遮阴对景东翅子树幼苗光合特性和叶绿素荧光参数的影响*
2022-02-19王博轶王齐吴训锋刘强邢海涛
王博轶,王齐,吴训锋,刘强,邢海涛
(云南林业职业技术学院, 云南 昆明 650224)
光是影响植物生长及种群更新的重要生态因子,对植物的生长、发育和形态建成具有极其重要的作用[1]。光合作用是植物生长的重要前提,不同光环境对植物的光合作用和生产力都有重要的影响。一般而言,喜阳植物适宜于强光下生长,喜阴植物适于弱光下生长。遮阴通过降低叶面温度和土壤温度,提高空气湿度和土壤中水分含量等,进而影响植物的光合作用[2]。植物种类不同,需光特性也不同,随光环境的改变,不同种类植物光合特性的响应及适应能力也存在差异。这些往往能反映植物在生长发育过程中对不同生态因子的需求,从而为植物的人工繁育、引种栽培以及进一步的开发利用提供依据[3]。近年来,光环境影响濒危植物更新的研究,也为濒危植物的生境保护及恢复提供了科学的参考依据[4-5]。
李冬林等[4]通过遮光对连香树(Cercidiphyllumjaponicum)幼苗光合特性及其叶片解剖结构影响的研究发现,连香树具有一定的光忍耐性和喜光性,对光照条件的生态幅较宽,但强度遮光对连香树幼苗气体交换参数和光合响应特征产生了显著影响。李小琴等[5]研究了遮阴对濒危植物风吹楠(Horsfieldiaglabra)幼苗叶形态和光合参数的影响,发现遮阴处理使风吹楠的最大净光合速率、表观量子效率、光饱和点显著降低,光补偿点和暗呼吸速率也相应改变,强光照下风吹楠并未发生明显光抑制和“午休现象”。表明光照充足的环境适合其正常生长,较大程度的遮阴使幼苗生长严重受阻。所以,研究濒危树种幼苗期对光照的需求和适应性,有助于了解植物的耐阴性,探索其保护措施以及如何提高幼苗的竞争力,以促进其野生种群的自然更新和演替。
景东翅子树(PterospermumkingtungenseC.Y.Wu ex Hsue)为梧桐科(Sterculiaceae)翅子树属(Pterospermum)植物,是中国滇中地区石灰岩山地常绿阔叶林中的特有种,属国家Ⅱ级保护的极度濒危物种[6]。在该植物的本底调查中,发现景东县境内共分布有景东翅子树约2 432株,分为9个居群,涉及5个乡镇。景东翅子树仅分布于景东县境内河谷沿线石灰岩地区,海拔1 440~1 530 m,物种对生境的选择十分严格,且成年植株较少,自然结实率低。按IUCN濒危等级标准评价属于“极危种(CR)”。景东翅子树大多分布在保护区以外,绝大多数分布在社区村民的山林内(集体林或自留山)、农地旁,或者县级主干道公路的沿线,常遭人砍伐当作薪柴,或受到公路维修的影响或破坏[7]。目前,关于景东翅子树的研究仅限于种群分布、年龄结构[7],以及扦插生根等方面的报道[8]。而对其光合特性等方面的研究尚未见报道。本研究通过不同遮阴处理,从叶绿素荧光特性和光合生理特性等方面研究景东翅子树幼苗对不同光照条件的响应机制,以期为景东翅子树的濒危原因和种质资源保护提供参考。
1 材料与方法
1.1 试验地概况
试验在云南林业职业技术学院(25°02′11″N,102°42′31″E,海拔约2 000 m)进行。该地属北亚热带至中亚热带半湿润山地气候,年均温14.5 ℃,极端最高温33.1 ℃,极端最低温-9.2 ℃;年均降水量1 008.9 mm,一年中有明显的干季(11月—翌年4月)和雨季(5—10月)之分,降雨多集中在雨季,约占全年降水量89%。土壤以山地红壤为主。
1.2 试验材料与设计
选择长势基本一致且生长健壮的景东翅子树1 a生实生苗(苗高45 cm,地径5 mm),于2019年3月定植,经过3个月缓苗生长,当年6月置于学院试验大棚进行遮光处理。不同遮阴处理采用黑色遮阳网进行,共4个处理:3个阴棚内的相对光强分别为自然光强的50%(大林窗中的光强,L1)、36%(小林窗中的光强,L2)以及12.5%(林下的光强,L3),以及不加遮阳网,即全光照处理(对照组,CK)。各处理下设置3盆幼苗,容器上口径19 cm,下口径16 cm,高20 cm,栽培基质一致。苗期常规管理,确保不同处理间管护方式一致。试验处理5个月后,于2019年11月对其光合参数及叶绿素荧光参数进行测定。
1.3 指标测定
1.3.1 叶绿素荧光参数的测定
用FMS2.01 型脉冲调制荧光仪(Hansatech公司,英国)于晴天测定不同光处理下光系统Ⅱ(PSⅡ)最大光能转换效率(Fv/Fm)和非光化学猝灭系数(NPQ)的日变化。每株选取向阳中部正常生长的成熟叶片,叶位一致,叶柄标记。第一次测定时间为上午7:00,之后每隔2 h测定1次,直至当天19:00,每次测定前对叶片测定部位充分暗适应30 min。以Fv/Fm为指标判断光抑制是否发生,以NPQ为指标判断植物热耗散的多少[9-10]。
1.3.2 光响应曲线的测定
用Li-6400便携式光合作用测定仪(Li-Cor, Lincoln, 美国)进行光响应曲线(Pn-PAR)的测定,使用光合仪配备LED红蓝光源,测定时间为上午9:00—11:30,重复测定3次。测定时使用开放气路,空气流速为0.5 L/min,温度25 ℃,相对湿度40%~50%,CO2浓度360 μmol/mol。光合有效辐射(PAR)为2 000、1 500、1 000、800、600、400、200、100、50和0 μmol/(m2·s),每个光强下稳定3 min后记录数据。测定前叶片在800 μmol/(m2· s)光强下诱导30 min。Pn-PAR曲线拟合采用非直角双曲线修正模型[11],其模型表达式为,
Pn=α (1-βPAR)/1+γPAR·PAR-Rd
式中:α为光响应曲线的初始斜率;β和γ为系数,Rd为暗呼吸。
计算出最大净光合速率(Pmax)、暗呼吸速率(Rd)和表观量子效率(AQY)。光补偿点(LCP)和光饱和点(LSP)利用低光条件下(PAR≤200 μmol/(m2·s)的Pn与PAR构建线性方程进行估算:
Pn= mPAR+ n,LCP=- n/m,LSP=(Pmax-n)/m。
1.3.3 光合日变化的测定
选择连续晴朗无风天气,所选取样叶与用于叶绿素荧光参数日变化测定的叶片为相同叶片。利用Li-6400便携式光合系统从8:00—18:00,每隔2 h 测定1 次叶片的气体交换特性,连续测定3 d,测定时均使用自然光源,标准叶室。自动记录PAR、气温(Ta)、大气相对湿度(RH)、CO2浓度(Ca)等环境参数,测定指标为净光合速率(Pn,μmol·m-2·s-1)。
1.3.4 叶片气体交换参数的测定
用Li-6400便携式光合作用测定仪进行气体交换参数的测定,测定时间与各参数设置与光响应曲线一致。光强设为1 000μ mol/(m2· s),叶片选择方法同上,测定参数包括蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)。
1.4 数据处理
所有数据采用统计软件IBM SPSS statistics 20进行分析,处理间比较采用方差分析和LSD检验。统计图采用Sigmaplot 12.5绘制。
2 结果与分析
2.1 不同生长环境光强对景东翅子树叶绿素荧光参数的影响
不同生长环境光强下,其光合有效辐射值的变化基本相同,8:00开始随时间的增加逐步增加,于12:00—14:00达到最高,之后下降(图1)。

图1 不同遮光条件下光合有效辐射(PAR)的日变化Fig.1 The diurnal variation of photosynthetically active radiation(PAR)under different shading treatment
一天当中各种不同遮光条件下,黎明Fv/Fm最高,随日间光强的升高,Fv/Fm降低,下午(14:00左右)光最强时Fv/Fm降到最低点,之后随光强的减弱缓慢回升,19:00光照强度较弱时,各光强下Fv/Fm仍未恢复到天亮前的水平,表明在日间不同光强下均发生了光抑制,全光照CK处理下光抑制最严重,在下午14:00左右Fv/Fm只有0.68左右(图2)。L1和L2处理下,下午14:00Fv/Fm最低,分别为0.72和0.76左右。

图2 不同光强下生长的景东翅子树幼苗光系统Ⅱ最大光能转换效率(Fv/Fm)的日变化Fig.2 The diurnal variation of maximum efficiency of PSⅡ photochemistry(Fv/Fm)for seedlings of P.kingtungense grown under different light regimes
不同光强下生长的景东翅子树幼苗非光化学猝灭系数NPQ的日变化表明热耗散随日间光强的增大而增多。全光照(CK)下的热耗散明显高于遮阴下L1、L2和L3处理的值(图3)。

图3 不同光强下生长的景东翅子树幼苗非光化学猝灭系数(NPQ)的日变化Fig.3 The diurnal variation of non-photochemical quenching efficiency(NPQ)for seedlings of P.kingtungense grown under different light regimes
2.2 不同生长光强下景东翅子树净光合速率(Pn)的日变化
从图4可以看出,景东翅子树幼苗Pn日变化曲线在不同光强下呈现出不同的类型。全自然光(CK)和50%自然光(L1)下,日变化呈“双峰”曲线。CK处理下,上午10:00左右出现第一个Pn高峰值,中午12:00左右出现光抑制现象,Pn只有0.8 μmol/(m2·s)左右,之后Pn回升,但幅度不大,到下午16:00左右出现第二个高峰值,随后下降,但峰值远低于第一个峰值。可以看出全光照CK处理下,景东翅子树幼苗受到了严重的光抑制。在L1处理下的变化与CK类似,但其变化较为缓和,在中午13:00点左右出现光抑制现象。 L2、 L3处理下均呈现“单峰”曲线,略呈“几”字形变化,Pn高峰值出现在上午11:00左右。
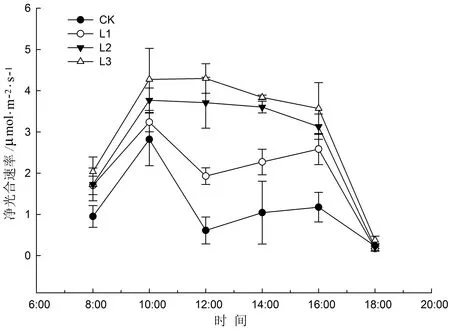
图4 不同光强下生长的景东翅子树幼苗净光合速率(Pn)的日变化Fig.4 The diurnal variation of net photosynthetic rates (Pn)for seedlings of P.kingtungense grown under different light regimes
2.3 不同生长光强对景东翅子树幼苗光响应曲线及其参数的影响
不同光强下生长的景东翅子树幼苗的光合-光响应(Pn-PAR)曲线(图5)显示,当光合有效辐射PAR≤ 200 μmol/(m2·s)时,各光强处理下的Pn均随PAR的增加而迅速增加,L2、L3处理下Pn上升速度比全光照CK处理下要快,说明对弱光的利用效率较高。各光处理下,当PAR升高至400~600 μmol/(m2·s)后,Pn逐渐趋于稳定,呈现光饱和状态。L2、L3处理下,在PAR增至1 600 μmol/(m2·s)时,Pn有明显下降,说明遮阴处理下景东翅子树幼苗Pn易受强光抑制(表1)。

图5 不同光强下生长的景东翅子树幼苗的光合-光响应(Pn-PAR)曲线Fig.5 Pn -PAR response curves for seedlings of P. kingtungense grown under different light regimes
从表1可知,遮阴处理显著提高了景东翅子树的Pmax,L3处理下Pmax比CK、L1、L2处理下分别增长了45%,34%和8%。但所有遮光处理下,景东翅子树的Pmax都比较低,最高值仅为4 μmol/(m2·s)左右,这可能也是景东翅子树生长缓慢,植株自我恢复能力差的一个原因。随遮光率的增加AQY升高,L2处理下AQY达到最高,L3处理下又有所下降。另外,光饱和点(LSP)在各光强处理之间无显著性差异。L2和L3处理下光补偿点(LCP)差异不大,但是明显比CK和L1处理下的LCP要低。暗呼吸速率(Rd)在L2处理下最低,L3遮光条件下有回升,但均明显低于CK和L1处理光强下的值。

表1 不同相对光强下生长的景东翅子树幼苗光响应曲线的特征参数Tab.1 Photosynthetic parameters of Pn -PAR response curves of Pterospermum kingtungense seedlings grown under different light regimes
从表2可知,CK处理下气孔导度(Gs)明显低于各遮阴处理下的值,L3处理下Gs最高。L2和L3处理下的胞间CO2浓度(Ci)差异不大,但是明显比CK和L1处理下的Ci要低。蒸腾速率(Tr)也有同样的变化趋势,L2和L3处理下的Tr没有差异,但均明显高于CK和L1处理光强下的值。

表2 不同相对光强下生长的景东翅子树幼苗的气体交换参数Tab.2 Gas exchange parameters of P.kingtungense seedlings grown under different light regimes
3 讨论与结论
植物在长期适应不同光环境的过程中,不同植物所接受的太阳辐射量存在明显差异,因此形成了自身特有的需光特性[12-14]。喜光植物喜欢在强光环境下生长,如山黄麻(Tremaorientalis)[15]、风吹楠[5]等;而喜阴植物适宜生长在低光强的林下环境,如玉蕊(Barringtoniamacrostachya)[16],谢君魔芋(Amorphophallusxiei)[17]等。
荧光参数Fv/Fm反映了PSⅡ反应中心光能转化效率,其下降是光抑制的重要特征。在未遭受光抑制并经过充分暗适应的植物叶片的Fv/Fm值一般介于0.80~0.83[18]。本研究中,只有在L2和L3处理下,早上8:00以前的Fv/Fm在0.80~0.82左右,其余任何光照处理下的任何时段均低于0.80,叶片光合作用均受到了光抑制。尤其在全光照CK处理下,14:00左右Fv/Fm值仅为0.68,光抑制程度最高。除L3处理,18:00之后Fv/Fm可基本恢复至0.80水平,其余光照条件下,直至晚上19:00均在0.78以下,未恢复至天亮前的水平,光抑制明显。非光化学猝灭系数(NPQ)反映了叶片对激发能利用的情况,尤其是热耗散可以防御光抑制的破坏,是植物保护PSⅡ的重要机制[9]。景东翅子树叶片热耗散随光照强度的升高显著增多,CK处理下其NPQ值达4左右。全光下热耗散的增加可以有效地保护光合机构免受强光的破坏[9,19]。
表观量子效率(AQY)是光合作用中光能转化效率的一种量度,代表叶片对弱光的利用能力,其值越大,叶片吸收与转换光能的色素蛋白复合体越多,利用弱光的能力也就越强,AQY降低是光合作用光抑制的显著特征之一[9]。通常植物的AQY在0.03~0.05之间[20]。本研究中,全光照CK处理下,景东翅子树AQY为0.02,显然受到了光抑制。随遮光率的增加AQY升高,L2处理下AQY最高0.06,之后L3处理下降低至0.03,仍在正常范围内。说明遮阴条件下,光抑制降低,且其利用弱光的能力增强。这也与梁文斌等[21]和唐星林等[22]的研究结果一致。
光补偿点(LCP)和光饱和点(LSP)代表植物叶片对弱光的利用能力。LCP低,表明植物对弱光的利用能力强,LSP低表明植物随光强增大很快达到最大净光合效率。暗呼吸速率(Rd)反映光照条件下叶片的线粒体呼吸作用大小。本研究中,遮阴处理下,景东翅子树光补偿点(LCP)和光饱和点(LSP)分别在20 μmol/(m2·s)左右和200 μmol/(m2·s)左右,为典型的喜阴植物,LSP低的喜阴植物更容易因光抑制而受伤害[21]。景东翅子树LSP在各光强处理之间无显著性差异,即LSP基本不受遮阴强度的影响。L2和L3处理下的LCP明显比CK和L1处理下的LCP值要低。而且,Rd在L2遮光条件下最低,虽L3处理下有所回升,但两者均明显低于CK和L1处理光强下的Rd值。弱光条件下景东翅子树通过降低LCP来提高对弱光的利用能力,通过降低Rd来减少有机物的消耗维持碳平衡,对弱光环境表现出一定的适应性。景东翅子树通过提高AQY,降低LCP和Rd等来适应弱光条件,这与刘柿良等[23]、李珍等[17]、蔡建国等[24]和王佳佳等[25]的研究结果相类似。
在CK与L1条件下,景东翅子树幼苗叶片均出现光合午休现象,而在L2和L3处理下却未出现此现象,说明在CK与L1处理下,景东翅子树叶片发生了光抑制现象。遮阴处理提高了景东翅子树的最大净光合速率Pmax,但所有光处理下景东翅子树的Pmax都比较低,最高值仅为4 μmol/(m2·s)左右,这可能也是景东翅子树生长缓慢,种群自我恢复能力差的一个原因。Gs可以反映叶片内CO2扩散阻力对光合速率的限制。本研究中,景东翅子树叶片CK处理下Gs最低,高光强影响了叶片的气体交换能力,而L3处理下Gs最大,说明遮阴条件下景东翅子树叶肉细胞内CO2的扩散阻力较小,光合速率增大。L2和L3处理下的Ci和Tr均与CK和L1处理下的值差异显著。可以看出,遮阴条件下,景东翅子树Ci降低,并伴随较高的Tr,Ci降低说明叶片固碳的能力提高。景东翅子树在遮阴后,叶片会通过增加Tr和提高CO2同化力来提高其光合速率。进一步证实弱光环境反而有利于景东翅子树增强其光合作用能力。
本研究结果可以看出,强光下景东翅子树幼苗对光抑制敏感,其通过增加热耗散,消耗掉过剩光能,在一定程度上保护了光合机构免受强光的破坏。景东翅子树有较低的Pmax、LSP、LCP和Rd等,以适应弱光环境。总之,由其光合特征与叶绿素荧光特性来看,景东翅子树幼苗是一种典型的喜阴植物,在育苗期进行适当地遮阴处理有利于其生长发育。
