甘加藏羊发情周期和乏情期HPO轴中THRβ mRNA和蛋白的表达分布
2022-03-28刘莉莉高何璇杨亚文包莹莹何玉琴杨志杰陈卫刚葛文博
刘莉莉,高何璇 ,杨亚文,包莹莹,何玉琴,*,杨志杰,陈卫刚,葛文博
(1.甘肃农业大学 生命科学技术学院,甘肃 兰州 730070;2.兰州银行 网络金融部,甘肃 兰州 730070;3.甘肃农业大学 动物医学院,甘肃 兰州 730070)
高原甘加藏羊是典型的季节性发情哺乳动物,生活在甘南藏族自治州海拔2 800 m以上的青藏高原天然草场高寒牧区,是甘肃优良的地方品种,也是我国宝贵的畜牧资源。甘加藏羊2.5岁开始配种,发情周期平均为(16±2)d,每年产羔一次,每次一羔,繁殖率低,影响了藏羊农牧业的快速发展。因此,研究与藏羊繁殖生理活动有关的生殖激素的调控作用机理,提高繁殖率扩大种群内优秀种羊数量,对甘肃草地畜牧业的发展具有非常重要的意义。
甲状腺激素(thyroid hormone,TH)包括三碘甲腺原氨酸 (triiodothyronine,T3)、四碘甲腺原氨酸 (thyroxine,T4)。TH 通过其特异性受体THRβ(thyroid hormone receptor β,THRβ)对动物生长发育、生殖活动等具有重要的调控作用。THRβ属于核受体超家族成员。有文献报道,THRβ在人和啮齿类动物下丘脑弓状核、室旁核、视上核及腹正中核等与生殖和代谢有关的核团上有特别高的表达,在黄金仓鼠整个发育过程中下丘脑、垂体、睾丸、卵巢有THRβ的表达。T3可以通过THRβ对下丘脑的黑色素瘤转移抑制基因()表达和亲吻素(kisspeptin)的蛋白表达产生影响,而基因通过其特异性G蛋白偶联受体54(GPR54)直接作用于GnRH神经元调控GnRH的脉冲释放,进而影响促卵泡素(FSH)和促黄体素(LH)的分泌,对哺乳动物初情期的启动,发情排卵等生殖活动发挥着重要作用。以上报道提示,TH及THRβ参与调控哺乳动物的繁殖活动。但目前有关TH及其THRβ对甘加藏羊繁殖活动的调控研究尚未见报道。本研究首次从基因和蛋白水平对甘加藏羊发情周期和乏情期松果体、HPO轴组织中mRNA及其蛋白的表达和分布规律进行了研究,以期为进一步阐明TH对藏羊生殖活动的调控及其机理研究提供科学依据。
1 材料与方法
1.1 试验动物与样品采集
在甘加藏羊非繁殖季节(3—4月)和繁殖季节(7—9月),于甘肃省甘南藏族自治州夏河县甘加乡海拔2 800 m以上的藏绵羊养殖合作社随机选择2.5~3.5岁雌性健康且未孕的雌性甘加型藏绵羊30只,分为5组,每组6只。在藏羊繁殖季节,参照四期分法,根据公羊爬跨试情以及阴道脱落细胞图片观察,将发情周期分为发情期(0—36 h)、发情后期(39—72 h)、间情期(4—13 d)、发情前期(14—16 d)。在藏羊非繁殖季节,确定乏情期为试验对照组。采用颈部局部麻醉颈动脉放血法无痛处死试验羊,将处死的试验羊迅速解剖,取出各时期松果体、下丘脑、垂体、卵巢等组织。将所取组织平均分为2份:一份置于液氮速冻后移至-80 ℃冰箱,备用;另一份置于4%多聚甲醛中固定,备用。
1.2 甘加藏羊松果体和HPO轴组织THRβ mRNA检测
依照RNA提取试剂盒(北京,TransGen)说明书实验流程进行操作,分别提取甘加藏羊发情周期和乏情期松果体、下丘脑、垂体和卵巢组织中的总RNA,并测定RNA浓度和纯度,将测定好的总RNA溶液分别稀释至500 ng·μL。根据反转录试剂盒(北京,TransGen),反转录合成cDNA链。根据NCBI中绵羊和-mRNA CDS区序列利用Primer Premier 5.0软件进行引物设计,并由上海生工生物科技有限公司合成引物(表1)。以反转录的cDNA为模板,RT-qPCR反应总体系为20 μL:模板1 μL,上、下游引物各0.8 μL,2×TransStartm Tip Green qPCR SuperMix 10 μL,ddHO 7.4 μL。每个样品设3个重复。扩增条件:95 ℃预变性30 s;94 ℃变性5 s,60 ℃ 退火延伸30 s,循环次数45次。所得相关数据用2方法进行计算,每个基因相对表达量均以内参基因为基准进行校正。所有数据均以“平均值±标准差”表示,用SPSS 26.0软件进行单因素方差分析,显著性水平选定为=0.05。

表1 THRβ和β-actin基因引物序列Table 1 Primers sequences of THRβ and β-actin gene
1.3 甘加藏羊松果体和HPO轴组织THRβ蛋白检测
用蛋白裂解液(1 mL裂解液+10 μL蛋白酶抑制剂)(Bioss,北京)提取各组织总蛋白,以BCA试剂盒(Bioss,北京)测定浓度,于十二烷基硫酸钠(sodium dodecy sulfate,SDS)上样缓冲液中100 ℃水浴 10 min 变性蛋白,冰上静置5 min备用。制备12%封底胶、12%分离胶和5%浓缩胶后以6 μL上样量进行SDS-PAGE电泳,电泳完毕对照Marker预染条带切取THRβ(53 kDa)、β-actin(42 kDa)对应条带,湿转法将分离胶上的蛋白转移至PVDF膜(Solarbio,北京)上。PBST(PBS+tween 20)(Bioss,北京)洗膜20 min,5%脱脂奶粉室温封闭1 h。孵育一抗(1∶500),4 ℃过夜。PBST洗膜4 h,37 ℃孵育二抗(1∶4 500)1 h,PBST洗膜4 h,暗室曝光成影拍照。利用Image Pro-Plus 6.0图像分析系统进行灰度值分析,并利用SPSS 26.0软件进行差异性分析。
1.4 甘加藏羊松果体和HPO轴组织THRβ免疫组织化学染色
将4%甲醛固定好的松果体、下丘脑、垂体、卵巢组织包埋制作5 μm厚石蜡切片,常规脱蜡处理后微波法抗原修复10 min,再用3% HO湿盒37 ℃孵育15 min,正常山羊血清工作液(A液)37 ℃封闭15 min,THRβ一抗稀释液(兔抗羊,1:350),阴性对照组一抗用PBS代替,4 ℃孵育过夜后37 ℃孵育1 h,然后用生物素标记的二抗(B液,山羊抗兔IgG)37 ℃孵育15 min,再用辣根过氧化物酶标记的链霉卵白素亲和工作液(C液)37 ℃孵育15 min。每次孵育后均用PBS(0.01 mol·L,pH=7.4)洗涤3次,每次5 min。DAB显色1 min,苏木精复染1 min,返蓝,脱水透明后中性树胶封片。免疫组织化学染色结果用Motic SK210型光学显微镜观察拍照。
2 结果与分析
2.1 甘加藏羊松果体、HPO轴组织中THRβ mRNA及其蛋白表达分析
RT-qPCR、Western blot检测结果显示,甘加藏羊发情周期和乏情期松果体、下丘脑、垂体、卵巢组织中均有mRNA和蛋白的表达,且各时期表达差异明显(图1和图2)。松果体中mRNA和蛋白表达量均在乏情期最高,显著(<0.05)高于发情周期;下丘脑中mRNA表达量在发情后期最高,与其他各时期差异显著(<0.05),蛋白表达量在间情期最高,与乏情期差异不显著(>0.05),与其他3个时期差异显著(<0.05);垂体中mRNA和蛋白表达量均在间情期最高,其中蛋白表达量在间情期与发情前期差异不显著(>0.05),其他各时期差异均显著(<0.05);卵巢中mRNA和蛋白表达量均在发情后期最高,与其他各时期差异显著(<0.05)。
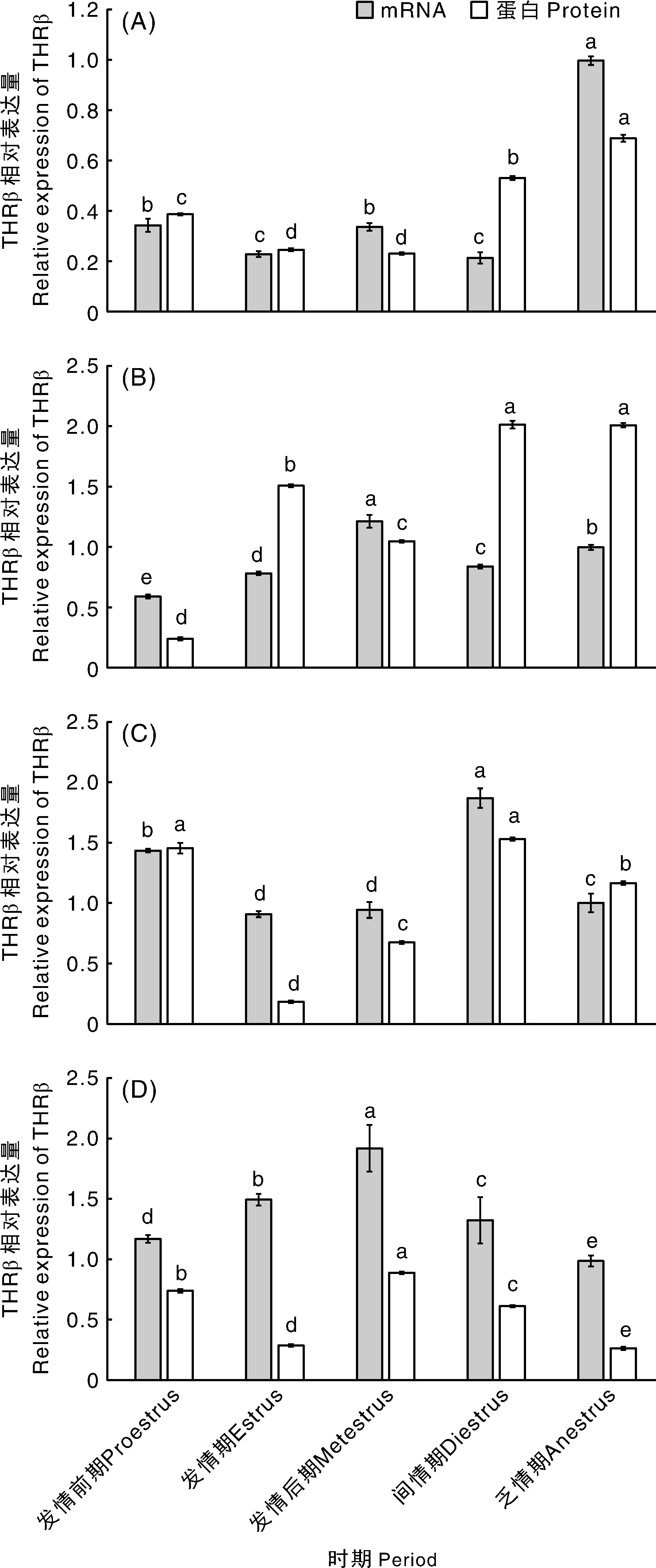
A,松果体;B,下丘脑;C,垂体;D,卵巢。同一系列柱上无相同字母的表示各时期mRNA或蛋白的相对表达量差异显著(P<0.05)。A,Pineal gland;B,Hypothalamus;C,Pituitary;D,Ovary.Bars marked without the same letters on the same series indicated significant differences in the relative expression of mRNA or protein at different periods at P<0.05.图1 甘加藏羊发情周期和乏情期松果体和HPO轴组织中THRβ mRNA和蛋白相对表达量Fig.1 Relative expression of THRβ mRNA and protein in Pineal gland and HPO axis during estrus cycle and anestrus of Ganjia Tibetan sheep
2.2 THRβ免疫阳性反应产物在甘加藏羊松果体和HPO轴组织中的分布
免疫组化试验结果显示,松果体、HPO轴组织中THRβ免疫反应产物为黄色或棕黄色,分布细胞浆和细胞核,所有对照组试验的标本均呈阴性,证明所用抗体具有THRβ免疫反应的特异性。
THRβ免疫阳性产物松果体组织中主要分布在松果体细胞胞浆(图3中A~E),细胞呈卵圆形或圆形,其中在发情后期(图3中C)着色深、强阳性表达。下丘脑组织中THRβ主要分布在神经胶质细胞、大神经元胞体和椎体细胞胞浆中有阳性表达(图3中F~J),其中在发情前期(图3中F)和发情期(图3中G)阳性表达强,染色深。在腺垂体组织中,THRβ主要分布在嗜酸性和嗜碱性细胞胞浆(图3中K~O),其中在发情期(图3中L)弱着色,在其它四个时期有强阳性表达、浓染。卵巢组织中,THRβ免疫阳性细胞在卵泡颗粒层、卵泡内膜、黄体细胞中均有分布(图3中P~T),其中在颗粒层细胞和黄体细胞免疫阳性产物着色深,高表达。
3 结果与讨论
本研究结果表明,甘加藏羊发情周期各个时期和乏情期mRNA及其蛋白在松果体 和HPO轴组织中的相对表达量呈动态变化,且存在一定的差异,这种表达的差异从基因和蛋白水平证明了THRβ参与甘加藏羊生殖调控,为进一步研究甲状腺激素及其受体对动物生殖生理活动的调控机制提供了科学的参考依据。
3.1 甘加藏羊松果体组织中的THRβ表达对藏羊繁殖的影响
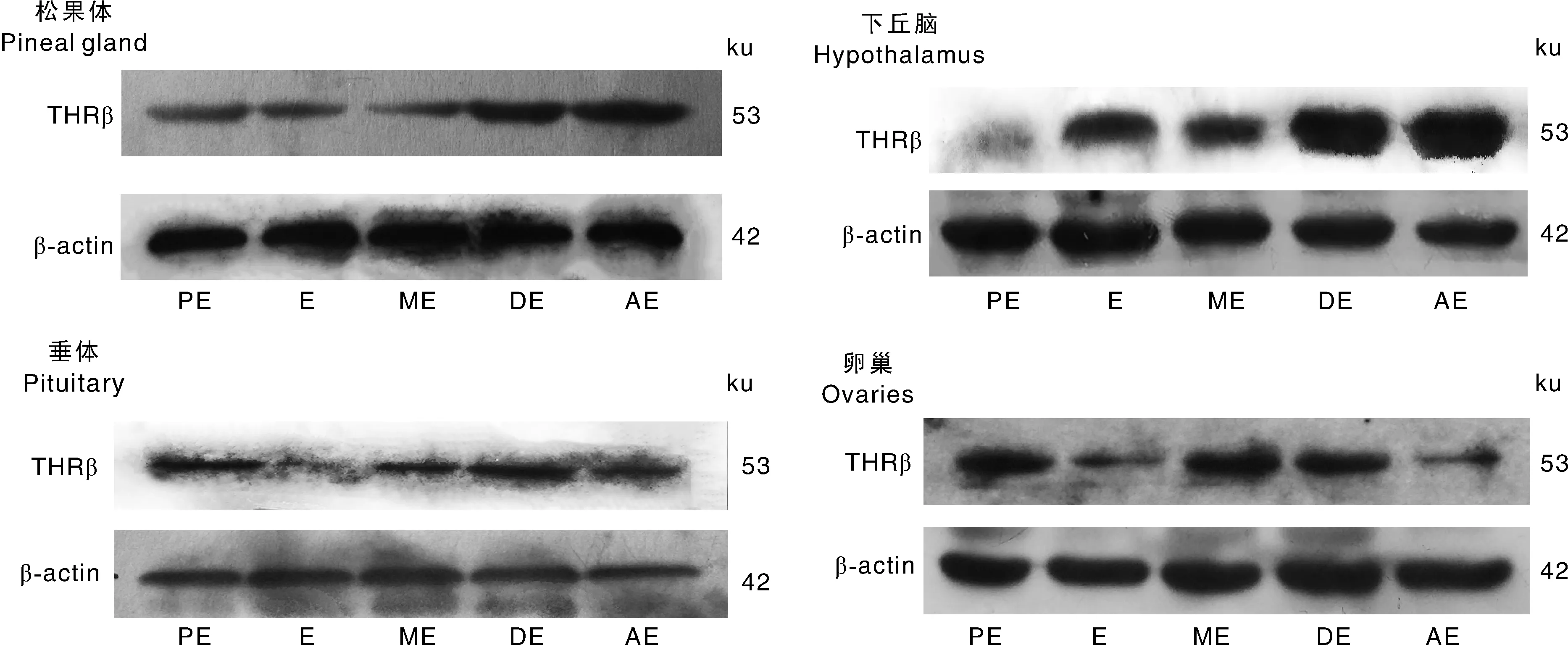
PE,发情前期;E,发情期;ME,发情后期;DE,间情期;AE,乏情期。PE,Proestrus;E,Estrus;ME,Metestrus;DE,Diestrus;AE,Anestrus.图2 甘加藏羊发情周期和乏情期松果体、HPO轴中THRβ的蛋白含量Fig.2 Content of THRβprotein in pineal gland and HPO axis during estrus cycle and anestrus of Ganjia Tibetan sheep
松果体是动物体内重要的神经内分泌器官,光周期和季节变化可以调节松果体节律性分泌褪黑素(MT)来调控动物的生殖等活动,MT作用于垂体结节部的促甲状腺激素和脱碘酶解码外界光信号,并通过血液中TH 与作用于下丘脑内侧基底部(MBH)正中隆起上的甲状腺激素受体结合,从而激活整个HPO轴,调控HPO轴相关生殖激素的变化。有研究报道显示,松果体分泌的MT对TH具有调节作用,张明泉等以人和大鼠作为研究对象,以对光线敏感的松果体为突破口,证实冬夏季节机体内TH的变化与松果体的高位调节有关。康玉明等通过去除大鼠松果体,观察大鼠甲状腺形态和TH分泌功能的变化,证实松果体可能通过抑制大鼠甲状腺分泌TH来调控动物的繁殖活动。目前已有报道显示,TH及其受体THRβ对动物的生殖活动具有调控作用,尤其在启动和终止季节性发情哺乳动物的发情过程中发挥着非常重要的作用。本研究首次对藏羊松果体组织进行免疫组织化学染色发现,THRβ在甘加藏羊整个发情周期和乏情期松果体组织的松果体细胞胞浆均有阳性表达,其中在发情后期强表达。传统的观点认为,甲状腺激素受体(THR)属于核受体,T3 通过结合到胞核内的THRβ 而起作用。但已有研究报道指出,在大鼠背侧海马神经元细胞中,TH被证实可通过与胞浆内THRβ结合,并快速激活 PI3K/AKT-mTOR 信号通路,且这种快速激活是不依赖于蛋白质和 RNA 的合成,而本研究也证实THRβ是在甘加藏羊松果体细胞胞浆发挥其调控作用,提示TH也可产生非基因组效应。且mRNA及其蛋白表达量在发情前期到发情期呈现下降趋势,从间情期开始呈现上升趋势,在乏情期表达最高,THRβ在松果体组织中的这种动态差异表达提示其参与调控藏羊的发情周期生殖活动。
3.2 甘加藏羊HPO轴组织中的THRβ表达对藏羊繁殖的影响
雌性哺乳动物的生殖生理活动受HPO轴的调控,且有多种激素和其它因子的协调参与,其中下丘脑GnRH的脉冲式分泌是激活动物发情周期的关键,其脉冲释放增加引起青春期启动,升高的LH作用于卵巢产生雌二醇(E2),促进青春期发育与成熟,对维持正常的性功能和生殖功能起重要的作用。有研究发现,下丘脑内注射T3能影响 LH 分泌,而LH是HPO轴调控动物季节性动情节律的关键激素,提示T3可能参与调控季节性发情动物的繁殖活动。已有研究证实在黄金仓鼠HPGA发育的整个过程中其下丘脑、垂体、卵巢中均有THRβ的表达。本研究结果显示,THRβ在甘加藏羊发情周期各个时期HPO轴组织中均有表达,且不同组织THRβ的表达量有显著差异,提示THRβ参与调控甘加藏羊的生殖活动。对下丘脑组织免疫组织化学染色发现,在下丘脑的神经细胞和椎体细胞中均有THRβ阳性表达,证实下丘脑组织中THRβ是在这两种细胞中发挥其生物学功能。研究发现,mRNA的相对表达量从发情前期开始呈现上升趋势,在发情后期达到最大值,而蛋白表达量在间情期达到最大值。基因和蛋白表达出现偏差的主要原因一方面可能是基因表达的内源机制,另一方面可能是由于一些特殊外在环境因素所影响,比如藏羊高海拔的生存环境等可能对藏羊机体中从基因到蛋白的表达产生影响。已有研究证实在啮齿动物和绵羊大部分下丘脑KISS1细胞均表达雌激素受体α(ERα),而雌激素的反馈作用正是由下丘脑中 Kisspeptin-GPR54信号系统实现,T3可以通过THRβ对下丘脑中GT1-7神经元细胞的1 基因表达产生影响,且T3缺乏可引起发育期大鼠大脑基因表达的升高,由此提示卵巢分泌的雌激素在发情后期和间情期通过调控下丘脑THRβ的表达量来影响GT1-7神经元基因的表达进而实现对下丘脑 GnRH 的反馈调节,影响垂体FSH和LH的分泌,调控动物的生殖活动。

A、F、K、P,发情前期;B、G、L、Q,发情期;C、H、M、R,发情后期;D、I、N、S,间情期;E、J、O、T,乏情期。P,松果体细胞;BV,血管;GC,神经胶质细胞;NC,神经元;Nu,神经细胞核;AC,嗜酸性细胞;BC,嗜碱性细胞;CC,嫌色细胞;FA,卵泡腔;GV,卵母细胞核;LC,黄体细胞;TF,卵泡膜;SG,颗粒层。A,F,K,P,Proestrus;B,G,L,Q,Estrus;C,H,M,R,Metestrus;D,I,N,S,Diestrus;E,J,O,T,Anestrusl.P,Pineal cell;BV,Blood vessel;GC,Glial cell;NC,Nerve cell;Nu,Neuronuclei;AC,Acidophilic cell;BC,Basophilic cell;CC,Chromophobe cell;FA,Follicular antrum;GV,Germinal vesicle;LC,Luteal cell;TF,Theca folliculi;SG,Stratum granulosum.图3 甘加藏羊发情周期和乏情期松果体和 HPO轴组织中THRβ免疫阳性反应产物分布Fig.3 Location of THRβ immunoreactive products of in pineal and HPO axis of Ganjia Tibetan sheep during the estrus cycle and anestrus
垂体作为机体内重要的内分泌腺,接受来自下丘脑分泌的GnRH,促进 FSH和LH的分泌,进而影响卵巢性激素的合成和分泌,参与调控动物的生殖生理活动。Hodin等人在大鼠垂体中检测到T3的功能性受体mRNA的表达。而本研究结果显示,在垂体组织的远侧部的嗜酸性细胞、嗜碱性细胞的细胞浆中均有THRβ的阳性表达,其基因和蛋白表达量均从发情前期开始呈现下降趋势,在发情期达到最小值,之后开始呈现上升趋势,在间情期达到最大值,提示THRβ在垂体组织中主要在间情期的嗜酸性和嗜碱性细胞中参与调控甘加藏羊生殖活动。主要原因可能是雌激素的反馈调节。已有研究发现,在大鼠垂体前叶中检测到T3的功能性THRβ,而垂体前叶是负责调节促性腺激素释放激素合成和分泌的区域,其分泌的LH是下丘脑-垂体-卵巢轴调控雌性动物发情、排卵等生殖活动的桥梁,且包莹莹等的研究发现,在藏羊垂体组织中也检测到基因和蛋白的表达,而THRβ能诱导ERα的转录,接受来自性腺激素的反馈调节,再次证实本研究结果。
卵巢作为主要的雌性生殖器官,对下丘脑和垂体所释放的生殖激素较为敏感,其释放的E2转而调控自身或对垂体和下丘脑进行反馈调节,继而参与机体生殖过程。Vissenberg等人发现,在卵巢上存在甲状腺激素受体和甲状腺激素转运体的表达,其通过对105篇文献灰度总结,数据显示甲状腺激素水平的改变影响卵泡发育、精子生成,降低受孕率,T3 可以促进颗粒细胞增殖,并通过 PI3K/Akt信号通路抑制其凋亡。本研究对藏羊的卵巢进行免疫组织化学染色,发现在卵巢组织的卵泡颗粒细胞胞浆和胞核、卵泡内膜细胞及黄体细胞胞浆中均有THRβ的阳性表达,其中在卵泡颗粒层细胞和黄体细胞胞浆有强阳性表达,提示THRβ参与调控卵泡的发育和黄体的成熟。而文献报道显示卵巢分泌的雌激素的反馈作用是通过Kisspeptin/GPR54信号系统实现的,而T3能通过THRβ影响1基因启动子活性来实现对mRNA 表达和 Kisspeptin 蛋白表达的调控。而本研究也证实mRNA和蛋白表达量均在发情后期达到最大值,在乏情期达到最小值,由此提示卵巢中THRβ可能通过卵巢中的雌激素反馈作用于下丘脑kisspeptin,参与GnRH脉冲释放的神经内分泌过程来实现对性腺系统的繁殖调控。且有研究证实在甘加藏羊整个发情周期卵巢颗粒细胞上存在雌激素受体(ERs)、KISS1和GPR54的表达。综上所述,THRβ主要在发情后期调控甘加藏羊的生殖活动,参与调控卵泡发育和排卵等生殖活动。
