亚胺培南作用下的大肠埃希菌膜渗透性改变的分子机制研究*
2022-01-14何建春赵峻英裴昌贞
何建春,杨 雷,赵峻英,董 剑,王 丹,裴昌贞
(重庆市大足区人民医院:1.检验科;2.睡眠心身中心 402360)
大肠埃希菌是临床最常见的条件致病菌,根据中国CHINET数据显示,其分离率一直居所有监测菌的首位。随着抗生素的广泛使用,产超广谱β-内酰胺酶(extended-spectrum β-lactamases,ESBLs)大肠埃希菌的分离率也越来越高[1-2]。碳青霉烯类抗生素一直被认为是治疗该类菌株引发感染最有效的抗生素。相关文献报道亚胺培南(imipenem,IPM)是碳青霉烯类抗生素中使用频率最高且效果最好的抗生素之一[3-5]。而在临床治疗过程中,很多时候由于感染灶不明确、患者个体药物吸收差异等原因使得IPM到达感染灶的剂量不足,导致临床治疗失败。在IPM作用于大肠埃希菌时,大肠埃希菌为了生存,主要通过什么途径来防御IPM的杀灭作用,膜孔蛋白和外排泵各自是否产生作用,到目前为止还不清楚,值得深入研究。
1 资料与方法
1.1 一般资料
回顾性收集大足区人民医院2019年1—6月痰液、血液、尿液等标本中首次分离的大肠埃希菌共16株,先分别选择2株非产β-内酰胺酶大肠埃希菌、2株产ESBLs酶大肠埃希菌、2株产AmpC酶大肠埃希菌、2株产ESBLs酶+AmpC酶的大肠埃希菌共8株,进行膜孔蛋白和外排泵差异基因的筛选。再按照上述选菌方式再选择8株大肠埃希菌进行差异基因的验证。
1.2 方法
1.2.1菌株鉴定及最低抑菌浓度(MIC)值
根据第4版《全国临床检验操作规程》进行微生物样本的培养和纯化,然后通过Vitek 2 Compact系统进行菌株鉴定及药敏测定,并依据CLSI M100-S29进行结果判读。
1.2.2微量肉汤稀释法
依据CLSI推荐的方法进行MIC值测定,配制好相应浓度的抗生素溶液和菌悬液,分别加入96孔板中,并设定阳性和阴性对照,孵育18~24 h后进行结果判读。
1.2.3ESBLs确证实验和ampC酶筛选实验
ESBLs确证实验[6]:选取Vitek 2 Compact初筛为产ESBLs的大肠埃希菌,然后选择药敏纸片头孢他啶(30 μg)、头孢他啶/克拉维酸(30/10 μg)、头孢噻肟(30 μg)和头孢噻肟/克拉维酸(30/10 μg)完成纸片扩散法实验,若头孢他啶/头孢噻肟加克拉维酸与不加克拉维酸的抑菌圈直径之差大于或等于5 mm,则为产ESBLs。
ampC酶筛选实验[7]:运用K-B纸片扩散法,贴上FOX药敏纸片,若FOX抑菌圈直径小于或等于18 mm则初步判定为产ampC酶。大肠埃希菌标准菌株ATCC25922作为阴性对照。
1.2.4ESBLs基因和ampC基因的PCR扩增
利用PCR法检测ESBLs基因(blaCTX-M、blaTEM和blaSHV)和ampC基因(blaDHA、blaEBC、blaCIT、blaACC、blaMOX 和blaFOX)。反应中所需引物序列和反应条件参考文献[7]。
1.2.5IPM进行菌株诱导
菌株诱导参考孙坤玲[8]的研究:按照说明书配制新鲜无菌LB肉汤,并配置相应浓度的IPM抗生素溶液加入上述LB肉汤中,得到含IPM的LB肉汤。再将本实验的8株新鲜菌分别用无菌LB肉汤进行比浊,加入上述含有抗生素的LB肉汤中,最终使实验菌在3/4 MIC值(亚抑菌浓度)的含IPM的LB培养基中生长,37 ℃摇床中培养8 h。
1.2.6RT-qPCR方法
将实验的8株原始菌和诱导8 h后的菌分别进行RT-qPCR检测基因表达量,检测的基因包括膜孔蛋白基因(ompF和ompC)、外排泵基因(acrA、acrB、tolC、macB和emrA)和外排泵调控基因(marA)。最后将上述差异基因在另外8株菌中进行统计学验证,验证差异基因是否在所有菌中具有统计学差异。其反应所需引物序列和反应条件参考文献[9-10]。
1.3 统计学处理

2 结 果
2.1 菌株的基本特征
8株大肠埃希菌从标本来源看:2株来源于痰标本,2株来源于血液标本,3株来源于尿液标本,1株来源于脓液标本。从产β-内酰胺酶来看:2株不产β-内酰胺酶,2株产ESBLs 菌株分别携带blaTEM和blaCTX-M,2株产ampC酶,2株产ESBLs酶+ampC酶。从MIC值来看:其对IPM的MIC值为0.125~1.0 μg/mL,对美罗培南(MEM)的MIC值为0.125~0.5 μg/mL和对厄他培南(ETP)的MIC值为0.125~0.5 μg/mL均为敏感,见表1 。
2.2 治疗后菌株的特征
对比分析8株菌在IPM治疗前后MIC值变化情况,结果发现:对IPM的MIC值无变化的有1株菌,增加2倍的有4株菌,增加4倍的有3株菌。而对MEM的MIC值无变化的有3株菌,增加2倍的有4株菌,增加4倍的有1株菌。而对ETP的MIC值增加2倍的有3株菌,增加4倍的有1株菌,增加8倍的有4株菌。诱导后引起IPM、MEM和ETP的MIC值发生改变的百分比达到83.3%(20/24)。而从4种类型β-内酰胺酶菌株诱导前后MIC变化情况来看,不产β-内酰胺酶菌株基本保持不变,而只产ESBLs酶/ampC酶菌株有一定变化,变化最大的为产ESBLs酶+ampC酶菌株,见表1。
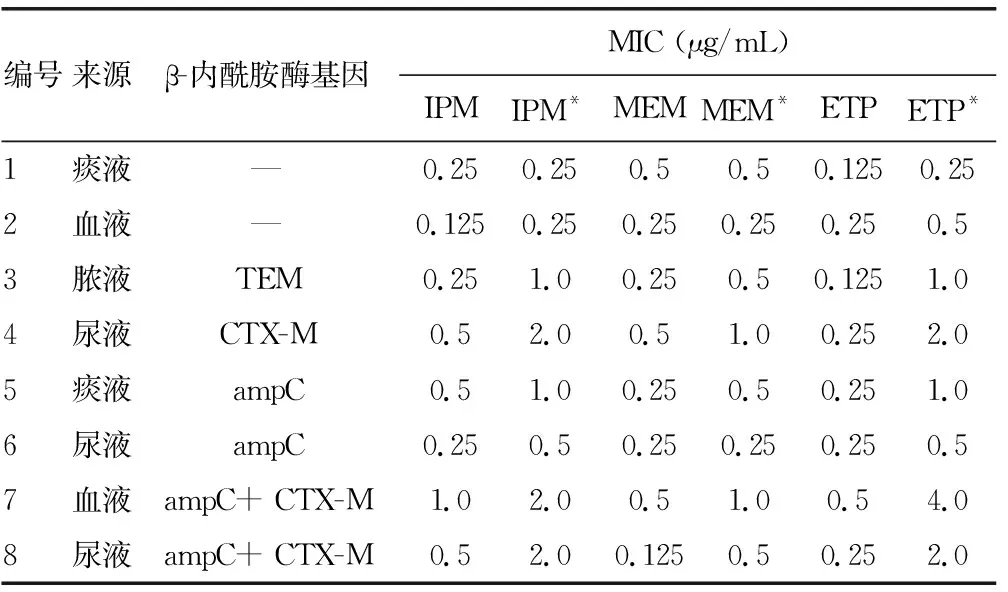
表1 实验菌株的基本特征
2.3 治疗前后菌株膜孔蛋白表达情况
对比分析8株菌在IPM治疗前后膜孔蛋白ompF和ompC表达量的变化情况,结果发现:与8株原始菌相比,诱导后菌株的ompC基因表达虽然均有不同程度的下调,但差异均无统计学意义;而对ompF基因来说,诱导后菌株的ompF基因表达也均有不同程度的下调,且有5株菌表达差异有统计学意义(P<0.05),见图1。
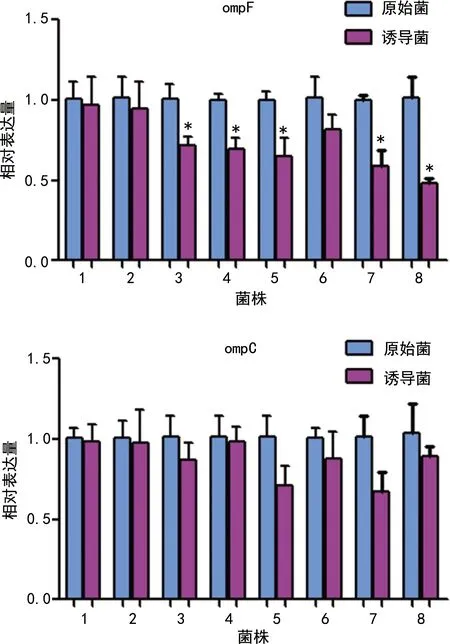
图1 IPM诱导对大肠埃希菌膜孔蛋白基因表达的影响
2.4 治疗前后菌株外排泵基因表达情况
对比分析8株菌在IPM治疗前后外排泵表达量变化情况,结果发现:与8株原始菌相比,绝大部分诱导菌外排泵基因表达有不同程度的上调,对tolC和acrB基因来说,除第1株不产β-内酰胺酶的菌差异无统计学意义外,其余7株菌差异均有统计学差异(P<0.05);对acrA基因来说,所有菌株的表达量差异均有统计学意义(P<0.05);对macB基因来说,所有菌株表达量差异均无统计学意义;对emrA基因来说,仅有2株菌表达差异有统计学意义(P<0.05),其余菌表达量差异均无统计学意义,见图2。
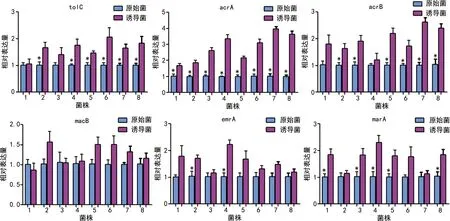
图2 IPM诱导对大肠埃希菌外排泵及调控基因表达的影响
2.5 菌株治疗前后外排泵调控因子表达情况
对比分析8株菌在IPM治疗前后外排泵调控因子表达量的变化情况,结果发现:与8株原始菌相比,诱导菌的marA基因表达虽然均有不同程度的上调,但第2、6和7共3株菌表达差异无统计学意义,其余株菌差异均有统计学差异(P<0.05),见图2。
2.6 acrA和ompF基因表达在更多临床菌株中的验证
进一步分析acrA和ompF基因在16株临床菌经过IPM治疗前后的表达差异。结果发现:原始菌组与诱导菌组acrA基因的表达分别为1.00±0.36、2.36 ±0.58,差异有统计学意义(P<0.01);ompF基因的表达为1.00±0.09、0.72±0.11,差异有统计学意义(P<0.01),见图3。
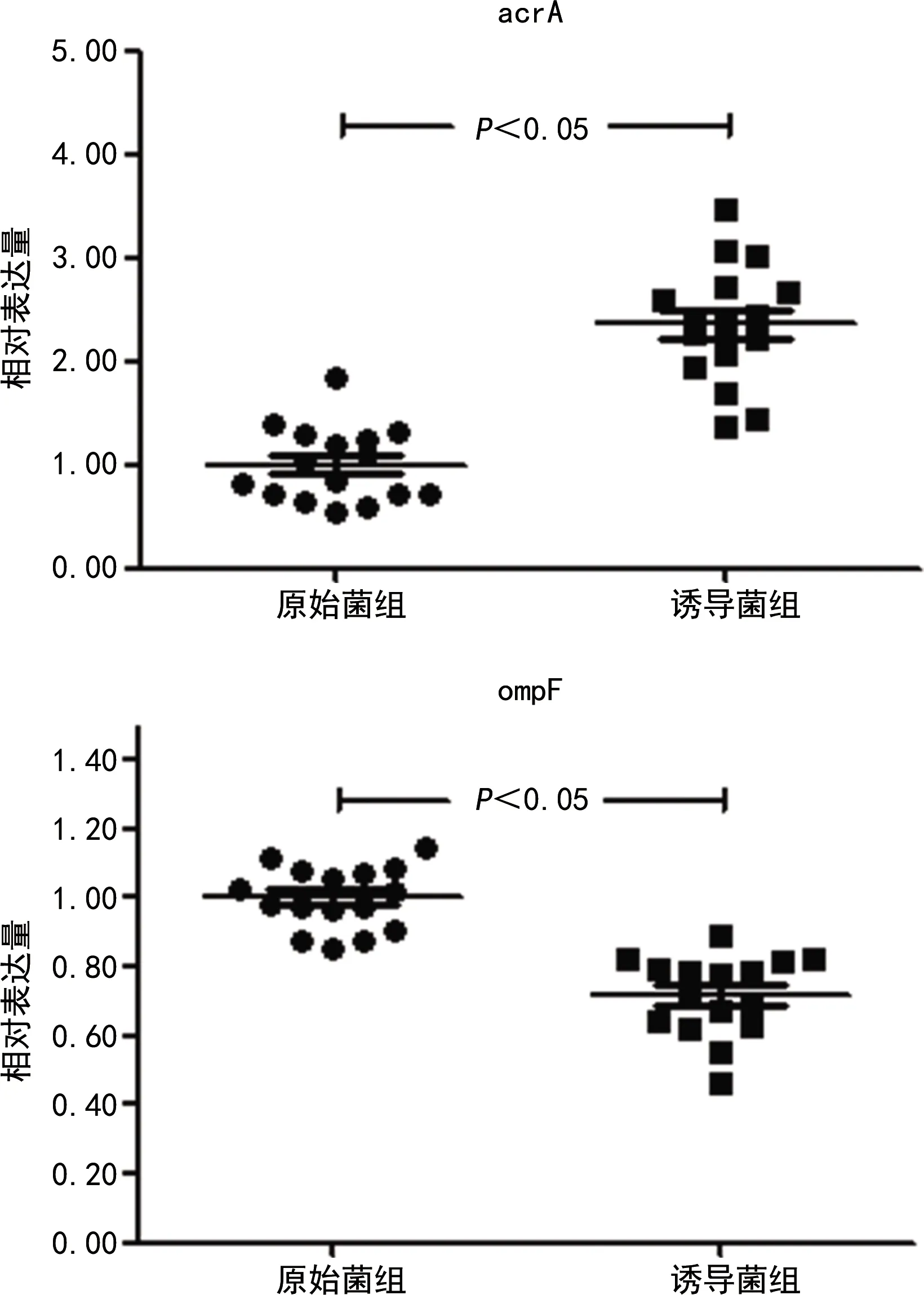
图3 IPM诱导对大肠埃希菌acrA和ompF基因表达的影响
3 讨 论
大肠埃希菌是临床所有分离菌株中检出率最高的细菌[2],由于产ESBLs酶,甚至产ampC酶、乙酰转移酶和/或氨基糖苷类钝化酶等产酶大肠埃希菌的广泛出现,使得临床使用大部分常见抗生素进行抗感染治疗均达不到理想效果[11]。此时,碳青霉烯类抗生素常常被临床使用,虽然耐碳青霉烯类大肠埃希菌已经出现,甚至达到了2%的检出率[2],但仍然是治疗产 ESBLs等酶大肠埃希菌最有效药物。其中亚胺培南是碳青霉烯类抗生素中临床使用率最高一种抗生素,也是第一个批准上市的碳青霉烯类药物[3,5]。因此,本研究首次实验并分析了在IPM作用下,大肠埃希菌膜孔蛋白和外排泵所产生的作用,从而为细菌和抗生素的相互作用机制提供依据,甚至新药的研发提供帮助,并进行如下讨论分析。
从所选菌株是否产β-内酰胺酶来看,实验菌中分别为2株不产β-内酰胺酶、2株只产ESBLs酶 、 2株只产AmpC酶和2株共产ESBLs酶+AmpC酶菌,使得实验能够多角度观察分析大肠埃希菌在IPM治疗下的膜孔蛋白和外排泵的变化。从IPM治疗前后菌株的MIC值变化来看,不产β-内酰胺酶菌基本保持不变,产生这种现象的可能原因是仅靠膜孔蛋白基因下调或者外排泵基因上调可能不足以引起碳青霉烯类抗生素有明显变化。而只产ESBLs酶/AmpC酶菌有一定变化,变化最大的为产ESBLs酶+AmpC酶菌;其中又以ETP的MIC值变化最大,甚至有些治疗后的菌达到了耐药水平。产生这种现象的原因可能是DELPHINE等[12]研究发现的CTX-M型基因的高表达合并膜孔蛋白下调能够引起ETP敏感性下降相关,也可能是部分研究者[13-14]发现的产ESBLs酶和/或AmpC酶合并膜孔蛋白缺失或外排泵的高表达引起碳青霉烯耐药相关。由于以上原因,相关研究[15]还建议在临床治疗产ESBLs酶和/或AmpC酶大肠埃希菌时,尽可能少用ETP治疗。本研究还发现,在产ESBLs酶和/或AmpC酶大肠埃希菌中,IPM和MEM的MIC值也发生了变化,说明在临床使用IPM治疗的过程中,对于产ESBLs酶和/或AmpC酶大肠埃希菌,也需要合理用药,否则很容易导致临床抗感染治疗失败,甚至导致耐药菌的出现。
从IPM治疗前后菌的膜孔蛋白表达结果来看,ompC基因表达差异无统计学意义,而ompF基因表达明显下调,且大部分差异有统计学意义。研究[16-17]表明,ompF是膜孔蛋白中最主要最重要的孔蛋白,大部分抗生素包括IPM进入细菌需通过ompF,且膜孔蛋白ompF的缺失能引起抗生素MIC值增高。而本研究第一次探讨了在IPM诱导下,大肠埃希菌最开始为了适应和生存在IPM环境中,细菌作出的其中一个重要的防御反应为膜孔蛋白ompF表达下调。从外排泵基因表达结果来看,本研究也第一次探究分析发现在IPM治疗下,macB和emrA基因表达差异无统计学意义,而外排泵tolC、acrA和acrB基因表达明显增加,且差异有统计学意义,说明IPM也能够诱导外排泵基因表达的改变,是大肠埃希菌作出的另一个重要防御反应。相关研究[10,18]表明外排泵也是控制抗生素进入细菌的重要因素,其中acrAB-tolC系统是目前为止最主要的与抗生素耐药相关的外排泵系统。
总之,在IPM治疗下,产ESBLs酶和/或ampC酶的大肠埃希菌更能引起对碳青霉烯类抗生素MIC值的增高,主要是通过膜孔蛋白ompF基因表达下调和外排泵acrAB-tolC基因表达上调引起的。进而能够适应碳青霉烯类抗生素,导致临床抗感染治疗失败。因此,临床应当合理使用IPM,以减少耐碳青霉烯类大肠埃希菌的流行。这也有助于理解大肠埃希菌最开始为了适应和生存在IPM环境中,细菌作出的重要防御反应。
