稻米蒸煮特性QTL定位及与感官食味品质的相关性分析
2022-03-11徐伟清王小雷欧阳林娟李炜星欧阳清兰贺浩华朱昌兰
徐伟清 王小雷 刘 杨 欧阳林娟 李炜星 欧阳清兰 贺浩华 朱昌兰,*
(1江西农业大学 作物生理生态与遗传育种教育部重点实验室/江西省超级稻工程技术研究中心,江西 南昌 330045; 2赣州市农业农村局,江西 赣州 341000)
水稻(OryzasativaL.)是一种重要的谷物作物,为世界一半以上的人口提供主食。杂交稻生产技术的成功应用,使水稻产量得到了极大提高,但对稻米品质的改良仍不尽人意,稻米品质仍有较大的提升空间[1]。稻米的品质性状包括外观品质、加工品质、蒸煮食味品质、营养品质和特殊品质,其中广大消费者最关心的是稻米的蒸煮与感官食味品质。已有研究表明,稻米的蒸煮和感官食味品质遗传较复杂,受多种因素影响[1-2]。淀粉是稻米的主要组成成分,约占稻米物质组分的90%[3-6]。前人对稻米淀粉理化特性与食味品质之间的关系进行了大量研究,认为直链淀粉含量(amylose content,AC)是影响稻米食味品质的最主要因素,凝胶稠度(gel consistency,GC)和糊化温度(gelatinization temperature,GT)也是重要影响因素[7]。参与淀粉生物合成的基因对稻米食味品质起着重要的调控作用[8],目前有20多个调控稻米淀粉生物合成的基因已经鉴定和克隆,包括Wx[9]、ALK[10-13]、BEIIb[14]、RAG2[15]、SSG4[16]、Du1[17]、SSI[18]、SSII[19]、SSIII[11-12]、ISA[19]、AGPlar[19]、AGPiso[13]、PUL[19]等。
鉴于淀粉理化特性与稻米食味品质性状的关系,以往对稻米食味品质的遗传调控研究主要集中在直链淀粉含量、糊化温度、凝胶稠度等间接理化指标,对于稻米吸水率(water absorption,WA)、延伸率(cooked rice elongation,CRE)和膨胀率(volume expansion,VE)等蒸煮特性的遗传调控研究较少。研究发现在煮熟的稻米中,高品质的水稻品种具有更大的延伸长度、更小的膨胀体积和吸水率[20]。已利用不同遗传群体在1~12号染色体上定位到稻米吸水率、延伸率、膨胀率相关QTL(quantitative trait locus)[21-32],多数QTL的效应较小,少数效应较大的QTL能在不同群体中重复检测到,如姜树坤等[29]利用粳粳交构建的F2:3群体对稻米的延伸性进行研究,共定位到8个与稻米延伸性相关的QTL,其中分布在第1、第2、第6和第11号染色体上的QTL与Tian等[21]、沈圣泉等[22]、张光恒等[23]、何予卿等[30]报道的QTL位置相同,这些研究为了解稻米蒸煮特性的遗传特性提供了一定依据,但关于其分子调控机制仍有待研究。因此,挖掘不同遗传资源中的稻米蒸煮特性相关QTL,不仅有助于深入了解稻米蒸煮特性的遗传机制,也可为遗传育种提供更多的优异基因资源。本试验利用籼稻昌恢121和优质粳稻越光构建的56个染色体片段置换系(chromosome segment substitute lines,CSSL),对稻米蒸煮特性相关QTL定位及感官食味品质相关性进行研究,旨在进一步挖掘控制稻米蒸煮特性的优异等位基因,了解稻米蒸煮特性与感官食味品质之间的相互关系,为稻米品质的遗传改良提供一定的理论基础。
1 材料与方法
1.1 试验材料
本研究前期以籼稻品种昌恢121为受体亲本,以优质粳稻品种越光为供体亲本,经3~5次回交和7~9次自交构建了一套CSSL[33],包含56个BC3F9、BC4F8和BC5F7株系,各株系遗传稳定,结实灌浆正常。其中受体亲本昌恢121为江西农业大学水稻育种课题组选育的强优势籼稻恢复系,培育出农业部超级稻淦鑫688等一系列强优势杂交稻组合[34];供体亲本越光为日本优质粳稻品种,其口感柔软富弹性,味道上佳。所有材料于2018年夏季在南昌江西农业大学试验田种植。每个株系种植3个重复,每个小区种植3行,每行10株,株行间距分别为17和20 cm,田间管理遵循正常的水稻生产程序。
1.2 56个置换系的图示标记基因型
利用135个两亲本间有多态性的分子标记对56个CSSL进行基因分型,56个株系的基因分型结果见图1。共135个有多态的分子标记,包含99个简单序列重复标记(simple sequence repeats,SSR)和36个序列标签位点(sequence - tagged site,STS),均匀覆盖在12条染色体上,覆盖水稻全基因组的1 427.7 cM,平均间距为10.5 cM,最大间距为35.8 cM。
1.3 稻米蒸煮特性的测定
稻米蒸煮特性的测定参照刘宜柏等[35]和Tian等[21]的方法,并做适当的修改。每个株系随机挑选15粒整精米,首先用游标卡尺测量其长、宽、厚,用电子分析天平称重,取其平均值作为该株系米粒的粒长、粒宽、粒厚和粒重,蒸煮前米粒体积=蒸煮前米粒长×米粒宽×米粒厚。然后将这15粒整精米转移到试管中,加满水后静置30 min,于沸水浴中煮10 min,将米饭取出,用吸水纸吸干表面水分,转移到垫有干滤纸的培养皿中,放入恒温干燥箱(19℃)中干燥30 min使吸附在米饭表面的水分被完全吸干,分别测量米饭的长、宽和厚,称重,取平均值作为该株系蒸煮后米粒的粒长、粒宽、粒厚和粒重,蒸煮后米粒体积=粒长×粒宽×粒厚。按下列公式计算稻米的吸水率、延伸率和膨胀率:
吸水率=(蒸煮后米粒重-蒸煮前米粒重)/蒸煮前米粒重×100%
(1)
延伸率=(蒸煮后米粒长-蒸煮前米粒长)/蒸煮前米粒长×100%
(2)
膨胀率=(蒸煮后米粒体积-蒸煮前米粒体积)/蒸煮前米粒体积×100%
(3)。
1.4 稻米蒸煮工艺流程及感官食味品质的评定
稻米蒸煮工艺流程:称取大米样品300 g→淘洗2次除去杂质和异味→注水量(米水比1∶1.2)→浸泡30 min→电饭煲蒸煮(电饭煲型号:CFXB-35A,生产商:中山市金美誉电器有限公司)→保温15 min。感官食味品质的评定主要按照GB/T 15682-2008粮油检验稻谷、大米蒸煮食用品质感官评价方法[36]的评分方法二进行,同时结合该套CSSL材料特点进行适当修改。感官评分值参照张亚东等[37]的方法,对米饭的外观、香味、味道、口感及感官综合评分与参照样品(亘古丝苗)进行比较,然后得出各株系的评价分。评价分为7个等级,分别为“相当好、好、略好、相同、略差、差、相当差”,记作3、2、1、0、-1、-2、-3分。感官评分值=70+平均得分×10,满分为100分。
1.5 统计分析
采用Microsoft Excel 2003进行数据录入、处理与作图;显著性使用SPSS 16.0进行t检验分析(P<0.01具有极显著性差异,0.01
2 结果与分析
2.1 亲本与CSSL感官食味品质的比较
对亲本与CSSL的感官食味品质进行比较(表1),结果表明,除米饭外观性状在两亲本间未达到显著性差异,其他感官食味品质性状在两亲本间均达到极显著差异,尤其味道和口感在两亲本间存在较大差异,其中味道在昌恢121和越光的分值分别为-0.77和0.54,相差1.31。CSSL的感官食味品质性状均出现超亲现象,变异系数(coefficient of variation,CV)在0.29%~4.09%之间,其中外观、香味和味道的CV均大于1。

注:黑色方块表示越光标记基因型,白色方块表示昌恢121标记基因型,灰色方块表示杂合标记基因型,图中左侧表示56个 CSSL株系的名称,图中上侧表示水稻12条染色体号及12条染色体上包含的99个SSR标记和36个STS标记。Note: The black, white and gray areas represenTThe marker genotype of Koshihikari, Changhui121 and heterozygote, respectively,the left side of the figure shows the names of 56 CSSL lines, and the upper side of the figure shows the 12 chromosome numbers of rice and 99 SSR markers and 36 STS markers contained on the 12 chromosomes.图1 56个CSSL株系的图示标记基因型Fig.1 Graphic marker genotype of 56 CSSL
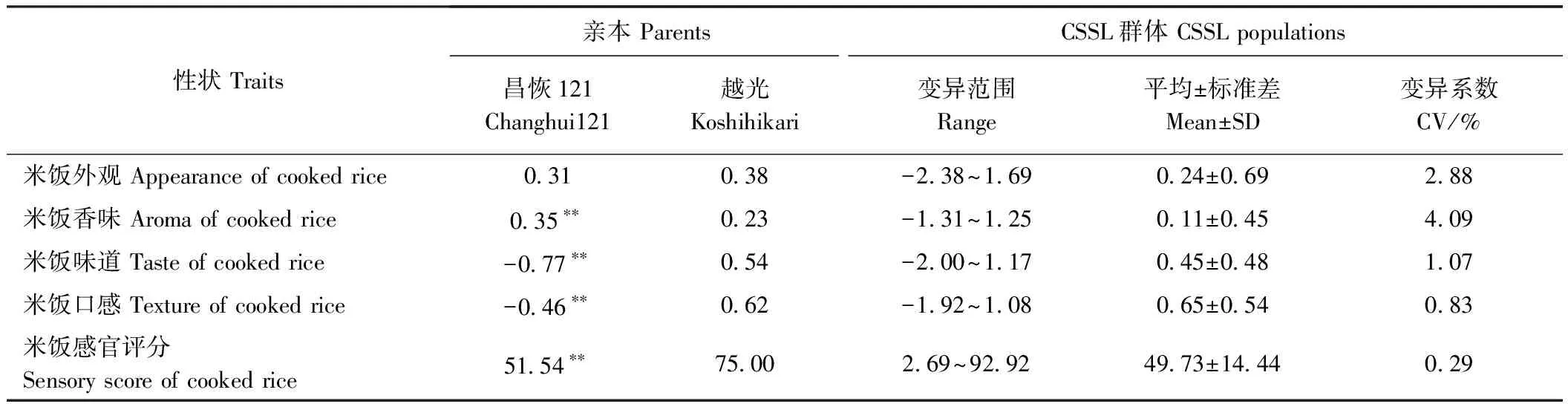
表1 亲本及CSSL的稻米感官食味品质比较Table 1 Comparison of rice sensory eating quality between parents and CSSL
2.2 稻米蒸煮特性在CSSL和亲本中的表现
由图2可知,稻米的吸水率、延伸率和膨胀率在两亲本间具有明显的差异,其中吸水率在两亲本之间的差异最大,并且其表型变异幅度也最大。吸水率、延伸率和膨胀率在56个CSSL中均存在广泛的分离和超亲变异。
2.3 稻米蒸煮特性与感官食味品质性状的相关性分析
对稻米蒸煮特性与感官食味品质性状间进行相关性分析(表2),发现稻米蒸煮特性与感官食味品质性状之间无相关性,但蒸煮特性中的米饭吸水率、延伸率和膨胀率之间两两呈极显著正相关,其中延伸率与膨胀率之间的相关系数最大,为0.79,表明吸水较多的稻米一般具有良好的延伸性;感官食味品质中的米饭外观、香味、味道、口感和感官评分之间两两呈极显著正相关,其中口感和感官评分之间的相关系数最大,为0.96,表明口感对感官评分的影响较大。

注:C: 昌恢121,K: 越光。Note: C: Changhui 121. K: Koshihikari.图2 稻米吸水率、延伸率和膨胀率在CSSL的分布Fig.2 Distribution of the water absorption, cooked rice elongation and volume expansion in the CSSL

表2 稻米蒸煮特性与感官食味品质性状的相关性分析Table 2 Correlation analysis between cooked quality and sensory evaluation related traits of rice
2.4 稻米蒸煮特性的QTL定位
利用56个CSSL对米饭吸水率(WA)、延伸率(CRE)和膨胀率(VE)3个稻米蒸煮特性性状进行QTL定位,共检测到4个QTL,分布于第8和第11号染色体上。其中qWA-8和qCRE-8位于第8号染色体RM5767标记附近,表型贡献率(contribution to the phenotypic variance,PVE)和加性效应值(additive effect,Add)分别为25.05%、25.94%和-25.68、-8.00,两者的增效等位基因均来自昌恢121;qCRE-11和qVE-11位于第11号染色体RM287标记附近,表型贡献率和加性效应值分别为27.95%、41.16%和8.30、30.31,两者的增效等位基因均来自越光,其中qVE-11的LOD值和贡献率均为最大,分别为6.33和41.16%,由此推测该位点可能为1个主效位点(表3)。

表3 利用56个CSSL检测到的稻米蒸煮特性QTLTable 3 QTL for cookinGCharacteristics of rice detected by 56 CSSL

图3 稻米蒸煮特性QTL在染色体上的位置Fig.3 Locations of the identified QTL for rice cooking quality on chromosome
2.5 亲本及置换株系的稻米蒸煮特性比较
根据稻米蒸煮特性的QTL定位结果可知,qWA-8和qCRE-8定位于8号染色体RM5767标记附近,由图1中的56个CSSL基因分型图谱可以发现,置换系X137在该区段插入了越光的染色体片段。观察测定该株系的米粒长度、米饭光泽度和米饭蒸煮特性,发现其米粒长度比昌恢121短,米粒蒸煮前后的光泽度高于昌恢121(图4),其米饭的延伸率小于昌恢121,差异达极显著水平(P<0.01),而米饭的吸水率与昌恢121间未达到显著性差异,可能是其他代换片段的影响;qCRE-11和qVE-11定位于11号染色体RM287标记附近,置换系X107该区段插入了越光的染色体片段。其米粒长度比昌恢121短,而蒸煮后的米粒长度比昌恢121长,其米饭的延伸率和膨胀率与昌恢121间均存在极显著差异(P<0.01)(图4)。

注: A、B:置换株系X137的吸水率和延伸率;C、D:置换株系X107的延伸率和膨胀率。E、F:稻米蒸煮前和蒸煮后,坐标纸每一个 小格的长宽为1 cm;G:米粒外观。E、F、G:从左至右分别表示为昌恢121、越光、置换系X137和X107;**表示差异极显著(P<0.01)。Note: A、B: The water absorption rate and elongation rate of the replacement strain X137, respectively. C、D: The elongation and expansion rates of the replacement strain X107, respectively. E、F: Rice quality traits before and after cooking respectively, the length and width of each small grid on the coordinate paper is 1 cm. G: Grain appearance. E、F、G: Changhui121, Koshihikari, X137 and X107 respectively from lefTTo right. ** mean significant differences at 0.01 level.图4 置换株系X137、X107与昌恢121之间的蒸煮特性表型比较Fig.4 Differences of phenotypic values of rice cookinGTraits between the strains of X137、X107 and Changhui121
3 讨论
稻米蒸煮特性与感官食味品质作为稻米品质的综合评价标准,越来越受到育种家的关注,但是关于稻米蒸煮特性与感官食味品质的评价标准还不够完全。稻米蒸煮特性遗传较为复杂,是由多个基因控制的复杂性状[3, 42]。
本研究利用前期构建的一套以籼稻昌恢121为受体亲本和优质粳稻越光为供体亲本构建的CSSL,对稻米的吸水率(WA)、延伸率(CRE)和膨胀率(VE)进行检测,共检测到4个QTL(qWA-8、qCRE-8、qCRE-11和qVE-11)。与前人的研究结果进行比较分析,发现8号染色体上检测到的qCRE-8与fgr[43]基因紧密连锁,该结果与Amarawath等[44]的研究结果一致。位于第11号染色体RM287标记处的qCRE-11和qVE-11此前还未被报道过,为新鉴定的QTL,其中qVE-11的LOD值、贡献率和加性效应分别高达6.33、41.16%和30.31,推测该位点可能为1个主效位点,该位点的增效等位基因来自越光。本研究定位到的QTL位点的稳定性仍需进行多年多点的稳定性鉴定。之后可利用X137和X107分别与昌恢121杂交构建F2分离群体对其进行验证和精细定位。
利用56个CSSL,共检测到4个QTL,分布于第8号和第11号染色体上,其中8号染色体RM5767标记附近同时检测到控制米饭吸水率和延伸率的QTL(qWA-8和qCRE-8);11号染色体RM287标记附近同时检测到控制米饭延伸率和膨胀率的QTL(qCRE-11和qVE-11),这些QTL成簇分布可能由一因多效或基因连锁引起,仍需进一步确认。通过对亲本昌恢121与置换株系的蒸煮特性表型值进行分析,发现除置换qWA-8株系X137与亲本昌恢121未出现显著性差异,可能是受到其他代换片段的影响,qCRE-8、qCRE-11和qVE-11这3个位点被置换的株系均与昌恢121有显著性差异(图4)。通过分析亲本昌恢121与置换株系的蒸煮特性表型值发现,置换了qWA-8和qCRE-8的X137株系的米粒长度比昌恢121短,米粒蒸煮前后的光泽度高于昌恢121;而置换了qCRE-11和qVE-11的X107株系的米粒长度比昌恢121短,蒸煮后的米粒长度比昌恢121长(图3)。以上结果表明qWA-8、qCRE-8、qCRE-11和qVE-11的置换的确对亲本昌恢121的蒸煮特性进行了改良,利用这些位点对品种进行改良有利于培育优质水稻品种。
稻米品质性状之间存在不同程度的相关性,某一性状的改变可能导致其他性状受到影响。Sood等[45]利用大量Basmati类型品种研究品质性状间的相关性,结果表明延伸性除与吸水性表现为显著正相关外,与米粒大小、直链淀粉含量、糊化温度及粗蛋白含量这些理化性状无相关性。本研究与上述结果相似,CRE与WA表现为显著正相关,此外与VE呈极显著正相关。同样,本研究利用该套CSSL对WA、CRE和VE 3个稻米蒸煮特性与理化性状进行相关分析,发现三者之间无相关性[46]。根据稻米蒸煮特性与感官食味品质性状之间的相关性分析,发现感官食味品质与蒸煮特性之间无相关性,但稻米蒸煮特性WA、CRE与VE之间两两呈极显著正相关;稻米感官食味品质性状AP、ARM、TA、TE和EQ之间两两呈极显著正相关。因此,在今后的品种改良和优质品种培育中,可以结合稻米品质性状之间的相关性,对稻米品质的目标性状进行改良。
4 结论
本研究利用籼稻昌恢121为受体亲本和优质粳稻越光为供体亲本构建的56个CSSL株系对WA、CRE和VE 3个稻米蒸煮特性进行QTL定位及感官食味品质相关性进行研究。共检测到4个蒸煮特性QTL,分别为qWA-8、qCRE-8、qCRE-11和qVE-11,其中qCRE-11与qVE-11为新鉴定的QTL。qVE-11的LOD值和贡献率最高分别为6.33和41.16%,推测可能为1个主效位点。对感官食味品质性状与蒸煮特性进行相关性分析发现,感官食味品质与蒸煮特性之间无相关性,稻米蒸煮特性WA、CRE与VE之间两两呈极显著正相关;稻米感官食味品质性状AP、ARM、TA、TE和EQ之间两两呈极显著正相关。本研究利用一套籼粳杂交置换系分析稻米蒸煮特性与感官食味品质之间的关系,定位稻米蒸煮特性QTL,以期为揭示稻米蒸煮特性与感官食味品质的遗传调控机制和遗传改良提供理论依据。
