1996—2015年滇西北香格里拉植被净初级生产力变化
2022-01-28栗忠飞王小莲徐钰涛文林琴黄丽春
栗忠飞,王小莲,徐钰涛,文林琴,黄丽春
1 西南林业大学生态与环境学院, 昆明 650224 2 丽江市林业与草原局,丽江 674100
陆地植被净初级生产力(Net Primary Productivity,NPP)是植物群落在其所处环境条件下生产能力的直接反应,是判定陆地生态系统碳源/汇特征的重要依据[1—4]。气候是全球植被格局最重要的决定因素之一,对植被类型的分布、结构和生态功能具有重要影响[5—7]。通过估算植被NPP可以直观反映出陆地生态系统对气候变化的响应,及其多尺度的相互作用过程[8—10],相关研究对进一步理解气候变化对陆地植被及生态系统碳平衡的影响、以及评价生态系统服务功能质量或恢复保护状况等均具有重要作用[11—12]。
NPP的估算方法包括站点观测法、实验法和模型估算等[13—15]。地面站点观测或实验法的估算结果较贴近实测值,但无法反映较大尺度的NPP及其动态变化,而模型的运用能更好地解决站点观测法的不足[16]。国际上,Lieth首次开展了利用模型对全球植被NPP进行估算的研究[17]。在模型估算中,CASA模型是NPP模拟研究中具代表性的一类模型[18—20],被广泛应用于全球及区域NPP研究中[14,21—24]。CASA模型利用3S技术和遥感数据与近地面的气候数据,能够反演出不同尺度植被NPP及其动态变化[2,10,20,25—26]。但是,因不同植被受水分、温度等因素的影响,CASA模型中,不同植被类型的最大光能利用率均定义为0.389 gC/MJ,对NPP的估算会出现较大误差[27]。
国内学者朱文泉等人,将模型引入植被覆盖分类参数,基于中国植被NPP的实测数据,确定了不同植被覆盖类型的最大光能利用率,实现了CASA模型相关参数的简化与本地化,在相关研究中取得了良好的模拟效果[28—30]。当前,这一改进的CASA模型已经被广泛应用于国内外陆地植被NPP的模拟研究中[21,29,31—32]。
滇西北香格里拉区域,地处青藏高原东南缘及滇西北“三江并流区”,区域内植被覆盖率高,其NPP对气候变化的响应,对于揭示气候变化下青藏高原的响应具有重要科学意义。当前,有关该区域内植被NPP时空格局变化的研究仍然非常稀缺[33—35],个别相关研究也缺少对该区域NPP时序变化特征,以及其对温度、降水等气候变化响应规律的分析。
由于植被覆盖水平、气候条件等的差异,植被NPP与NDVI之间的统计关系在生长季内不同时期存在差异,一般在6—8月份表现最优[36],能够代表年度内最大NPP月份。气候变化下森林生态系统敏感的响应能力有可能导致生产力下降,NPP时空格局发生改变[37—38]。青藏高原东南缘香格里拉区域内分布有大面积的针叶林,是我国高山植被分布的典型区域。本研究运用改进的CASA模型,基于香格里拉区域的基础地理数据、近地面气象数据和MODIS-NDVI数据[39—42],对区域内1996—2015年间,逐年6—8月NPP时序变化进行模拟估算,并分析其时空变化特征。研究旨在揭示香格里拉县近20年来区域植被NPP在温度、降水格局变换下的时空动态规律,为促进滇西北地区的植被保护与建设提供科学依据。
1 研究区概况
云南省香格里拉(99°23′—100°31′E,26°52′—28°45′N)地处青藏高原东南缘、横断山脉腹地,是中国地势三大阶梯中第一、二阶梯的过渡带,具有高海拔、低纬度的地理特征,区域内地形走势为西北高、东南低,境内平均海拔3459 m,地貌类型主要为高原山地、盆地、河谷等。
区域内气候类型属于温带、寒温带季风气候,河谷区属北亚热带季风气候,年平均气温6.3 ℃,年均降雨量651.1 mm,该地区终年太阳辐射强烈。区域内植被类型丰富,尤以针叶林、阔叶林、高山灌丛、高山草甸为主,并呈现典型的垂直带普特征,其中针叶林所占面积最大[34]。区域内土壤类型主要有棕壤、暗棕壤、草甸土等。
2 数据获取及其研究方法
2.1 遥感数据及处理
遥感数据采用NASA对地观测系统数据共享平台(https://ladsweb.modaps.eosdis.nasa.gov/search/)提供的2000—2015逐年6—8月MODIS 1B数据,空间分辨率为250 m、时间分辨率为16 d的MODIS植被指数产品MOD13Q1,经最大值合成法(MVC)得到逐年6—8月NDVI数据[43]。
1996—2000年的NDVI数据采用地理空间数据云(http://www.gscloud.cn/)提供的Landsat 5 TM影像,经辐射定标、大气校正、融合裁剪等预处理后,执行波段运算与叠加,并经最大值合成法(MVC)处理,获得1996—2000逐年6—8月NDVI数据。然后,将前后NDVI数据重采样统一分辨率后逐像元对比,在两种数据源之间按一元线性回归进行序列同化,实现NDVI数据序列的插补展延[44],最终获得全部1996—2015年逐年6—8月NDVI时间序列栅格数据。
植被类型数据采用地理空间数据云提供(http://www.gscloud.cn)的Landsat TM影像数据。首先,进行数据预处理得到研究区域遥感影像;其次,采用监督分类中的支持向量机方法,结合目视解译以及谷歌地图等辅助参考数据作为选取样本依据,得到30 m空间分辨率的研究区土地利用分类图,经过影像解译精度评估,Kappa系数在85%以上,解译效果良好;最后,经重采样后将其转换为与以上NDVI数据相匹配的栅格数据。
2.2 气象数据及处理
气象数据采用中国气象数据网(http://data.cma.gov.cn)提供的包括中甸、德钦、维西等香格里拉研究区域及周边14个气象站点20年间6—8月日平均温度、日降雨量、日照时数及太阳总辐射量等数据。首先,进行精度验证,去除异常值;其次,结合各站点的经纬度信息,基于气象插值软件ANUSPLIN 4.3[45],运用反距离权重法(IDW)对气象数据进行空间插值,得到温度、降水量及总辐射量序列数据,最后通过掩膜研究区,裁剪得到1996—2015年空间分辨率为250 m的气象栅格数据。
2.3 研究方法
2.3.1模型选取
本文采用朱文泉改进的CASA模型估算NPP,NPP由植物光合有效辐射(APAR)和实际光能利用率(ε)两个因子来表示,估算公式及其意义详见文献[22,30,46]。
NPP(x,t)=APAR(x,t)×ε(x,t)
(1)
式中,APAR(x,t)表示像元x在t月吸收的光合有效辐射,MJ/m2;ε(x,t)表示像元x在t月的实际光能利用率,gC/MJ,NPP(x,t)是像元x在t月的净初级生产力。
APAR(x,t)=SOL(x,t)×FPAR(x,t)×0.5
(2)
式中,SOL(x,t)表示t月在像元x上的太阳总辐射量(MJ/m2),FPAR(x,t)植被层对入射的光合有效辐射吸收的比例,常数0.5表示植被利用的光合有效辐射(波长为0.4—0.7 μm)占太阳总辐射的比例。在一定范围内,FPAR与NDVI存在线性关系,所以,可根据NDVI得到对应的FPAR[2]。
ε(x,t)=Tε1(x,t)×Tε2×Wε(x,t)×εmax
(3)
式中,Tε1(x,t)和Tε2(x,t)表示低温和高温对光能利用率(NPP累积)的胁迫作用;Wε(x,t)为水分胁迫影响系数,反映水分条件的影响;εmax是植被在理想条件下的最大光能利用率(gC/MJ)。
NPP的大小由温度、降水量影响光能转化率的下调因子δ来决定[30],其中δ因子(δ=Tε1×Tε2×Wε)表征植被净生产力受气候因素影响的大小。
FPAR采用MOD13Q1产品中NDVI数据反演得出。公式(1)和(3)中,ε表示光能转化率,反映植被把所吸收的光合有效辐射转化为碳的比率,其主要与温度和水分有关。公式(3)中,Tε1和Tε2表示温度对光能转化率的影响;Wε是水分胁迫系数;εmax是理想条件下的最大光能转化率,其值通常介于0.09—2.16 gC/MJ之间。最大光能转化率随着植被类型的不同而发生变化,朱文泉等[28,30]根据不同植被类型调整的最大光能利用率参数,适合中国植被状况,其参数适用全国不同区域研究[40—42]。本文中选取各植被类型的最大光能转化率参数见表1。
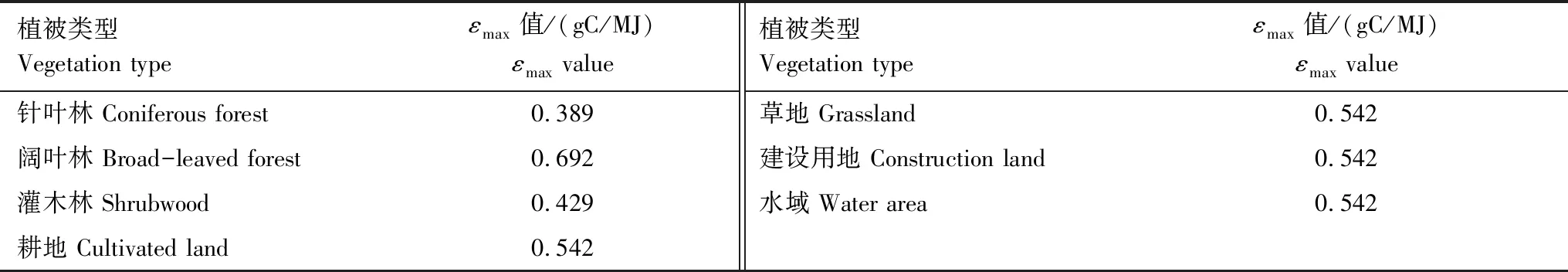
表1 不同植被类型的最大光能利用率εmax
2.3.2NPP年际变化趋势分析
采用一元线性回归法,运用改进的CASA模型[26,28,47],计算得到香格里拉1996—2015年间最大月6—8月不同植被类型的NPP,分析1996—2015年研究区逐年最大月NPP的变化规律,以及与温度、降水等气候因子的相关性规律。
3 结果
3.1 香格里拉区域1996—2015年间气候因子变化
图1显示了1996—2015年香格里拉6—8月期间月平均温度、总降水量及总辐射量变化。月均温在13.2—14.5℃之间波动,平均为13.9℃,20年来气温总体呈升高趋势,上升斜率为0.037℃/a,在横向上呈正态分布,且峰值向右偏移。2009、2013年月均温最高,均达到了14.5℃(图1)。
6—8月总降水量范围在262.2—512.8 mm之间,多年均值为373.1 mm,最大绝对降水距平达到139.8 mm。20多年间6—8月总降水量呈波动下降特征,下降斜率为4.32 mm/a,其中前10年的均值为383.2 mm,后10年的均值为362.9 mm,10年下降幅度为5.3%(图1)。
区域内20年间6—8月太阳总辐射平均值为563.8 MJ/m2,并以此为中心呈现波动平衡状态,从其趋势线的负值斜率上,显现出微弱下降特征(图1)。

图1 1996—2015年香格里拉6—8月气候因子变化特征Fig.1 Time variation curves of climatic factors during 6—8 month from 1996 to 2015 in Shangri-La
3.2 不同植被类型最大月NPP年际变化
图2显示,1996—2015年间,香格里拉地区不同植被类型逐年最大月NPP具有明显差异,阔叶林NPP显著高于其它植被类型,其1996—2015年的平均值达到286.9 gC/m2,最高年份1998年达到347.6 gC/m2,最低年份2004年为201.1 gC/m2;其次为耕地,其20年间平均值为182.1 gC/m2,此后依次为草地、灌丛植被,其20年平均值分别为149.5、137.3 gC/m2。所有植被类型中,针叶林NPP最低,20年均值仅为128.9 gC/m2,显著低于阔叶林NPP(图2)。
从变化趋势看,各植被类型逐年最大月NPP呈微弱增加趋势(图2)。各植被类型平均增加斜率为0.46 gC m-2a-1,其中阔叶林最低,仅为0.02 gC m-2a-1,草地最高,为0.78,针叶林、耕地、灌丛分别为0.69 gC m-2a-1、0.51 gC m-2a-1、0.26 gC m-2a-1(图2)。从前10年,到后10年,各植被类型最大月NPP从171.6 gC/m2增加到了182.3 gC/m2,上升了6.2%(图2)。
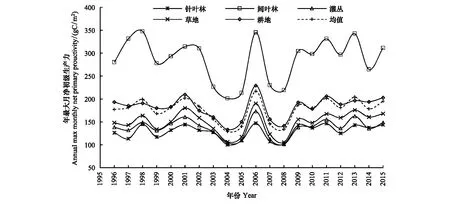
图2 1996—2015年间香格里拉地区各植被类型NPP变化Fig.2 Inter-annual variation of NPP of different vegetation types in Shangri-La from 1996 to 2015
3.3 区域植被NPP空间格局变化特征
图3显示了香格里拉区域,1996—2015年间5个时期的最大月NPP空间分布格局,总体上表现出由中部向四周及两端逐渐增加的趋势。研究区域的南、北两端区域内的最大月NPP多达200 gC/m2以上,区域中部及偏北范围内,其NPP达到了300 gC/m2以上(图3)。图3显示,从1996到2005年期间,香格里拉NPP空间分布总体上呈现下降特征,而从2005年开始到2015年又呈现上升趋势,尤其是在东部,以及西南部区域,变化尤其明显。

图3 1996—2015年香格里拉县植被NPP平均值变化Fig.3 NPP mean change of vegetation in Shangri-La from 1996 to 2015
3.4 香格里拉植被NPP与气候因子的相关性
图4显示,各植被类型最大月NPP与月均温均达到了显著相关水平(P<0.05)。其中,针叶林相关性最高,R2达到0.3474,其次为草地,最低的为灌丛。各植被类型NPP均值与温度的相关性同样达到显著水平(R2=0.2679,P<0.05)。所有植被类型NPP均表现出受到温度正向调控的显著影响(P<0.05)。

图4 不同植被类型NPP与月均温的相关性Fig.4 The correlation between NPP of different vegetation types and mean monthly temperature
图5显示,各植被类型最大月NPP与降水量均未呈现出显著相关性特征(P>0.05)。从其斜率的负向变化来看,多数植被类型均呈现出随降水量增加、NPP微弱下降的特征。
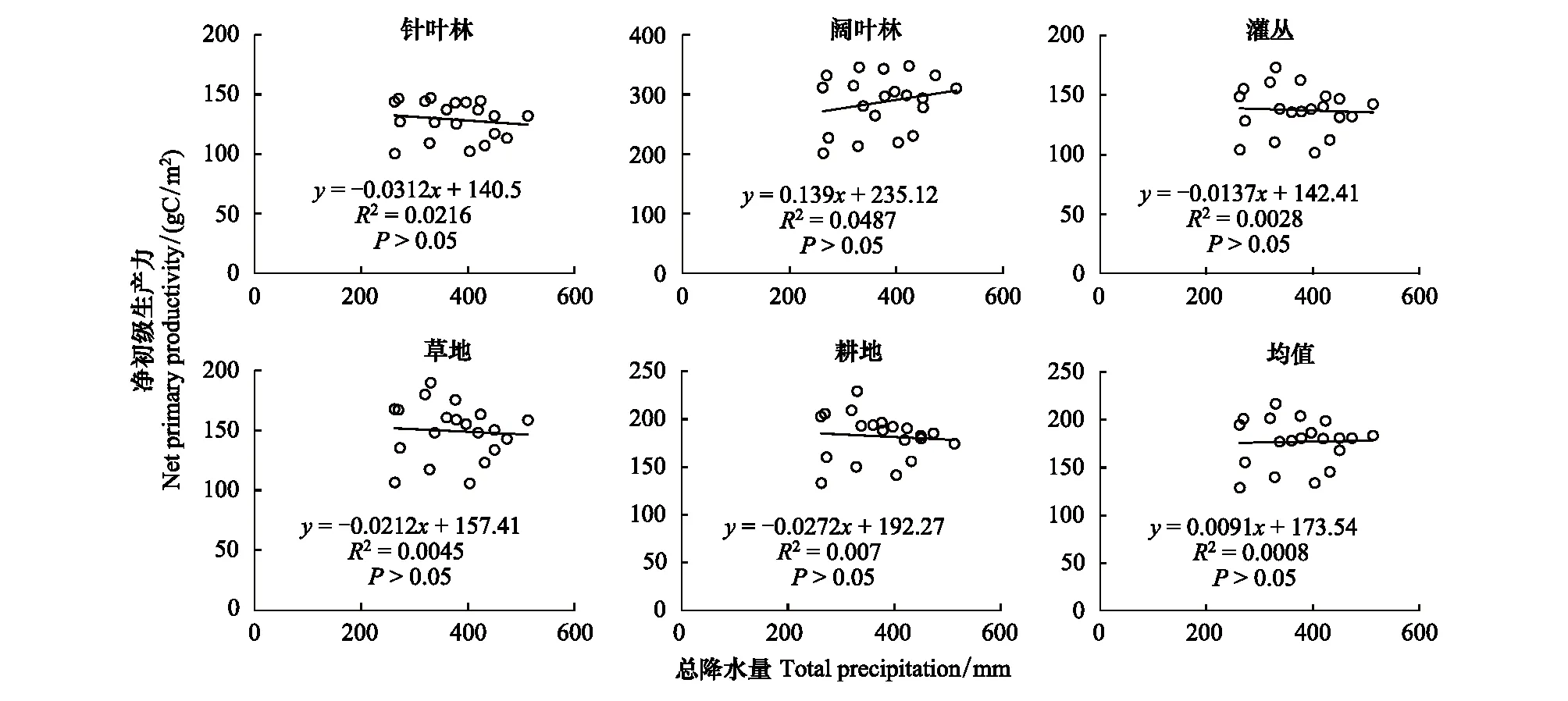
图5 各植被类型NPP与总降水量的相关性Fig.5 The correlation between NPP of different vegetation types and total precipitation
图6显示,相对于温度和降水而言,各植被类型最大月NPP与总辐射量呈现出显著的正相关特征(P<0.01),其R2一致超过了0.4。

图6 各植被类型NPP与总辐射量的相关性Fig.6 The correlation between NPP of different vegetation types and total solar radiation
4 讨论
4.1 基于改进的CASA模型模拟的香格里拉区域最大月NPP
香格里拉一年中的生长季3—10月[35],实质上,受低纬度、高海拔特点的影响,区域内水热分布集中在夏季,因此,6—8月是最为关键的生长季节,也是NPP积累的主要季节,期间,从森林到农田等各植被类型均具有相对较高的NPP(表2)。表2显示了本研究中香格里拉区域20年间各植被类型最大月NPP,与岳彩荣等人[35]对同一区域的模拟结果比较显示,本研究中6—8月最大月NPP占到全年的29.6%—65.9%,占比最大为阔叶林,最小的为针叶林。对比全国尺度上的模拟结果[48—49]),本研究与其基本处于接近的范围。改进的CASA模型,能够较好的适用于本研究区域,并能客观刻画区域各植被类型NPP的时空变化。

表2 香格里拉植被NPP模拟结果的横向比较*/(gC/m2)
4.2 气候因素对香格里拉区域NPP时空格局的影响
NPP作为评定陆地生态系统的重要指标,直接反应区域植被在自然条件下的生产能力[50],而区域温度、降水和辐射等气候因子是绝对自然状态下植被NPP的关键因素[51]。相关研究显示,温度和降水大体上与NPP值呈现正相关特征[52]。本研究中区域内植被NPP与温度的变化显示出显著的相关性特征(P<0.05)(图4),表现出区域受温度调控的强烈影响。但是,研究中,NPP对降水变化的响应却未见正相关性特征,从趋势线来看,反而呈现一定的负相关特征(图5)。植被对温度和降水的气候因子的响应存在地域差异,有相关研究也表明,青藏高原区域内,植被NPP与降水呈负相关特征[53—55]。此外,本研究显示出2005年是区域NPP上升的突变年份(图2),相关研究也显示了类似的结果[56]。表明,区域温度因子在时间序列上的波动变化,对区域内植被NPP的波动变化产生了显著的调控作用,与此同时也体现出,水分条件并未形成对区域植被NPP的限制与调控效应。
本研究中,植被NPP与总辐射量之间极显著的相关性特征(图6),表明了受辐射因子的限制限制性影响作用。本研究的香格里拉区域内,每年从5月份进入雨季后,雨雾天气现象占据主导地位,在每年生长季盛期的太阳总辐射量稀缺,作为CASA模型的关键驱动因素,区域植被NPP的积累直接受了辐射因子的限制与调控,并呈现出一致强烈的正相关作用。
4.3 香格里拉区域植被NPP时空格局及人类活动影响
除上述气候因素外,人类活动也是影响区域植被NPP积累的不可忽略的因素[16]。本研究区域内,人为干扰主要表现为人类聚居区域内,土地利用方式的改变、植被破坏等,以及国家及当地政府长期以来所采取的保护、治理措施。20世纪90年代后期及新世纪初期,地方政府在经济、社会发展的压力下,自然植被资源过渡开发利用,区域内天然植被持续退化,尤其是靠近人类聚居区域的空间范围内,其NPP明显偏低(图3)。随后的10多年里,当地政府及相关部分大力开展的生态保护与建设,以及国家生态文明建设的大力推进[57],很大程度上促进了区域内植被NPP的增进趋势[58]。此外,研究中显示出的针叶林NPP相对较低的特征,表明其一旦破坏、过渡采伐等,将需要较长的持续时间才能恢复,因此,区域内大面积的针叶林更需要进行重点保护[59]。
地处青藏高原东南缘的香格里拉区域,对气候变化的影响更加敏感[60],加之日趋复杂的人类活动影响,区域内植被NPP的变化将存在高度的不确定性,面向区域植被NPP变化的实测研究,以及基于模型的模拟研究,仍需要更为深入的开展。
5 结论
1)1995—2015年间,香格里拉地区6—8月平均气温总体呈上升趋势,增加速率为0.037℃/a;6—8月份月总降水量为373.1 mm,总体呈微弱下降趋势;6—8月总辐射量平均为563.8 MJ/m2,20年间辐射量基本维持波动稳定状态。
2)1996—2015年间,香格里拉区域最大月植被NPP平均值为176.9 gC/m2,不同植被类型NPP的多年平均值位于128.9—286.9 gC/m2之间;空间格局上,表现为“西北部、东南部及金沙江沿岸向中部递减”的空间格局;
3)1996—2015年间,植被NPP显著地受到温度变化的影响(P<0.05),随着20年间温度的持续上升,区域NPP呈现增加趋势;此外,区域植被NPP未表现出受降水量的显著影响(P>0.05),表明水分不是区域植被NPP的限制性因素。
4)人类聚居区域及其周边的NPP相对较低,表明区域内植被NPP受到不同程度人类干扰的影响;分时段看,区域内植被NPP从1996—2005年间呈现下降趋势,2006—2015年间呈现上升趋势,这也显示出香格里拉区域所实施的生态保护建设各类政策措施对植被NPP的促进作用。
