衰老对2种猕猴血液基因表达的影响及与人类的对比
2022-01-26王鑫颀严超超王娇兰月范振鑫四川大学生命科学学院生物资源与生态环境教育部重点实验室成都60065中国科学院成都生物研究所中国科学院山地生态恢复与生物资源利用重点实验室生态恢复与生物多样性保护四川省重点实验室成都6004四川大学生命科学学院四川省濒危野生动物保护生物学重点实验室成都60065
王鑫颀, 严超超, 王娇, 兰月, 3, 范振鑫*(. 四川大学生命科学学院, 生物资源与生态环境教育部重点实验室, 成都60065; . 中国科学院成都生物研究所, 中国科学院山地生态恢复与生物资源利用重点实验室, 生态恢复与生物多样性保护四川省重点实验室, 成都6004; 3. 四川大学生命科学学院, 四川省濒危野生动物保护生物学重点实验室, 成都60065)
衰老是一个系统的过程,表现为逐渐减慢的新陈代谢和细胞器官水平的衰老,并伴随着许多生理指标的变化(Lópezotín.,2013)。年龄是心脏病、癌症和中风等高死亡率疾病的主要风险因素,也是许多慢性病最重要的风险因素(van den Akker.,2014;Peters.,2015)。衰老主要受遗传和环境因素的影响。目前对衰老遗传机制的研究主要来自模式物种和长寿人群,例如和基因的突变被认为与中国汉族人的寿命有关(Li.,2009)。遗传可能只在人类寿命中发挥15%~30%的决定作用,其余的影响则来自环境因素(Ruby.,2018b)。
猕猴属是分布最广的非人灵长类动物,也是人类疾病研究和临床药物开发领域中最重要的动物模型之一(Lan.,2020;Yan.,2020)。老年的猕猴属动物会患上多种人类老年疾病,如心脏病和癌症等(Justice.,2016);普通猕猴(CR)和人类在衰老过程中有许多相似之处,如毛发颜色变浅变薄、身体脂肪重新分布、皮肤和肌肉张力降低以及活力丧失等(Mattison.,2017)。此外,随着年龄增长,CR还会出现糖尿病、肿瘤、肌肉萎缩、骨质流失、免疫功能改变和认知能力下降等症状(Ngwenya.,2015)。对CR的研究还发现,营养限制对延缓衰老的机制在灵长类动物中保守(Mattison.,2012),这种机制可能对人类也同样适用(Colman.,2014;Mattison.,2017;Simpson.,2017)。藏酋猴(TM)是中国特有种,是个体最大的猕猴属物种,也是亚洲最大的猴科Cercopithecidae动物之一(Smith.,2010),是潜在的良好的医学模式动物,其肥胖个体易出现高脂血症等疾病(杨锋等,2010)。
转录组研究发现,基因调控和表达模式的变化在物种进化中起重要作用,而高通量测序技术为研究衰老机制提供了一种新的有效方法(Lan.,2020)。在不同的转录组衰老研究中,只有小部分涉及了与年龄相关的基因集的交叉比较,这无疑增加了构建统一的与年龄相关标记基因的难度。与年龄相关的基因集在不同组织和物种的功能水平上的表达有许多相似之处,这表明通过功能分类研究年龄相关的基因集比分析单个基因更有可能揭示衰老过程的基因表达变化(Zahn.,2006;Passtoors.,2008)。目前猕猴属转录组比较研究没有涉及2个不同的物种年龄相关转录组的对比,而是对比了不同组织间的差异,如Chaves等(2020)对食蟹猴的脑微血管系统、内皮连接介质转录组进行了比较。
本研究收集了本团队之前研究的CRs和TMs随年龄变化的转录组数据(Yan.,2020),系统比较了2种猕猴在年龄增长过程中基因表达的共性和异性,同时比较了这2种猕猴和人类年龄变化相关的血液转录组数据,分析了它们和人类在衰老过程中基因表达的异同,为认识灵长类物种在衰老过程中的基因表达变化特征提供数据。
2 材料和方法
2.1 数据采集与预处理
CRs和TMs血液组织转录组数据共52个样本(7雌21雄CRs和12雌12雄TMs):CRs年龄为1.5~20岁:1.5~5岁7个、6~10岁10个、11~15岁6个、16~20岁5个;TMs年龄为3~18岁:3岁6个、5岁6个、8岁3个、10岁2个、11岁 1个、15岁4个、17岁1个、18岁1个。
下载与CR和TM的年龄相关差异表达基因(differential expressed genes,DEGs),其中,CR 2 138个(阈值:<0.05;933个上调和1 205个下调),TM 2 523个[假发现率(false positive rate,FDR)<0.05;1 358个上调和1 165个下调]。此外,从人类血液组织转录组研究(Peters.,2015)(共14 983个样本)中下载得到了1 497个(600个上调和897个下调)与人类年龄相关的DEGs(阈值:<2.2E-5)。在上述3项研究的差异表达分析中,年龄均被视为连续变量。
2.2 年龄相关DEGs的种间比较
使用VennDiagram包对基因集进行比较并绘制Venn图(Chen,2018)。由于同源基因在各物种中拥有的基因名不同引起了误差的产生(Durinck.,2005),所以在对人类DEGs与2种猕猴进行比较时,将人类基因提交至Ensembl BioMart与猕猴基因进行一对一直系同源基因的同源筛选。
2.3 差异表达基因的富集分析
使用g:Profiler(Raudvere.,2019)对DEGs做GO功能和KEGG通路富集分析。在Significance threshold项选择Benjamini-Hochberg FDR,其他选项为默认,并最终选择FDR<0.05的GO和KEGG富集结果。使用R包ggplot2(Wickham,2011)绘制GO富集结果气泡图和KEGG富集结果条形图。
3 结果
3.1 衰老对TMs和CRs基因表达的影响
341个共同上调和389个共同下调的DEGs与2种猕猴的年龄相关(图1)。而3个DEGs(、、ENSMMUG00000030508)在TM中下调,在CR中上调。
对2种猕猴年龄相关DEGs分别做了GO和KEGG富集分析。TMs的上调DEGs富集在能量代谢途径、自噬途径等相关条目中,如GTPase活性的调控(GO:0036211)、利用自噬机制的过程(GO:0061919)(图2:左)。而TMs的下调DEGs富集在翻译和物质合成代谢的相关条目(图2:右),如翻译(GO:0006412)、肽生物合成过程(GO:0043043)、肽代谢过程(GO:0006518)。CRs的上调DEGs富集在天然免疫途径和凝血等相关条目中,如炎症反应(GO:0006954)、凝血(GO:0007596)等(图3:左)。CRs的下调DEGs富集在翻译、体液免疫等相关条目(图3:右),如基因表达(GO:0010467)、B细胞活化(GO:0042113)。

图1 普通猕猴和藏酋猴上、下调差异表达基因的Venn图Fig. 1 Venn diagram of up- and down-regulated differentially expressed genes (DEGs) in Macacca mulatta and M. thibetana
2种猕猴与年龄相关的DEGs富集结果显示出差异性。在凝血方面,CRs的上调DEGs富集在与凝血相关的生物过程(GO:BP)中,如凝血(GO:0007596),而TMs却没有富集到与凝血相关的GO条目,但TMs上调的DEGs富集到了与凝血相关的KEGG条目,如血小板活化(ko04611)。在能量代谢方面,CRs和TMs上调的DEGs都富集到了GTPase相关途径,如调控GTPase活性(GO:0043087)(图4:右)。但有所区别的是标注了所有CRs和TMs的DEGs的KEGG通路。CRs只在氧化磷酸化(ko00190)的一个调控节点[F-type ATPase (Eukaryotes) c]表现下调,大部分DEGs随着年龄增长而上调。与CRs相反的是,大部分TMs的DEGs在氧化磷酸化途径中随年龄的增长而下调。
2种猕猴DEGs的富集结果也显示出许多相似之处。比如,2种猕猴的下调DEGs都富集到了与蛋白质翻译有关的GO条目,如基因表达(GO:0010467)(图4:右)。这些富集到与蛋白质翻译有关的条目中下调的DEGs表达会影响核糖体的整体功能,这与TMs中下调DEGs富集到的核糖体(ko03010)通路及蛋白质合成过程一致。
2种猕猴的上调DEGs均富集到与细胞自噬相关的GO通路,如囊泡介导的转运(GO:0016192),这与在衰老过程中的细胞代谢减弱所导致的细胞自噬现象一致。此外,CRs上调DEGs富集到的唯一一条KEGG通路是溶酶体(ko04142),与抗衰老密切相关,它同时出现在TMs的上调DEGs富集到的KEGG通路中(图5)。CRs下调基因富集到 2条KEGG通路,分别是原发性免疫缺陷(ko05340)和B细胞受体信号通路(ko04662),其中B细胞受体信号通路也出现在TMs的下调DEGs富集到的KEGG通路中。
在衰老过程中,2种猕猴都经历了免疫上的变化,但也有一些差异。CRs的1 205个下调的DEGs富集在体液免疫相关的GO条目中,并且富集到了B细胞受体信号(ko04662)的KEGG通路。在TMs的B细胞受体信号通路中,则是富集了更多的上调DEGs。通过KEGG富集图(图6)可以发现, 2种猕猴在B细胞受体信号通路中基因表达水平的巨大分歧。大多数CRs的年龄相关DEGs随年龄的增长而表达下调,只在5个节点(PIR-β、SOS、PI3K、CaN、IKKγ)表现为上调。而在TMs中,虽然B细胞信号受体通路的起始阶段(CD72、Igα、Igβ、CD81、CD19等)的基因表达随着年龄增长而有所下降,但由于复杂的原因,通路的中间节点(SOS、PKCβ等)和近末尾节点(CaN、ErK等)的基因表达量总体升高。

图2 藏酋猴的差异表达基因富集到的生物过程GO功能条目(取校正后P值前20)Fig. 2 Top 20 enrichment of biological process-related GO terms of differentially expressed genes (DEGs) in Macacca thibetana
左. 上调DEGs, 右. 下调DEGs
Left. up-regulated DEGs, right. down-regulated DEGs

图3 普通猕猴的差异表达基因富集到的生物过程GO功能条目(取校正后P值前20)Fig. 3 Top 20 enrichment of biological process-related GO terms of differentially expressed genes (DEGs) in Macacca mulatta
左. 上调DEGs, 右. 下调DEGs
Left. up-regulated DEGs, right. down-regulated DEGs

图4 普通猕猴和藏酋猴的差异表达基因共同富集到的GO条目(取校正后P值前15)Fig. 4 Top 15 enrichment of GO terms by the common differentially expressed genes (DEGs) in Macacca mulattaand M. thibetana
左. 上调DEGs, 右. 下调DEGs; 灰色为普通猕猴, 黑色为藏酋猴
Left. up-regulated DEGs, right. down-regulated DEGs; gray presents, black represents
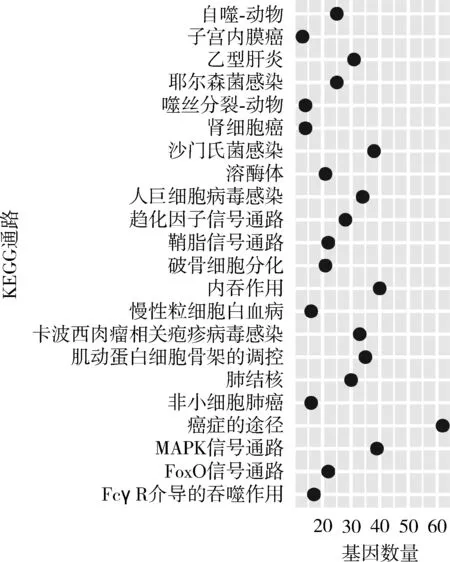
图5 藏酋猴的上调差异表达基因富集到的KEGG通路Fig. 5 Enrichment of KEGG terms by the up-regulated differentially expressed genes of Macacca thibetana
3.2 TMs、CRs与人类年龄相关的DEGs的比较分析
3个物种的年龄相关DEGs一起比较发现,在1 497个人类年龄相关的DEGs中,有309个同时也与TMs的年龄相关,其中133个上调,150个下调。CRs与人类年龄相关的共有DEGs 310个,其中118个上调,174个下调(表1)。三者共有DEGs 127个,如、等,其中23个上调,38个下调。
衰老过程对保守基因表达的影响在不同物种间存在差异。在TMs中,与人类年龄相关DEGs表达方向相反的DEGs包括16个上调(在人类中下调)和10个下调(在人类中上调)。TMs与人类表达方向相反的26个DEGs富集在Th17细胞的分化(ko04659)中,该通路富集了3个与年龄相关的DEGs(、和),其中和的表达随年龄增长而上调。起到抑制通路效果的则是随着年龄增长而下调。在CRs和人类中具有相反趋势的18个年龄相关DEGs并未富集到功能条目,但这些相反的基因也参与到了代谢和延缓衰老等过程,如凋亡调节因子1()、RNA结合基序蛋白11()、真核翻译起始因子2亚基3()。

表1 人类、藏酋猴和普通猕猴的差异表达基因交集统计表Table 1 Intersection statistics of differentially expressed genes (DEGs) in human, Macacca thibetana and M. mulatta
3个物种共同的38个下调DEGs富集到了细胞免疫相关GO通路,比如髓系白细胞介导的免疫(GO:0002444)。3个物种共同的23个上调DEGs富集在与体液免疫密切相关的B细胞受体信号通路(ko04662)。为了更全面地了解3个物种衰老过程中的免疫差别,将所有年龄相关DEGs标注在与体液免疫密切相关的B细胞受体信号通路上。在B细胞受体信号通路中,3个物种在起始阶段(Igα、Igβ、CD72、CD19)的基因表达随着年龄增长下调,人类和TMs在中间节点和近末尾节点(人类:VAV、SHIP、Erk等;TMs:SOS、PI3K、PKCβ、BCL-10等)的基因表达总体上调,而CRs则是下调。从整个通路来说,CRs富集了更多下调DEGs。
4 讨论
4.1 衰老对2种猕猴新陈代谢和免疫力的不同影响
在衰老过程中,CRs和TMs的基因表达的改变有很多相似的地方。2种猕猴的蛋白质翻译功能相关基因表达均显著下调,核糖体的形成过程如核糖体生物发生(GO:0042254)的基因也出现表达下调。这2个过程联系紧密,并造成一系列连锁反应。在核糖体的形成过程中,rRNA不足影响核糖体的合成量;核糖体合成量不足又反过来阻碍蛋白质的翻译过程。翻译是蛋白质代谢的环节,不足的蛋白质代谢影响嘌呤和嘧啶代谢(Rudolph,1994),从而阻碍DNA合成,进而减缓 mRNA 的表达和蛋白质自身产量,并最终表现为机体的代谢率减慢。

图6 人类、藏酋猴和普通猕猴年龄相关差异表达基因富集的KEGG比较通路(B细胞受体信号通路)Fig. 6 Comparison of the B cell receptor signaling pathway enriched by the age-related differentially expressed genes (DEGs) in human, Macacca thibetana and M. mulatta
不同的颜色代表DEGs表达方向的不同; 左边代表人类基因表达水平, 中间代表藏酋猴基因表达水平, 右边代表普通猕猴基因表达水平, 红色表示与年龄正相关DEGs的KEGG同源基因单元, 绿色表示与年龄负相关DEGs的KEGG同源基因单元
Different colors represent different expression directions of DEGs; left part represents the gene expression of human, middle part represents the gene expression of, and right part represents the gene expression of; red represents the KOs including positively age-associated DEGs, green represents the KOs including negatively age-associated DEGs
在2种猕猴随着年龄增长而整体代谢逐渐衰减时,也有部分生物过程被强化,如凝血和自噬相关通路等。个体衰老过程往往伴随着心血管相关疾病(如动脉粥样硬化)的发生(Wilkerson & Sane. 2002)。巨噬细胞是动脉粥样硬化的主要效应细胞(Moore.,2013),2种猕猴的上调DEGs富集到一些巨噬细胞相关的条目:巨噬细胞活化的调控(GO:0043030)、巨噬细胞自噬(GO:0016236,FDR<0.01)。而血栓通常发生在动脉粥样硬化斑块的侵蚀或破裂之后,故巨噬细胞的活跃会影响血栓的形成(Prandoni.,2003)。同时血小板活化通路(ko04611)中增强的凝血功能是血栓形成的风险因子之一,这个通路的功能在TMs中随年龄的增长而增强。推测2种猕猴在衰老过程中血栓的发生率随年龄的增长而增加,这与本团队之前在CRs中的研究一致(Yan.,2020)。
在衰老过程中,2种猕猴的天然免疫能力都显著增强,促进体内形成活跃的炎症环境(Shaw.,2010)。天然免疫系统通过激烈的细胞自噬、吞噬作用等迅速吞噬老旧细胞、病菌等有害物质,是个体对抗衰老、维护健康的手段(Huynh.,2007)。CRs和TMs免疫能力改变的差异表现在以B细胞为主体的体液免疫上。B细胞受体信号通路会随着年龄增长而功能减弱,最终导致B细胞合成减少,表现出免疫老化(Niiro & Clark,2002)。KEGG富集图发现,TMs的B细胞受体信号通路随着年龄的增长而变得更加活跃,表明体液免疫随着年龄的增长而增强(Murphy & Weaver,2012),CRs的情况则相反。以上暗示TMs特异性免疫力下降可能没有CRs明显,CRs的免疫老化现象随着年龄的增长可能更加突出(Asquith.,2012)。
2种猕猴在衰老过程中的基因表达总体相似,但也表现出一些差别,如TMs年龄相关的DEGs更多富集在减弱的代谢活动中,而CRs更显著地表现出免疫老化。
4.2 猕猴与人类在衰老过程中的差异
机体代谢水平和疾病易感性的改变是衰老的重要特征之一(Kenyon,2010;Ruby.,2018a),这些改变也反映在3个物种的血液转录组中。3种灵长类动物共同的38个下调DEGs富集到了B细胞受体信号通路(ko04662),该通路通过激活B细胞来启动下游的体液免疫。在标注了3个物种所有DEGs的B细胞受体信号通路富集图中,3个物种存在4个相同下调的环节(CD72、CD19、Igα、Igβ)。CD72和CD19是免疫球蛋白基因超家族的成员,Igα和Igβ与表面免疫球蛋白共同构成B细胞抗原受体复合物,CD72、CD19、Igα和Igβ都是B细胞抗原受体表达和功能所必需的分子。衰老对3个物种的共同影响体现在B细胞受体信号通路的起始节点,衰老下调起始物质的表达,从而影响B细胞免疫。另外与人类相比,CRs富集了更多的下调基因,TMs富集了更多的上调基因。由此推断年龄增长过程中,TMs整体的B细胞免疫可能比人类的更活跃,即拥有更强的体液免疫能力。在衰老过程中,CRs的B细胞数量显著下降,导致其衰弱的体液免疫(Didier.,2012)。推断在衰老过程中,体液免疫能力由强到弱依次是TMs、人类、CRs。这种能力的不同可能反映了3个物种免疫系统的差异。但这一推测还需要采用其他研究手段加以验证。
另一个相似的现象是蛋白质的翻译功能、核糖体的形成过程,这些过程的相关基因在这3种灵长类动物中都是随着年龄增长而下调。人类衰老过程中参与蛋白质翻译的基因显著下调(Jung.,2015)。蛋白质合成速率降低是衰老过程中的一个重要环节,会导致代谢衰减。但代谢衰减是衰老的原因,还是衰老过程产生的结果尚不明确(Anisimova.,2018)。
