基于多度格局模型的游泳动物群落结构研究-以闽南海域为例
2022-01-22蔡建堤刘勇徐春燕马超庄之栋
蔡建堤,刘勇,徐春燕,马超,庄之栋
( 1. 福建省水产研究所,福建 厦门 361013)
1 引言
游泳动物是海洋生态系统中重要的生态类群之一,具有较强的运动器官,在海水中能自由选择其行动路径,移动距离较长[1-2],其群落结构可用于评价海洋生态系统健康状况[3-4]。群落组成、分布、结构、稳定性、变化以及与不同环境因子相关关系是游泳动物群落结构研究的主要内容[5-12]。其中,物种个体数量是群落结构组成研究中最基础的内容之一,是群落结构的重要反映[10-12]。一个群落中每个种的多度(个体数量)从大到小排序,形成一个多度格局。多度格局反映了物种间多度关系,是群落结构的重要特点,对理解群落结构具有重要意义[13-14]。因此,多度格局引起了国内外学者的广泛关注。用数学方法结合生态学意义建立种-多度分布模型模拟多度格局的时空变化,是研究多度格局的重要手段[13-20]。目前,针对群落多度格局建立了许多适用于不同类型群落或集合的模型[13-20],并在很多方面开展了应用研究,如森林群落[21-28]、草地群落[29-31]、昆虫群落[32-34]等。海洋生态系统是多度格局研究的薄弱环节,应用多度模型研究海洋游泳动物群落多度格局,将弥补多度格局应用研究的不足,为揭示游泳动物群落结构的类型、性质和群落演替等提供帮助。
闽南海域是鱼类、虾类、蟹类和头足类等游泳动物产卵、索饵与越冬的重要场所[35]。自20世纪70年代以来,随着捕捞能力和海洋开发力度不断加大,闽南海域渔业资源的衰退程度不断加剧,许多重要经济种类资源及群体结构发生了明显变化,呈现小型化、低龄化和低值化的特点[36-39]。为进一步理解闽南海域游泳动物群落结构,本研究根据2019年秋季闽南海域渔业资源调查数据,应用多度格局模型分析了群落类型在组成上的特点及变化,探究了群落结构变化的生态学过程及影响因素,挖掘群落结构潜在的、有意义的特征,为从生态水平上合理管理渔业资源提供有价值的资料[40]。
2 材料与方法
2.1 材料
2019年9月,开展闽南海域渔业资源的专项调查,共设10个调查渔区(图1)。调查船为“闽东渔61792”底拖网船(渔船吨位为51 t,主机功率为183 kW,网口宽度为10.0 m,网口高度为5.5 m,囊网网目孔径为1.8 cm)。渔业资源调查按《海洋渔业资源调查规范》(SC/T 9403-2012)进行。环境因子(水深、温度、盐度、pH和溶解氧浓度)调查按《海洋监测规范》(GB 17378.1-2007)进行。本次调查取全部渔获物,带回福建省水产研究所生态实验室进行种类鉴定和生物学测定,对每个种类分别进行称重和计数。物种鉴定主要参考《福建鱼类志(上卷、下卷)》《中国动物志(头足纲)》《中国海洋动物图集(第六册)》《南海对虾类》等。
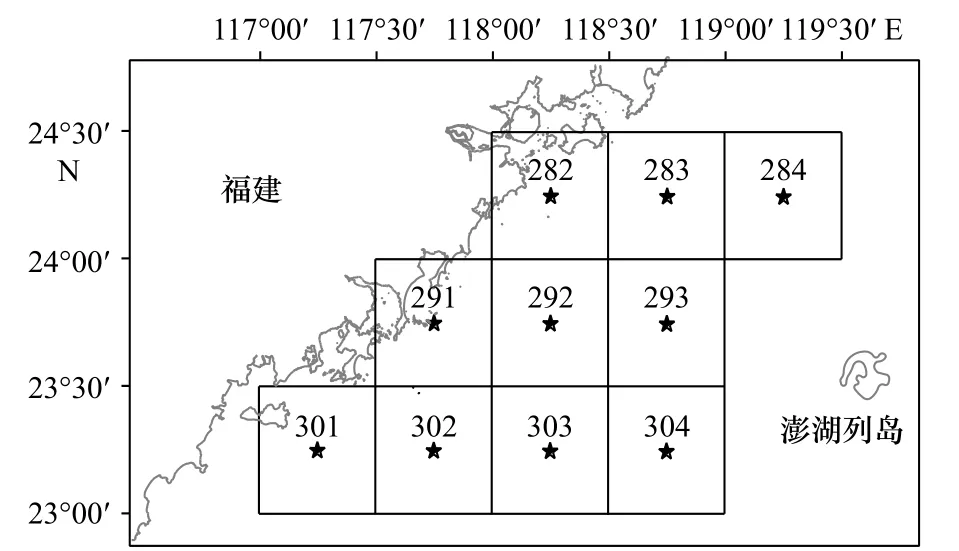
图1 调查站位Fig. 1 Survey stations
2.2 模型与优势度
多度格局模型可划分为生态位模型和统计类模型,其中生态位模型为多度格局的主要模型,它是具有明确生态意义的模型。多度分布模型达10多种[13-34],本实验选取了普遍认可和应用较广泛的5种生态位模型:生态位优先模型(Niche Preemption Model,NPM)、分割线段模型(Broken Stick Model, BSM)、重叠生态位模型(Overlapping Niche Model, ONM)、优势优先模型(Dominance Preemption Model, DPM)、随机分配模型(Random Assortment Model, RAM)[22-24,30]。
2.2.1 生态位优先模型
生态位优先模型要求优势种具有较强优势,认为最优势种不占绝对优势,其生态位小于总生态位的一半[3,16]。生态位优先模型假设一个群落中有S个种,有限资源为1,最优势种占有总资源的k份,剩下(1-k),第二优势种占有剩余资源的k份,即k(1-k),而剩余(1-k)2。依此类推,第(S-1)个种占有k(1-k)S-2,剩下的k(1-k)S-1为最后一个物种占用,这样就形成了一个从大到小的几何序列,则第i个物种的理论多度所对应的多度比例yi为
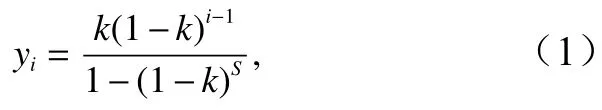
式中,S为物种数;i表示第i个物种;k为最优势种的实际多度比例。
2.2.2 分割线段模型
分割线段模型描述的是群落中缺乏相对重要性极强的物种,适合分类上比较接近的种形成的群落,个体数分配比较均匀。分割线段模型假定一个群落中的有限资源生态位(资源总量)是单位长度为1的一根棍,在其上随机设置S-1个点,把棍分成S个部分,每一部分长度表示一个种的多度,把S部分进行从大到小排序,即将物种从最普通到最罕见排列,则第i个物种的理论多度比例yi为

2.2.3 重叠生态位模型
重叠生态位模型允许种类在资源利用上重叠。假定群落资源总量视为一根棍,每个种的生态位等于棍上随机的两点间的距离,各个种是彼此独立的。重叠生态位模型不是把棍分配给各个种,而是每个种取其所需资源比例,这样群落的总生态位或资源总量不再是1,同一段棍可分成多个种占用的重叠生境,则第i个物种的理论多度所对应的多度比例yi为
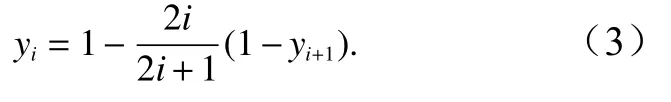
2.2.4 优势优先模型
优势优先模型显示,在群落中最优势种优先占用总生态位的一半以上,次优势种再占用剩余的一半以上,以此类推。此模型保证优势种优先利用资源,所有种在生态位占有上明显形成一个等级。Tokeshi[14]认为,此模型的k为0.75,第i个物种的理论多度所对应的多度比例yi为

2.2.5 随机分配模型
随机分配模型假定一个群落中各个种的多度之间没有联系,种间竞争不存在,多数情况下物种不能全部占用其生态位。此模型认为,物种对生态位是随机的选取,将S个物种占用生态位的大小从大到小排序,则第i个物种的理论多度所对应的多度比例yi为

2.2.6 优势度
McNaughton种类优势度指数[41]为
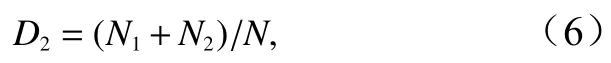
式中,D2为McNaughton 种类优势度指数;N为样品中的总个体数;N1、N2分别表示样品中居第一位、第二位种类的个体数。
2.3 数据处理
群落中游泳动物个体数为多度指标,游泳动物个体数与该群落总个体数的比值为多度比例,每个物种的多度指标从大到小排序,形成物种的多度排列。多度模型计算得到的所有物种的多度比例乘以所有物种的多度和,即可算出各种物种的理论多度。5个模型的生态学意义不同,其对同一群落的理论多度各不相同,将理论多度与多度排列进行线性回归分析,判定模型的理论多度分布与实际多度分布的拟合效果,结合模型的生态学意义,解释、分析游泳动物真实多度分布特征。数据整理采用Excel 2016软件,理论多度计算采用Matlab 2017b软件,数据分析、模型拟合、统计检验采用SPSS 19软件。
3 结果与分析
3.1 种类组成与数量分布
经鉴定,闽南海域游泳动物共有192种,隶属19目84科134属,其中鱼类132种,蟹类31种,虾类13种,头足类10种,口足类6种(附表A1)。

表A1 游泳动物名录Table A1 The list of nekton
10个渔区游泳动物以鱼类的种类数最多,其次为蟹类。鱼类、蟹类和头足类在10个渔区均有分布。284渔区、293渔区有3类游泳动物(鱼类、蟹类和头足类),类群数最少,种类数较少。鱼类种类数以292渔区、282渔区和304渔区较多,蟹类种类数以292渔区和282渔区较多,虾类种类数以282渔区最多,头足类种类数以302渔区较多,口足类以283渔区和292渔区较多,游泳动物种类数以292渔区最多,284渔区最少(图2)。
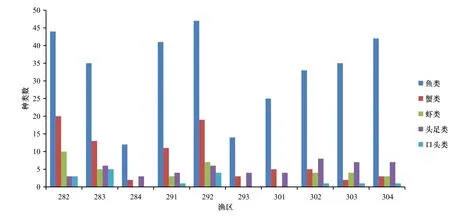
图2 不同渔区类群组成Fig. 2 Composition of groups in different fishing zones
闽南海域中南部近岸海域(291渔区、292渔区、293渔区、301渔区、302渔区)游泳动物个体数量较少,其尾数占闽南海域总尾数的6.00%、7.15%、5.69%、5.01%、6.26%。5种优势种的多度比例最小的渔区为292渔区,其值为36.28%,其余渔区多度比例均大于50%,其中284渔区、293渔区大于90%。5种优势种多度分布不均匀,284渔区第一优势种杜氏枪乌贼(Loligo duvaucelii)个体数占总个体数的91.05%,多度下降最快,优势种多度分布均匀性最差;293渔区第一优势种杜氏枪乌贼个体数占总个体数的69.20%,多度变化较大,优势种多度分布较不均匀,303渔区第一优势种绿布氏筋鱼(Bleekeria anguilliviridis)个体数占总个体数的46.50%,多度变化大,优势种多度分布不均匀;292渔区第一优势种细颌鳗(Oxyconger leptognathus)个体数占总个体数的10.41%,多度下降缓慢,优势种多度分布最均匀(表1)。
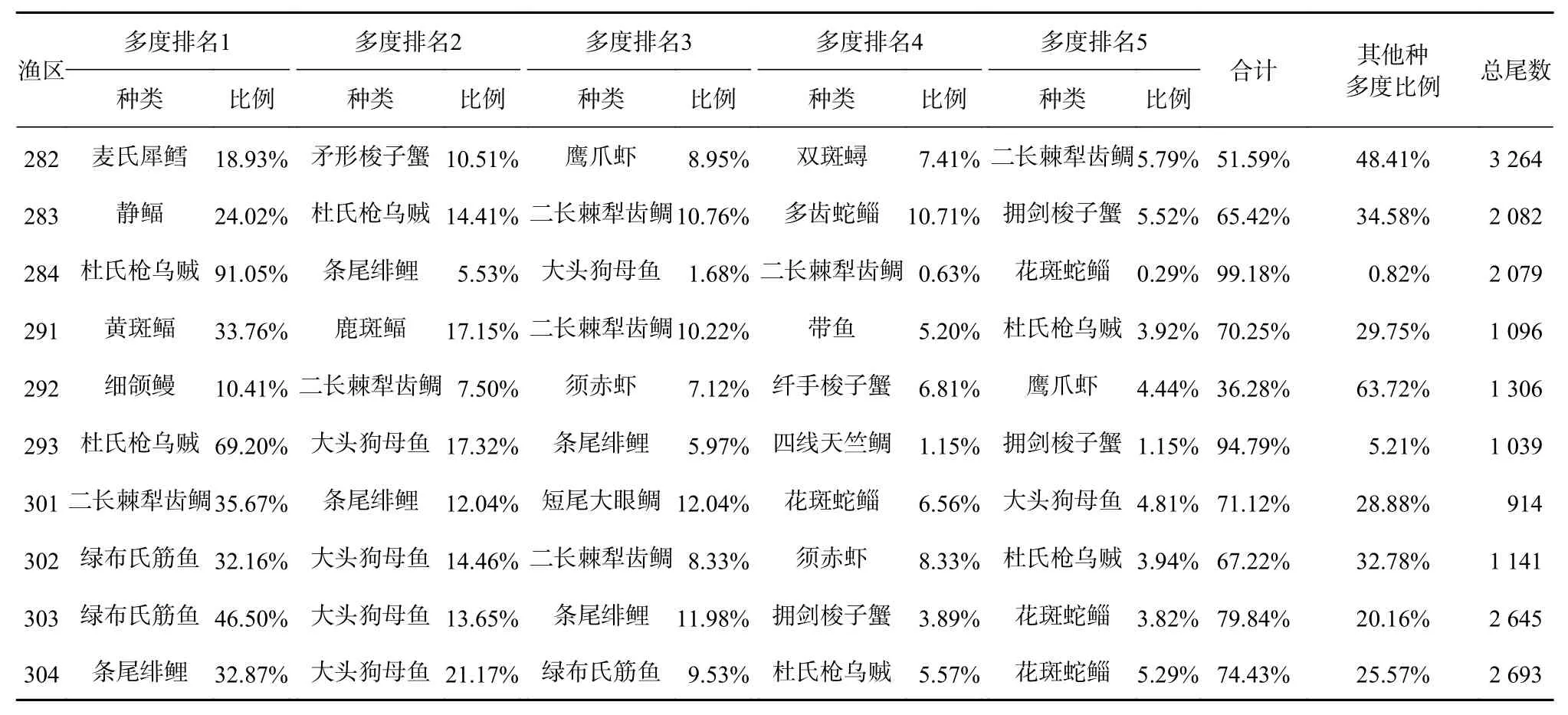
表1 不同渔区5种优势种的多度分布Table 1 Abundance distribution of five dominant species in different fishing zones
3.2 多度格局模型的拟合效果
闽南海域游泳动物群落的优势度指数为28.0%,NPM、BSM、ONM、DPM、RAM的理论多度与游泳动物真实多度的决定系数R2分别为0.953、0.516、0.286、0.618、0.797,且均小于0.001。NPM拟合游泳动物多度分布的效果最优,ONM的拟合效果不佳。同样,NPM对各渔区游泳动物多度分布格局效果较好,ONM的拟合效果较差(表2)。表明闽南海域游泳动物群落结构具有优势种占有较大优势和生态位之间不重叠的特点。不同多度格局模型拟合不同渔区群落多度分布的效果并不一致。DPM对284渔区和293渔区游泳动物多度分布的拟合效果理想,RAM拟合291渔区群落多度分布的效果较好,BSM拟合群落多度格局效果最好的渔区为292渔区,NPM对282渔区、283渔区群落多度分布的拟合效果优于其他多度格局模型(表2)。
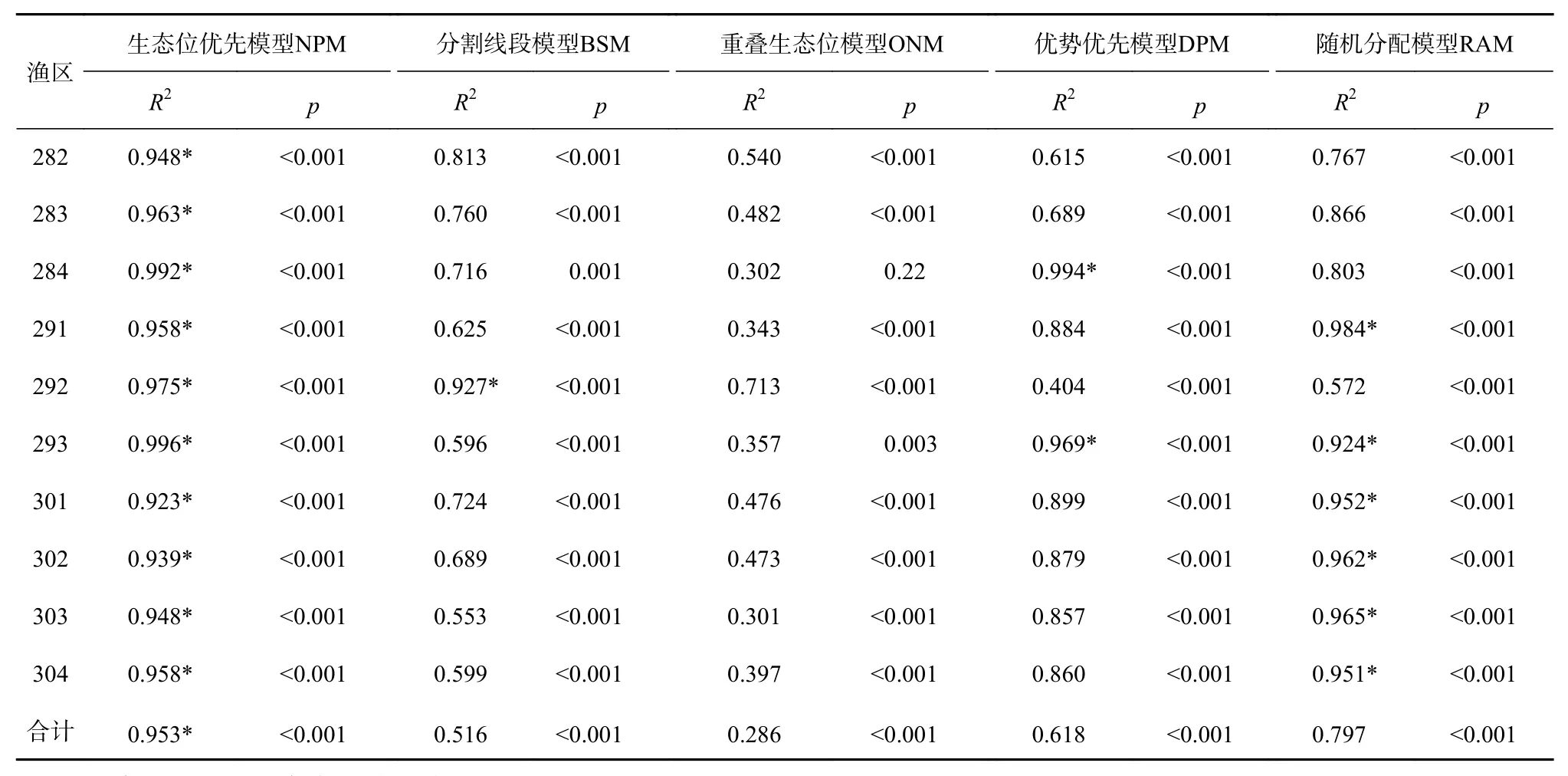
表2 5种模型对不同渔区游泳动物-多度数据拟合结果比较Table 2 Comparison of five models fitness to the species abundance data from nekton in different fishing zones
3.3 群落结构的空间变化
闽南海域的北部(282渔区、283渔区、284渔区)、中部(291渔区、292渔区、293渔区)、南部(301渔区、302渔区、303渔区、304渔区)的游泳动物真实多度与NPM的理论多度的决定系数R2分别为(0.948、0.963、0.992)、(0.958、0.975、0.996)、(0.923、0.939、0.948、0.958),p值均小于0.001(表2)。游泳动物真实多度与NPM的理论多度属于极显著相关。由外海到近岸的渔区,NPM的拟合效果渐差,优势种的地位逐渐降低。BSM对近岸渔区的游泳动物多度分布的拟合效果优于离岸渔区(表2),由外海到近岸的渔区,相对重要性极强的游泳动物逐渐缺失,个体数目分配越来越均匀,种类数逐渐增多。
水较深的渔区为284渔区、293渔区和303渔区,平均水深为52.1 m,游泳动物真实多度与DPM的理论多度决定系数R2分别为0.994、0.969、0.857,p值均小于0.001,种类数分别为17种、21种、50种,优势度指数分别为96.5%、86.5%、60.1%,而水较浅的渔区为282渔区、291渔区和301渔区,平均水深为28.2 m,游泳动物真实多度与DPM的理论多度决定系数R2分别为0.615、0.884、0.899,p值均小于0.001,种类数分别为80种、60种、35种,第一优势种个体数占总个体的比例分别为18.93%、33.76%、35.67%(表2,图3)。DPM在水深的渔区游泳动物多度分布拟合效果较好,沿纬度梯度从低纬度到高纬度,游泳动物多度分布的拟合效果渐好,优势种的优势地位逐渐升高,种类数变少,而DPM在水浅的渔区游泳动物多度分布的拟合效果较差,优势种的优势地位逐渐下降,种类数变多。
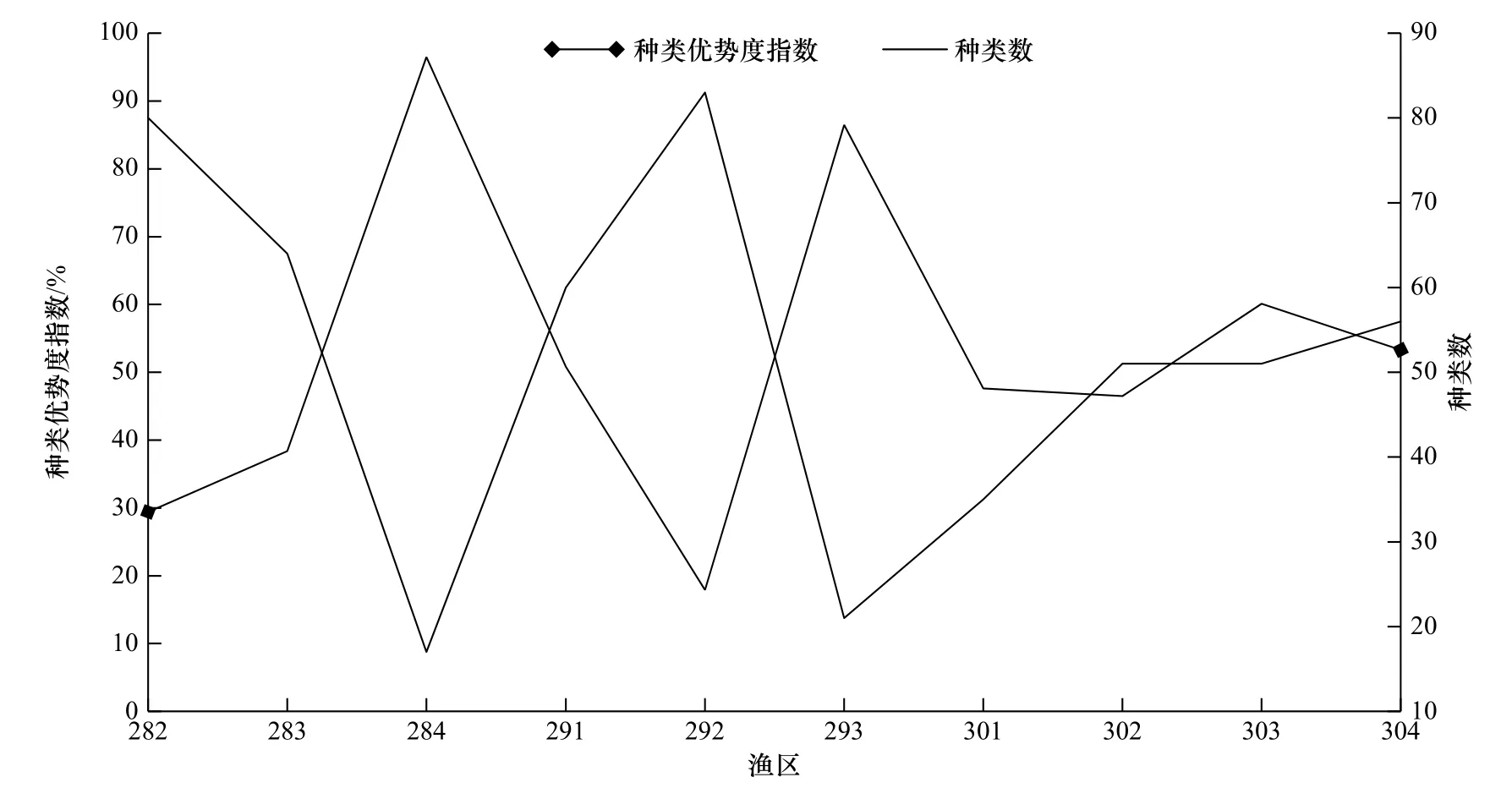
图3 不同渔区种类优势度指数和种类数Fig. 3 Species dominance index and number of species in different fishing zones
4 讨论
4.1 基于NPM、ONM的群落结构特征
在使用不同的生态学模型拟合物种多度分布后,能更好的反映出群落结构特征及其生态学意义。不同的多度格局模型均有各自的生态学过程含义,不同模型适合于不同情况。NPM要求优势种具有较大优势,仅适合于研究物种贫乏的环境或者群落演替的早期阶段[42-43],而ONM认为生态位之间有重叠,适用于资源丰富的群落类型[23]。NPM对闽南海域游泳动物多度分布的拟合效果均较好,而ONM的拟合效果并不理想。根据NPM和ONM的生态意义,要求群落结构需具备优势种有较大优势、物种数与个体数量贫乏、生态位之间不重叠和物种资源并不丰富的特征。过度捕捞导致闽南海域渔业资源衰退,种类数及群体数量严重下降[36-39],游泳动物资源贫乏,而部分游泳动物获得更多的空间和资源,种群数量增多,使其得以发展成为优势种,物种多度分布符合NPM的生态学意义。生态位重叠在海洋游泳动物群落中非常普遍[19],由于游泳动物资源并不丰富,物种有较大生存空间,种间的相互作用可能未达到生态位重叠的程度,表现为ONM的拟合效果不理想。
4.2 基于复合模型的群落结构特征
自然群落是复杂的,复合模型反映出,一个群落物种多度分布不只是遵循一种规律,生态现象及其内在机制不能依靠单个的生态模型来解释[14]。DPM要求优势种有绝对优势[22],可能仅适合于简单群落的研究[13]。DPM和NPM拟合284渔区群落游泳动物多度分布效果好,杜氏枪乌贼多度比例为91.1%,处于绝对优势地位,游泳动物种类数最少,为17种(图3),游泳动物种类最贫乏,类群数最少(仅3类)(图2),群落结构简单。RAM假设种的多度大小与生态位分配无关,种间竞争不存在,多数情况下物种不能全部占用其生态位,这样的群落体系的一个最大的特点是没有足够的时间提供给基于资源竞争的内部关系去发展进化[30]。RAM拟合种群数量较少的渔区(291渔区、292渔区、301渔区、302渔区)物种多度分布拟合效果较好。形成此现象的原因可能是这些渔区种群数量少,导致多数物种不能全部占用其生态位,而不是基于资源竞争。RAM、NPM和DPM模拟293渔区群落游泳动物多度效果好,杜氏枪乌贼的多度比例为69.2%,占有绝对优势,物种数较少,为21种,总个体数较少,为1 309尾,游泳动物资源处于贫乏状态,游泳动物有较大的生存空间,种间关联小,类群数最少(仅3类)(图2),群落结构相对简单。RAM、NPM模拟291渔区、301渔区、302渔区、303渔区和304渔区群落游泳动物多度效果好。闽南渔场8月16日伏季休渔结束后,大量的捕捞活动对这些渔区个体较大的经济鱼种的损害高于个体小的非经济鱼种,会有一些个体大的经济种类,如带鱼(Trichiurus lepturus)、蓝圆鲹(Decapterus maruadsi)等种群数量大幅减少,在持续高强度捕捞压力下,没有足够的时间去发展壮大,无法占用全部生态位,表现出物种多度大小与生态位分配无关,由于种间竞争弱,个体小的非经济鱼种,如黄斑鲾(Leiognathus bindus)种群数量增多,进而发展成优势种,其数量占全部个体的大多数,符合NPM的生态学意义。分割线段模型是描述群落中缺乏相对重要性极强的物种,在生态位上的分布相对均匀,此分布符合随机生态位假说[44-45]。NPM和BSM适合拟合292渔区群落游泳动物多度分布,说明群落结构中个体数量上的优势没有得到足够的发展,不同种类的数量虽然有差别[44],但是不显著,该群落细颌鳗多度比例最大,为10.41%,其次为二长棘犁齿鲷(Evynnis cardinali)、须赤虾(Metapenaeopsis barbata)和纤手梭子蟹(Portunus gracilimanus),多度比例分别为7.50%、7.12%、6.81%。NPM拟合效果要稍好于BSM,可能是群落处于早期阶段而导致群落的物种多度近于相等[23,45]。
4.3 群落结构变动的响应机制
游泳动物群落结构的影响因素复杂多样[46],如种类、个体发育阶段、生理性状况、各种生活机能、栖息的环境条件等。鱼类群聚分布与水深有关[47]。在同一纬度,随着水深的增加,群落多度分布格局NPM的拟合效果逐渐增强,BSM的决定系数R2逐渐变小,DPM对水深的海域的拟合效果最好,说明随着水深的增加,优势种的地位从非绝对优势到绝对优势、生态位分配从不合理到比较合理、群落结构从复杂到简单。捕捞对游泳动物群落结构有重要的影响[48],近岸渔区捕捞压力较大,在捕捞干扰下,近岸海域大个体生物群体的优势种大量损失,优势种优势地位下降,群落缺乏相对重要性极强的游泳动物,小个体生物群体得以发展,个体数目分配均匀度逐渐增大,物种数量逐渐增大,DPM拟合物种多度分布效果较差,而外海渔区受人类干扰相对较少,优势种有较大生存空间,优势地位进一步升高,DPM拟合物种多度分布效果较好。温度也是影响物种多度分布的重要因素[47]。随着水温的升高,水深的渔区(303渔区、293渔区和284渔区)优势种的优势地位逐渐升高,DPM的拟合效果越好。优势种在数量上的优势程度对游泳动物群落结构产生影响,决定物种多度分布格局。在生态学上,优势种对整个群落具有控制性影响[31],优势种所占资源比例的大小势必影响其他物种的多度分布,改变物种多度分布格局。从数学理论上来说,生态位优先模型的k小于0.5,RAM等同于k等于0.5的生态位优先模型,而DPM等同于k等于0.75的生态位优先模型。在实践上,RAM拟合优势度指数50%左右的群落多度分布拟合效果较好,NPM对优势度指数明显小于50%的群落多度分布拟合效果优于其他多度格局模型,DPM对优势度指数大于75%的群落多度分布拟合效果优于其他多度格局模型,BSM对优势度指数小于20%物种多度分布有较好的拟合效果(表2)。
4.4 群落结构维持与演替机制
群落结构维持机制一直是群落生态学研究的核心问题,其中的关键是物种的共存机制[49-50]。生态位理论认为,群落内的不同物种如果要共存则必须有生态位的分化,且生态位分化等确定性因素占主导地位,而群落中性理论认为,在满足个体水平的生态等价性和群落饱和性两个基本假设下物种可以共存,扩散和随机作用是主要决定因子[51-54]。从本文的研究结果来看,基于生态位的多度格局模型研究闽南海域游泳动物多度分布是成功的,生态位理论可以解释物种共存的现象[55]。闽南海域游泳动物优势种聚集和种间分离是群落主要空间构型,物种间的空间分离减弱种间竞争,阻止或减缓竞争优势物种对竞争劣势物种的竞争排除作用,进而维持物种共存,有利于群落结构的维持。闽南海域渔业资源衰退,游泳动物种类数及群体数量严重下降[37-40],因而生态位在中性理论中没有充分发挥作用。如果渔业资源恢复到群落处于饱和的状态,中性理论也许在解释闽南海域的群落结构特征方面存在潜力。如今许多生态学家认为,生态位理论和中性理论应是相容互补的而不是矛盾的[54-57]。Tilman[58]认为,中性作用存在于生态位群落构建中,提出了随机生态位理论。NPM拟合292渔区物种多度分布的拟合效果好,生态位理论可以解释物种共存现象[55],而BSM拟合292渔区物种多度分布拟合效果,揭示了随机过程对群落结构的影响[34],随机生态位理论能够很好地解释292渔区群落结构维持、物种多度分布和物种共存等现象,说明经典生态位理论和中性理论并非绝对对立。
群落演替也是游泳动物群落结构研究的重点和难点。生态位理论认为,群落构建是确定性过程,群落之间有着可分辨的边界,群落在受干扰后能够逐渐演变到原来的状态,在群落物种的相互作用下,演替是从一个方向有规律地向另一个方向变化,演替的最后阶段是稳定的单元顶极或多元顶极[52]。当生态学过程发生变化时,物种多度格局会从一种模型转变为另一种模型,所以多度格局可以指示群落演替的生态过程变化[17,19,22]。从本文的研究结果来看,近岸海域受人类干扰活动程度大于外海海域,近岸海域物种多度拟合效果较好的模型主要为NPM,而外海海域主要为DPM,如果减少人类活动的干扰,近岸海域游泳动物多度分布格局由DPM向NPM转变,群落结构将可能从复杂到简单、优势种将向绝对优势的方向演替。可见,运用多度格局模型进行多度格局拟合是深入认识海洋游泳动物群落演替生态过程的有效手段。然而,由于海洋生态系统本身极度的复杂性,生态位模型还不能够完全真实地反映海洋游泳动物群落演替的全部过程,相关群落演替研究的理论、方法和技术手段都远未成熟,但模型对于海洋游泳动物群落演替部分过程的真实反映依然能够为渔业管理提供依据。
附录

续表 A1

续表A1

续表 A1
续表 A1
致谢:感谢叶孙忠、谢少卿为本文提供数据,谨致谢忱!
