外源硫化氢对成龄桃树根系生长的影响
2022-02-08高晓兰孙茂祥吴雪莲肖元松彭福田
高晓兰,孙茂祥,吴雪莲,肖元松,彭福田
(1.山东省果树研究所,山东 泰安 271000;2.山东农业大学园艺科学与工程学院,山东 泰安 271018)
桃树根系主要分布在表层土壤中,以水平生长为主,没有明显的主根,同级根系分枝粗细相近。桃树根系的水平分布范围一般与树冠冠径相近或略广,垂直分布大部分根系在1 m以内。但桃树根系的分布受环境条件的影响较大,在不同环境中表现出明显的差异。
硫化氢(H2S)是继一氧化氮(NO)和一氧化碳(CO)之后第3个被发现的气体信号分子[1],参与干旱胁迫下十字花科植物光合作用的调节[2],而且可与植物激素交叉调控种子的萌发[3]。近年研究发现,硫化氢能够调控植物根系的生长发育。李东波等[4]发现低浓度外源硫化氢可以提高根尖边缘细胞存活率、根尖组织可溶性蛋白含量,进而促进豌豆胚根生长。硫化氢预处理通过减少根系ROS的积累和保护质膜完整性,减少根尖细胞的死亡来缓解Cd胁迫对根长的抑制,减少由ROS积累引起的细胞死亡[5]。Li等[6]研究表明硫化氢可与生长素(auxin,IAA)相互作用,调节甘薯和番茄植株的不定根和侧根形成。硫化氢不仅对根系的伸长具有剂量效应,而且与植物侧根的发生相关[7]。Lin等[8]研究发现,NaHS通过CO产生酶血红素加氧酶-1(haem oxygenase-1,HO-1)诱导黄瓜不定根的发生。Fang等[9]发现硫化氢参与生长素诱导的番茄侧根形成。Jia等[10]发现硫化氢通过调节ABP调节肌动蛋白依赖性生长素运输引起拟南芥中根发育的改变。Mei等[7]研究表明,硫化氢通过过氧化氢诱导番茄幼苗侧根形成。
桃树根系生长状况对养分和水分的吸收利用至关重要,硫化氢参与调控植物生长发育的很多生命过程。笔者前期研究发现,外源硫化氢可促进桃实生苗根系的生长[11]。但在大田条件下,硫化氢对成龄桃树根系生长的作用效果尚不明确。为此,本试验以硫化氢对根系生长的调控作用为切入点,在前期试验的基础上,以7年生‘瑞光33/毛桃’为试材,研究外源硫化氢对成龄桃树根系生长的影响,同时利用实时荧光定量PCR技术,检测外源硫化氢对桃实生苗根系生长素合成编码基因YUC1和YUC2及生长素转运编码基因AUX1、PIN1、PIN2和PIN3的表达水平,探讨硫化氢调控桃树生长发育的作用效果,最终明确外源硫化氢对成龄桃树根系生长的调控作用,以期为桃树栽培生理研究提供有益参考。
1 材料与方法
1.1 外源硫化氢对成龄桃树根系生长的影响试验
1.1.1 试验设计与处理方法 于2018年6月至10月在大田条件下进行。在前期研究筛选出硫化氢供体NaHS适宜浓度为0.2 mmol·L-1的基础上,以7年生‘瑞光33/毛桃’为试材,供试土壤为粘壤,设置2个处理:0.2 mmol·L-1NaHS(NaHS)、清水对照(CK)。每隔15天处理一次,共处理5次,每处理7次重复,单株小区。每次使用0.2 mmol·L-1的NaHS(对照用3 L清水)3 L浇灌在树干两侧30 cm处选取的40 cm见方的土壤中,为防止水分和释放的硫化氢散失,处理后用聚乙烯薄膜覆盖处理位置的土壤表面。
1.1.2 测定指标与方法 处理结束后进行破坏性取样,取出整个根系,冲洗干净,使用专业版WinRHIZO根系分析系统测定根系构型参数,包括总表面积(cm2)、根尖数、总体积(cm3)、分枝数、交叉数、根系平均直径(cm)、侧根数量、根系总长度(cm)。每处理重复3次,结果取平均值。
测定结束后,根系按直径大小进行分级并解析(<2 mm为细根,2~5 mm为中等粗根,>5 mm为粗根),置于烘箱中烘干,测定各类根系的干重,每处理3次重复,结果取平均值。
1.2 外源硫化氢对桃根系生长素合成及转运编码基因表达的影响试验
1.2.1 试验设计与处理方法 为了研究外源硫化氢对桃根系生长素合成转运及细胞周转蛋白编码基因的影响,以长势基本一致、无病害的5片叶毛桃实生苗为试验材料,幼苗定植于盆中,盆为圆柱形,内径24 cm,高12.8 cm,盆中基质为石英砂。设置CK(每盆浇施100 mL的清水,对照)、NaHS(每盆浇施100 mL 0.2 mmol·L-1的硫氢化钠)、HT(每盆浇施100 mL 0.2 mmol·L-1的硫化氢清除剂亚牛磺酸)3个处理,每处理30株。处理2h后,将各处理根系取样清洗干净后,放置在-80℃冰箱中备用。利用实时荧光定量PCR技术测定根系生长素合成编码基因YUC1和YUC2及生长素转运编码基因AUX1、PIN1、PIN2和PIN3的表达水平。
1.2.2 实时荧光定量检测 采用EASYspin Plus植物RNA快速提取试剂盒(北京爱德莱生物科技有限公司)提取样品RNA,利用反转录试剂盒(Perfect Real Time,TaKaRa)获得cDNA。实时荧光定量PCR采用SYBR Green PCR PremixEx Taq(宝生物公司),操作参照说明书。使用CFX96 Touch Real-Time PCR Detection System(Bio-Rad,USA)进行PCR反应。反应程序为:95℃预变性30 s;95℃变性5 s,退火30 s(退火温度根据设计的引物温度而定),退火延伸65℃5 s,40个循环。所有反应设3次技术学重复,所得数据采用2-ΔΔCt进行计算分析。引物参见表1。

表1 引物信息
1.3 数据分析
试验数据采用Microsoft Excel进行图表绘制,采用DPS软件对数据进行差异显著性检验(Duncan’s新复极差法,P<0.05)。采用Beacon Designer 7软件进行荧光定量引物设计。
2 结果与分析
2.1 外源硫化氢对成龄桃树根系表型性状的影响
2.1.1 对根系表型结构参数的影响 由表2可以看出,外源硫化氢可在一定程度上促进成龄桃树根系的生长。与CK相比,外源硫化氢处理成龄桃树的根系总长度、根系总表面积、根系总体积、根尖数、分枝数和交叉数分别提高了12.79%、19.26%、12.86%、12.96%、10.83%和25.28%,差异显著;根系平均直径降低了17.46%,差异显著,反映了硫化氢处理促进了成龄桃树细根的发生。并且,硫化氢处理显著提高了成龄桃树的根系活力(图1)。
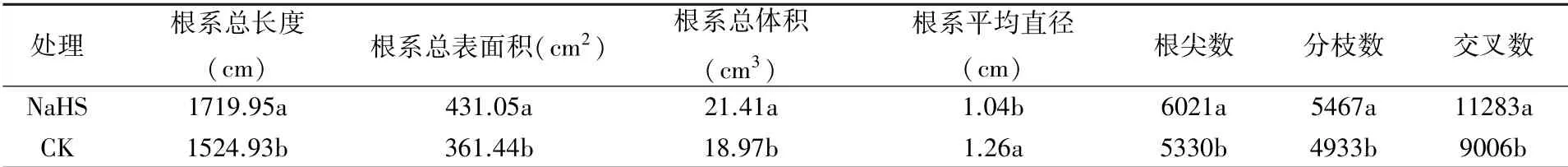
表2 外源硫化氢对成龄桃树根系生长的影响

图1 外源硫化氢对成龄桃树根系生长活力的影响
2.1.2 对不同粗度根表型性状的影响 与对照相比,外源硫化氢处理显著提高成龄桃树直径0~2.0 mm的细根长度、表面积、体积,分别提高了15.54%、27.88%、33.52%,差异显著,但对直径2.0~4.0 mm的中粗根和>4.0 mm的粗根长度、表面积、体积影响不显著(图2~图4)。

图2 外源硫化氢对成龄桃树不同直径根系长度的影响
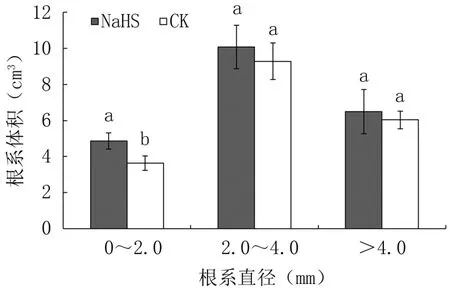
图4 外源硫化氢对成龄桃树不同直径根系体积的影响
2.1.3 对根系分形维数的影响 外源硫化氢处理后成龄桃树根系分形维数较对照提高了2.89%,差异显著(图5)。表明外源硫化氢处理可使成龄桃树根系复杂程度变大。

图3 外源硫化氢对成龄桃树不同直径根系表面积的影响

图5 外源硫化氢对成龄桃树根系分形维数的影响
2.2 外源硫化氢对成龄桃树根系干物质积累量的影响
由图6可以看出,外源硫化氢处理成龄桃树直径0~2.0 mm的细根干物质积累量与对照相比显著提高36.22%;但对直径2.0~4.0 mm的中粗根和>4.0 mm的粗根的干物质积累量影响不显著。可见,在成龄桃树上施用外源硫化氢当年主要促进直径0~2.0 mm细根的生长。
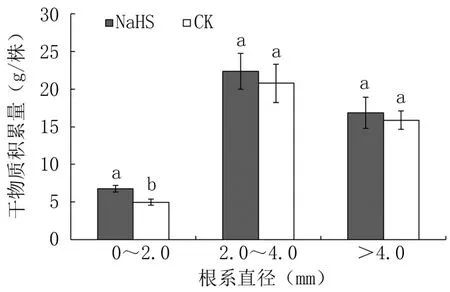
图6 外源硫化氢对成龄桃树根系干物质积累量的影响
2.3 外源硫化氢对桃根系生长素合成及转运编码基因表达的影响
外源硫化氢处理桃实生苗2 h后,根系生长素合成编码基因YUC1和YUC2上调表达(图7);根系生长素转运编码基因AUX1、PIN1、PIN2、PIN3的表达量也上调,尤其PIN1上调表达幅度较高(图8)。表明外源硫化氢可以通过影响根系生长素合成和转运编码基因的表达来影响桃实生苗根系的生长。

图7 外源硫化氢对桃根系生长素合成编码基因表达水平的影响

图8 外源硫化氢对桃根系生长素转运编码基因表达水平的影响
3 讨论与结论
李东波等[4]发现用低浓度外源硫化氢处理豌豆能够使根尖组织可溶性蛋白含量升高,根尖边缘细胞存活率上升,粘胶层相对面积变小,从而促进豌豆胚根生长。本研究结果也表明,施用0.2 mmol·L-1NaHS可显著提高大田条件下7年生‘瑞光33/毛桃’成龄桃树根系总长度、根系总表面积、根系总体积、根尖数、分枝树和交叉数,与对照相比分别提高12.79%、19.26%、12.86%、12.96%、10.83%和25.28%,根系平均直径降低了17.46%,差异均达显著水平;尤其对细根生长促进明显,能使直径0~2.0 mm细根的长度、表面积、体积和干物质积累量分别显著提高15.54%、27.88%、33.52%和36.22%;外源硫化氢处理后成龄桃树根系分形维数较对照显著提高2.89%。由于本试验只进行了一季,时间较短,对根系的长期影响效果可能还未完全显现,且大田条件下成龄桃树根系生长对NaHS的敏感性要小于实验室条件,因此今后有必要对外源硫化氢影响成龄桃树根系生长的长期效果进行深入研究。
Zhang等[12]研究发现NaHS处理能够诱导切根番薯的不定根发生,而加入生长素转运抑制剂和NO清除剂能够逆转IAA和INO对不定根的诱导作用,推测硫化氢可能通过IAA和NO来诱导植物不定根的发生。PIN蛋白是生长素运输所必需的,PIN2蛋白是IAA向基运输的关键调节因子,PIN1蛋白调控IAA的向顶运输。PIN1和PIN2都对根系构型的形成具有重要作用[13]。在PIN系列中,PIN1、PIN3、PIN4和PIN7调节植物侧根的形成[13]。pin1突变体和pin3pin4pin7三重突变体表现出侧根弯曲程度明显减弱[14,15]。有研究表明,当拟南芥根系内源性硫化氢水平升高时,PIN1、PIN2和PIN7的表达增强[10]。另外,控制细胞周期进展的下游分子有助于拟南芥的侧根发生[16,17]。在水稻中,细胞周期调节基因ORC3突变破坏细胞周期进程并阻断主根上侧根的发生[18]。并且生长素可通过调节根系外周细胞中细胞周期调控基因如CYCA2;1、CYCA3;1、CYCD3;1和CDKA1的转录调控侧根的形成[19,20]。
本研究发现,外源硫化氢处理桃实生苗后,根系中生长素合成编码基因YUC1、YUC2及生长素转运编码基因AUX1、PIN1、PIN2、PIN3均上调表达,表明外源硫化氢可能通过影响桃根系中生长素的合成和转运来调控根系生长,但其具体调控机制还有待进一步研究。
