计算公式为:
式中SC为相似性系数;w代表土壤种子库和地上植被共同拥有的物种数;a代表土壤种子库中物种数;b代表地上植被物种数。
SC值在0~0.24为极不相似;0.25~0.49为中等不相似;0.50~0.74为中等相似;0.75~1.00为极相似。
1.5 数据统计与分析
野外调查及室内试验所得实验数据在Excel表格中进行初步处理,在SPSS 21.0软件中进行多重比较和相关性分析,在Canoco4.5进行RDA分析。不同海拔间土壤各性质、种子库特征及地上植被特征均采用单因素方差分析和LSD法进行多重比较,用Person相关系数法进行相关性分析,土壤种子库物种多样性与土壤因子间的关系进行RDA分析与制图。
2 结果与分析
2.1 不同海拔高度土壤种子库特征
2.1.1 土壤种子库密度由表2可知,不同海拔高度土壤种子库密度具有差异性。5个海拔高度土壤种子库总密度分别为(825±16.07)、(390±10.41)、(685±5.00)、(30±5.00)和(860±18.93)seeds·m-2,表现为2 000 m>1 200 m>1 600 m>1 400 m>1 800 m,且差异显著(P<0.05)。其中,一年生草本种子密度表现为1 200 m>1 600 m>1 400 m>2 000 m>1 800 m,多年生草本种子密度表现为1 800 m>1 400 m>2 000 m=1 200 m=1 600 m,灌木或小灌木种子库密度表现为2 000 m>1 800 m=1 200 m,且海拔间均差异显著(P<0.05)。

表2 不同海拔土壤种子库组成及密度
海拔2 000和1 200 m处土壤种子库总密度分别为(860±18.93)和(825±16.07)seed·m-2,显著高于其他海拔,但两者种子来源并不相同。海拔1 200 m处种子主要来源于一年生草本,为(810±15.28)seed·m-2,显著高于其他海拔;海拔2 000 m土壤种子库主要来源于灌木和小灌木,为(710±7.64)seed·m-2,显著高于其他海拔。除此之外,海拔1 600 m土壤种子库主要来源于多年生草本,为(120±7.64)seed·m-2,显著高于其他海拔。
2.1.2 土壤种子库多样性和相似性特征如图1所示,不同海拔高度的土壤种子库多样性具有差异性。物种多样性常用Shannon-Wiener指数、Simpson优势度指数、Pielou均匀度指数和Margalef丰富度指数来表示。5个海拔高度土壤种子库中,全部植物种Shannon-Wiener指数为1 200 m>2 000 m>1 600 m>1 800 m>1 400 m,Simpson优势度指数为1 600 m>1 200 m>1 800 m>2 000 m>1 400 m,Pielou均匀度指数为1 800 m>1 600 m>2 000 m>1 200 m>1 400 m,Margalef丰富度指数为1 200 m>2 000 m>1 600 m>1 800 m>1 400 m,且各指数在海拔间的变化差异均显著(P<0.05)。
随着海拔升高,一年生草本、灌木或半灌木的Shannon-Wiener指数和Simpson优势度指数增大,而一年生草本的Pielou均匀度指数增大,灌木或半灌木的Margalef丰富度指数和Pielou均匀度指数均减小,且海拔间有显著差异(P<0.05)。
如表3所示,海拔间植物群落土壤种子库相似性不同。海拔1 200 m土壤种子库与1 600 m土壤种子库相似性最高,相似性系数为0.45,共同物种有5种;海拔2 000 m植物群落与1 200m、1 600 m和1 800 m海拔植物群落土壤种子库相似性均较大,相似性系数分别为0.32、0.35、0.29,共同物种分别有4种、3种、2种;其余海拔间土壤种子库相似性均较低。
由于明清两朝时代接近,《四库全书总目》(以下简称《四库》)著录明代别集的数量最多,“远远超过了之前各个朝代文献数量的总和”[1],正印证了别集小序所说的“有明以后,篇章弥富”[2]12。虽然《四库》是一部官家的目录学代表作,却也涉及到文学批评的范畴;不过鉴于其官学及政治背景,提要中反映出的文学评价仍有许多问题值得探讨,而以晚明文学最为突出。

表3 不同海拔间土壤种子库相似性
2.2 不同海拔土壤种子库与相应地上植被的物种组成关系
2.2.1 土壤种子库与地上植被物种组成由表2和表5可知,不同海拔高度土壤种子库物种组成具有差异。在研究区土壤种子库中共有植物22种,隶属于7科,禾本科种类最多,为8种,藜科6种,菊科4种,蒺藜科、大戟科、豆科和玄参科各1种。其中海拔1 200 m处有植物12种,隶属于7科,其中禾本科5种,藜科2种,蒺藜科、大戟科、豆科、玄参科和菊科各1种。
1 400 m处有植物2种,均隶属于禾本科。1 600 m处有植物7种,隶属于4科,其中禾本科4种,藜科、菊科和大戟科各1种。1 800 m处有植物4种,隶属于2科,其中禾本科、菊科各2种。2 000 m处有植物10种,隶属于3科,其中藜科5种,菊科3种,禾本科2种。随着海拔升高,隶属于禾本科的物种数呈下降趋势,菊科和藜科呈增加趋势,蒺藜科、大戟科、豆科和玄参科消失。
如表4和表5所示,样地地上植被群落共出现33种植物,隶属于13科,菊科种类最多,为8种,禾本科7种,藜科4种,豆科3种,唇形科和旋花科各2种,百合科、远志科、茜草科、蔷薇科、蒺藜科、鼠李科和罂粟科各1种。其中,一年生草本共有8种,隶属于3科,藜科种类最多,为4种,菊科3种,禾本科1种;多年生草本共有17种,隶属于8科,禾本科种类最多,为6种,菊科3种,唇形科和豆科各2种,百合科、远志科和旋花科各1种;灌木或半灌木共有8种,隶属于7科,菊科种类最多,为2种,旋花科、豆科、茜草科、蔷薇科、蒺藜科和鼠李科各1种。

表4 不同海拔地上植被物种重要值

表5 不同海拔土壤种子库与地上植被物种数及其科类分布
2.2.2 土壤种子库与地上植被物种生活型构成由表6、表7可知,地上植被与土壤种子库物种生活型构成在5个海拔高度存在差异性。其中,土壤种子库物种生活型构成海拔间的变化差异显著(P<0.05),而地上植被物种生活型构成在海拔间差异不显著(P>0.05)。一年生草本在海拔1 200~1 800 m土壤种子库占比较大,但在地上植被中占比明显低于多年生草本和灌木或半灌木。在海拔1 200 m处土壤种子库密度中一年生草本种子密度最大,贡献了该海拔98.18%的种子,但地上植被占比最大的为多年生草本,这可能是由于一年生草本的繁殖策略所导致的。与其他海拔相比,海拔1 800 m土壤种子库密度中多年生草本占比相对较高,相对应的地上植被占比最大的也为多年生草本。在海拔2 000 m处土壤种子库密度中灌木或半灌木种子密度最大,贡献了该海拔82.60%的种子,但地上植被占比最大的为多年生草本,这可能是由于灌木或半灌木的繁殖策略所导致的。
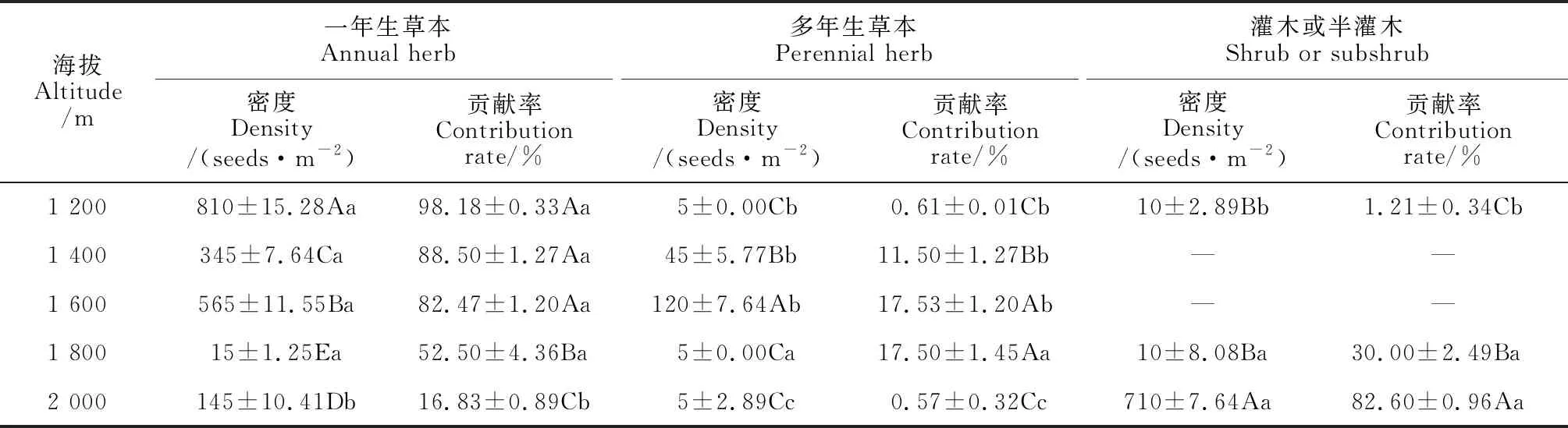
表6 不同海拔土壤种子库物种生活型构成

表7 不同海拔地上植被物种生活型构成
2.2.3 土壤种子库与地上植被耦合关系如表8所示,5个海拔高度地上植被与土壤种子库的相似性均较低,且地上植被与土壤种子库在物种组成上有显著差异。5个海拔高度地上植被与土壤种子库的相似性系数分别为0.12、0.33、0.11、0.12和0.12,共有种分别为3、2、1、1和1。海拔1 400 m相似性较高,为0.33;海拔1 600 m相似性较低,仅为0.11。说明该研究区不同海拔高度地上植被群落稳定性和发展趋势具有差异性。

表8 不同海拔土壤种子库与地上植被相似性
2.3 土壤种子库物种多样性与土壤理化因子的RDA分析
为了更好地揭示土壤种子库与土壤环境因子之间的关系,采用冗余分析的方法进行排序,将不同海拔高度土壤种子库物种多样性指数(Shannon-Wiener指数、Simpson优势度指数、Pielou均匀度指数和Margalef丰富度指数)作为响应变量,土壤各理化指标作为解释变量,进行RDA分析,筛选能明显影响土壤种子库变化的土壤理化因子。由表9可知,RDA排序图的前两个排序轴特征值分别为0.708和0.248,相对应的物种数据变量积累百分比分别为70.8%和95.6%,故轴1能较好地反映土壤种子库物种多样性与土壤理化因子的变化特征。由图2和表10可知,土壤种子库物种多样性指数与土壤容重、pH、有机质、全碳、全氮、碱解氮、速效磷呈正相关,与含水率、总孔隙度、电导率、碳氮比呈负相关,其中与pH呈显著正相关,与电导率呈显著负相关,即pH、电导率对土壤种子库物种多样性的影响较大。

表9 土壤种子库物种多样性与土壤理化因子的RDA排序分析
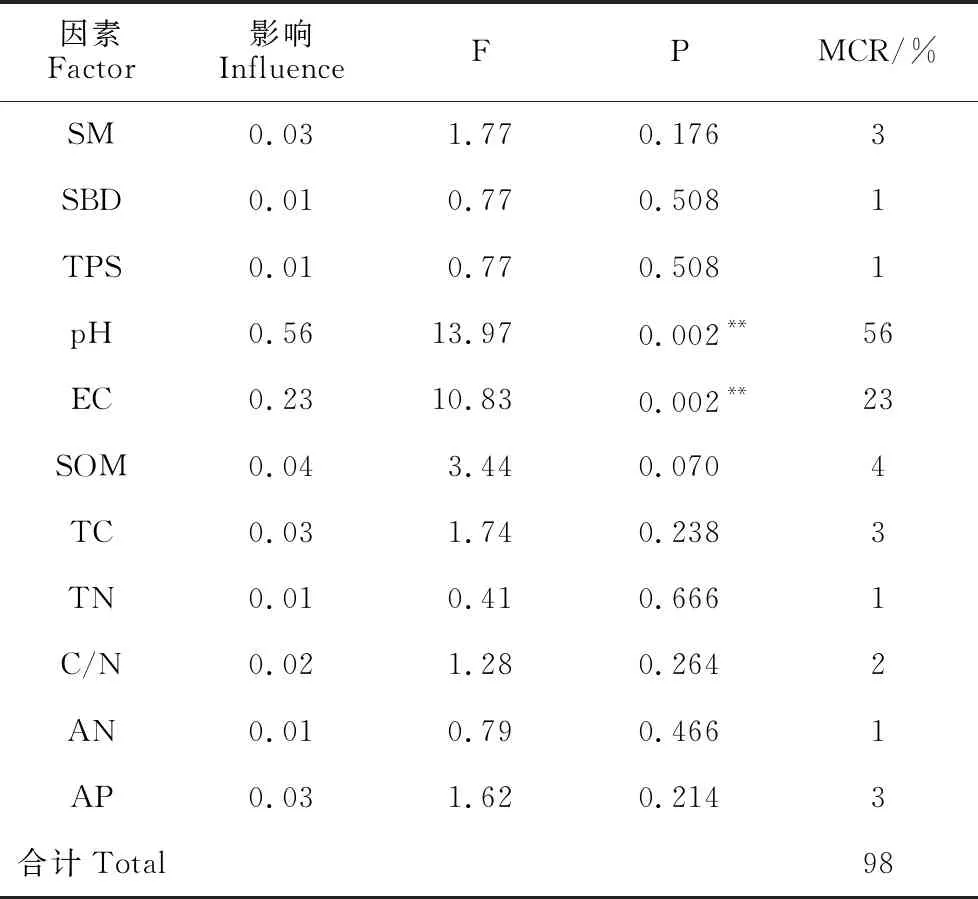
表10 土壤理化因子对土壤种子库物种多样性的RDA条件解释变量
3 讨论与结论
3.1 不同海拔土壤种子库特征
土壤种子库特征显著影响该生态系统稳定性、演替趋势、植被恢复潜力[21]。在本研究中,5个海拔高度的土壤种子库种子密度、物种组成、生活型、多样性均存在差异。海拔等地形因素显著影响土壤种子库种子密度[22]。海拔2 000 m和1 200 m处土壤种子库密度显著高于其他海拔,但其种子来源并不相同,海拔1 200 m处种子主要来源于一年生草本,海拔2 000 m土壤种子库主要来源于灌木或半灌木,海拔1 600 m土壤种子库主要来源于多年生草本。种子库萌发物种以禾本科草本植物为主。随着海拔升高,隶属于禾本科的物种数呈下降趋势,菊科和藜科物种呈增加趋势,蒺藜科、大戟科、豆科和玄参科物种消失。
土壤种子库间相似性可用来预测该植物群落发展、演替。在本研究中,5个海拔高度的植被群落土壤种子库相似性具有差异性,海拔2 000 m植物群落与其他海拔植物群落土壤种子库相似性均大于该海拔与其他海拔植物群落土壤种子库相似性,可能是因为海拔2 000 m植物群落土壤种子库中优势种画眉草、猪毛菜常见于其他海拔。
土壤种子库多样性指数常用来表征植物群落潜在种群稳定性,可用来预测群落演替方向等。物种多样性常用Shannon-Wiener指数、Simpson优势度指数、Pielou均匀度指数和Margalef丰富度指数来表示。在本研究中,随着海拔升高,一年生草本、灌木或半灌木的Shannon-Wiener指数和Simpson优势度指数显著增加,一年生草本的Pielou均匀度显著增加,灌木或半灌木的Margalef丰富度指数和Pielou均匀度指数显著降低。海拔1 400 m处土壤种子库的稳定性指数、生态优势度指数、均匀度指数和物种丰富度指数均显著低于其他4个海拔,即海拔1 400 m处土壤种子库多样性最低。海拔2 000 m处土壤种子库表现为一年生草本和灌木或半灌木的物种多样性有更高的稳定性、生态优势度,一年生草本分布均匀,可能是因为海拔2 000 m处虽然物种数量多,但灌木或半灌木植物物种数量占优势。贺兰山低山区土壤种子库多样性受自身物种组成影响,还可能受地上植被制约,需进一步研究。
3.2 不同海拔土壤种子库与相应地上植被的物种组成关系
地上植被与土壤种子库物种组成显著影响生态系统稳定性、演替趋势、植被恢复潜力[21]。在本研究中,地上植被与土壤种子库在密度、物种组成、物种多样性以及相似性均具有较大差异,与Smith[23]、张起鹏等[24]研究结果一致,与Yuan[25]、Valk[26]不一致。本研究区内,贺兰山5个海拔高度地上植被共计调查33种植物隶属于13科,土壤种子库共计萌发22种植物隶属于7科,各海拔高度土壤种子库物种数均显著低于地上植被,表明贺兰山不同海拔典型植物群落地上植被与土壤种子库的物种组成差异较大。对5个海拔高度进行植被调查和土壤种子库萌发实验,发现禾本科、菊科、藜科植物在地上植被群落和土壤种子库中出现比例均较高,且随着海拔升高,禾本科和藜科植物占比减少,菊科植物占比增加。说明禾本科、菊科、藜科植物在贺兰山适应度相对较高,且随海拔变化而变化,繁殖策略偏向于种子繁殖。在物种生活型组成上,土壤种子库中物种数占比最大为一年生草本,地上植被为多年生草本,差异较大,可能是因为植物的不同繁殖策略引起的,一年生草本生活史较短,种子小且量大,多年生草本、灌木或半灌木倾向于量小种子大的繁殖策略。
地上植被与土壤种子库相似性可用来衡量该植物群落稳定性和预测该群落的发展趋势[27]。在本研究中,有6种植物共同出现在土壤种子库和地上植被中,16种植物仅出现在土壤种子库中,27种植物仅出现于地上植被中,土壤种子库和地上植被之间具有较低的相似性,除海拔1 400 m的相似性指数为0.33外,其余海拔高度相似性系数均为0.11和0.12。地上植被与土壤种子库相似性较低,与齐丹卉[28]、张志明等[29]研究结果一致。可能是因为贺兰山地处干旱半干旱区,气候干旱少雨,种子在土壤种子库中的持久性受到影响;也可能是由于地上植被的物种表现为以多年生草本占优势,其繁殖策略形成的种子量少,对土壤种子库形成的贡献率较低。
植被恢复的潜力与土壤种子库种子密度和物种多样性密切相关[30]。土壤种子库物种多样性既受地上植被特征制约,同时也受土壤特性影响[31]。在本研究中,pH、电导率对土壤种子库物种多样性的影响较大,与王晓荣[32]、翟付群等[33]研究结果一致。可能是通过影响种子形成、储存、萌发等过程直接或间接影响土壤种子库,关于这方面的问题还需进一步研究。在本研究中,5个海拔高度土壤种子库密度均较低,土壤种子库种源尚不能满足地上植被恢复所需种源量;5个海拔高度土壤种子库物种多样性均较低,无法满足植被自然恢复需求,特别是海拔1 400 m处物种多样性显著低于其他海拔,恢复潜力最低。贺兰山低山区风蚀严重、植被稀疏,土壤种子库自身所具有的种子无法满足植被自然恢复需求,可采取飞播等生态恢复措施来弥补表层土壤种子的不足,从而满足地上植被恢复所需种源量。关于种子种类的选取和实际应用时的外界因素的干扰,还需进一步研究。
