不同煤阶煤制生物甲烷的代谢功能差异性研究
2022-01-13王子升周艺璇杨伟强王银伟
侯 彪,王子升,周艺璇,杨伟强,王银伟
(1.河南理工大学 能源科学与工程学院,河南 焦作 454000;2.国家能源投资集团 神东技术研究院,陕西 神木 719315)
0 引 言
厌氧消化最早是在生物有机体的自然生理过程中引入的,现已被战略性地应用于煤层生物气资源的开采与利用。研究表明,煤的生物利用度可能会限制生物气生成的速率和产量[1-2]。因此,目前大部分研究主要集中在如何增加煤的溶解性以及解聚性。然而,煤层生物气生成的主要条件不仅包括底物煤,还存在较复杂的生物化学反应,即水解、酸化、产酸和产甲烷等决定性步骤[3]。根据条件的不同,每个步骤都有可能成为限制生物气生成速率和产量的关键[4]。例如,产酸细菌与水解部分有关,可作用于聚合物(主要是多糖和蛋白质)产生短链脂肪酸(如乙酸、丁酸),使这些小分子酸类物质用于由产甲烷古菌驱动下的进一步甲烷的生成[5]。值得注意的是,任何上述成分或者含量发生变化都会导致厌氧消化过程出现差异。
据统计,大多数煤制生物甲烷的研究主要集中于煤的固体结构及液相产物变化对最终甲烷生成的影响[6-9]。然而,厌氧微生物中的功能基因和一些相关酶的表达对底物(即碳水化合物、蛋白质等)代谢以及最终生物甲烷的生成至关重要[10-11]。而现阶段关于这一主题的研究内容很少。尽管对于煤阶与微生物群落结构和分布特征之间的相关性已得到初步证实[12]。但转化过程中功能性酶的基因表达和甲烷产量之间的联系仍然不清楚。
鉴于基因组学技术的快速发展,目前可以实现在复杂条件下对参与生物甲烷生成的关键基因和功能酶的鉴定[13]。因此,基于煤的生物产气潜力与关键基因和功能表达的耦合关系,分析了不同煤阶煤制生物甲烷过程中的产气特征,同时评估了以不同煤阶煤作为厌氧发酵底物下的甲烷生成潜力。之后通过16S rRNA基因功能预测技术分析了不同煤阶煤制生物甲烷过程中参与底物代谢的相关基因功能表达水平。研究将煤的生物产气特征与微生物基因功能表达进行了有机结合,为生物成因煤层气的潜在生成机制提供新的见解。
1 试 验
1.1 试验材料
试验煤样选择内蒙古胜利煤田、河南义马千秋煤矿、山西大同泉岭煤矿和河南平顶山煤矿新鲜的褐煤、长焰煤、气煤和焦煤,煤样工业分析和元素分析分别按照国家标准GB/T 30732—2014和GB/T 31391—2015进行,测试结果见表1。试验前将煤样粉碎至80~100目(0.15~0.18 mm),并在60 ℃下干燥24 h,密封保存。
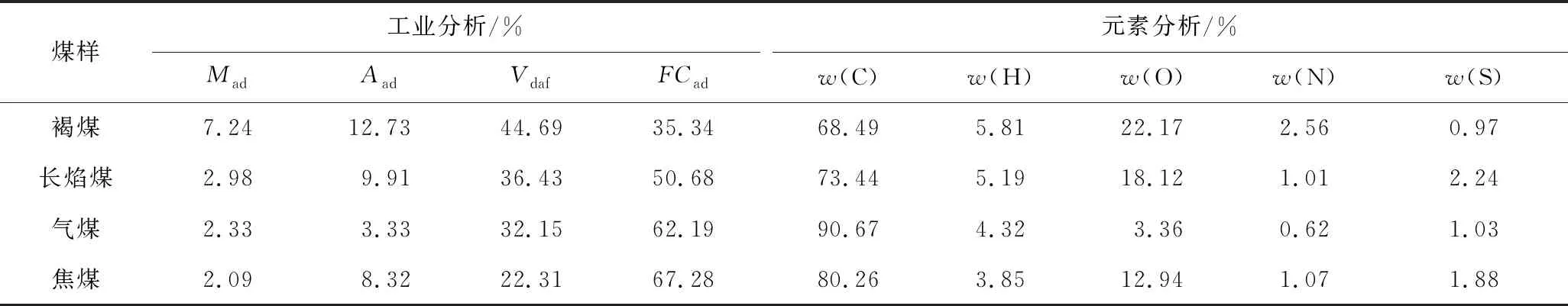
表1 煤样的工业分析与元素分析
1.2 生物产气试验
培养基配制方案按照文献[14]进行准备。将配制好的培养基放入高温灭菌锅内于121 ℃灭菌3 h,灭菌结束后按照3∶1的比例倒入驯化的菌液(来源为矿井水),利用N2驱替瓶内空气3 min,用封口膜密封,放置在35 ℃恒温培养箱中富集4 d。将20 g粉碎后的煤样加入250 mL锥形瓶内,同时加入富集4 d的菌液200 mL,利用N2驱替瓶内空气3 min,搅拌均匀后放入35 ℃恒温培养箱进行为期32 d的产气实验,每组实验设置3个平行样。用集气袋收集气体,甲烷定量分析使用气相色谱仪(Agilent 7890GC)。分别用HM、CYM、QM和JM表示以褐煤、长焰煤、气煤与焦煤为底物的生物产气试验。
1.3 甲烷生成动力学
修正的Gompertz模型通常用于模拟厌氧发酵过程中CH4的累积生成特征,使用该模型(式(1))评估和对比不同煤阶煤制生物甲烷过程中CH4生成动力学,该等式描述如下:
(1)
其中:y为累积甲烷产量,mL/g;A为累积甲烷产量潜力,mL/g;μm为最大比甲烷产量,mL/(g·d);λ为滞后期,t,为CH4首次产生所用的时间,也表示微生物适应环境所用的时间。
1.4 微生物代谢功能预测
4个试验组的发酵液的全基因组DNA是用E.Z.N.ATMMag-Bind Soil DNA Kit试剂盒提取的。使用Qubit 2.0评估DNA浓度,以确保达到测序要求。在测序之前,每个发酵液中的聚合酶链反应产物以等摩尔量标准化,用于构建聚合酶链反应扩增子文库。之后所有样本被送到上海生工进行测试,根据说明使用Illumina MiSeq(2×300 bp)系统进行文库构建,最终利用PICRUSt软件分析不同发酵液的代谢功能与功能基因差异。
2 结果与讨论
2.1 煤制生物气产气特征分析
对不同煤阶煤厌氧发酵过程中的累积产气量与甲烷产量进行了分析,结果如图1所示。
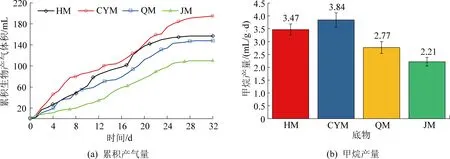
图1 累积产气量与甲烷产量生成特征
从图1可以看出,CYM的累积产气量明显高于其他试验组(图1a),且生物甲烷产量较HM、QM和JM分别增加了10.82%、38.88%和73.49%(图1b)。前人的研究证实,煤的生物产气量会随着煤阶增加而逐渐降低。这是因为与低阶煤相比,高阶煤的杂原子含量较低,芳香结构缩合度较高[15],导致其生物利用度降低。然而,FALLGREN等[16]的研究表明高阶煤可能比低阶煤具有更高的甲烷生成潜力。同时,有学者证实沥青比干酪根更具微生物转化甲烷潜力[17]。这表明煤阶高低不是生物甲烷产量潜力的有利指标。煤中的金属元素含量、黏土矿物以及导电性对发酵环境也有重要的影响。产生的影响如下:
1)金属元素。①释放金属离子,是厌氧发酵中厌氧微生物的必要营养来源[18];②有助于DNA复制、关键酶的形成[19];③降低了发酵体系中H2S的含量[20]。
2)黏土矿物。具有较高的比表面积和规则的孔道结构,吸水性和膨胀性较好。黏土吸水使其层间距增大,可提高微生物与煤的有效接触,且黏土中的无机矿物对微生物产甲烷有促进作用[21]。同时,起到pH缓冲作用,从而稳定厌氧发酵系统。
3)导电性。激活细胞间的信号传递,可在细菌和氢营养型产甲烷菌之间形成间直接电子转移的通道。SU等[22]研究证实在内蒙古区块采集的褐煤中高的金属元素含量(是长焰煤的9倍)会对微生物生长有毒害作用,同时也会抑制酶的活性甚至失活,这是导致褐煤的煤制生物甲烷量下降的原因之一。同时,煤中黏土矿物的含量会随着煤阶的增加而增多[23],长焰煤中黏土矿物含量较褐煤有所增加,有利于生物甲烷的生成。先前的研究表明,羧基在低阶煤中含量丰富,并且随着煤阶的升高而缓慢下降[24],羧基是降解的优先目标,与芳香族和脂类C—C键相比,其键离解能相对较低,可被微生物优先利用,同时高阶煤顽固的芳香结构不利于煤的生物降解[25]。综上所述,长焰煤可作为煤制生物甲烷的优势底物。
另外,前人的研究已经证实,煤的生物产气可分3个重要阶段,即①缓慢增长→快速增长阶段;②快速增长→缓慢增长阶段;③缓慢增长→产气停止阶段。从图1a中可以发现,产气量越大的试验组3个阶段越明显,且HM和CYM在16~24 d产气量增幅较大,而QM与JM的累积产气量没有较明显上升期。
2.2 甲烷的动力学模型
改进的Gompertz模型能够有效预测厌氧发酵的动力学强度[26]。滞后期(λ)表示微生物适应发酵环境动态变化所需的延迟响应时间[27]。随着底物煤阶的升高,滞后期也在逐渐的增加(表2),这可能是由于高阶煤中较顽固的芳构化结构。培养基中微生物的驯化需要延长的滞后期,高λ表明微生物适应反应环境的能力低,沼气和CH4的产生时间较长,说明高煤阶煤制生物甲烷会明显延长厌氧发酵时间。然而,HM显示最低的λ,但理论甲烷总产量较CYM低19.30%,结合KOCH等[28]的结论,这可能的原因是由于VFA积累,使得pH下降,进而破坏了产甲烷菌活性。

表2 生物甲烷产气潜力预测
2.3 代谢功能的差异特征
在不同煤阶煤的厌氧发酵产甲烷过程中,CYM参与微生物代谢功能的相对丰度明显高于其他试验组(图2),这意味着以长焰煤作为发酵底物,可明显改善有机物如碳水化合物、氨基酸和其他化合物的生物转化。众所周知,厌氧发酵过程中碳水化合物和氨基酸被生物代谢成挥发性脂肪酸是重要的且必经的决定步骤,因此,微生物代谢功能的增强与高挥发性脂肪酸的产生是一致的。
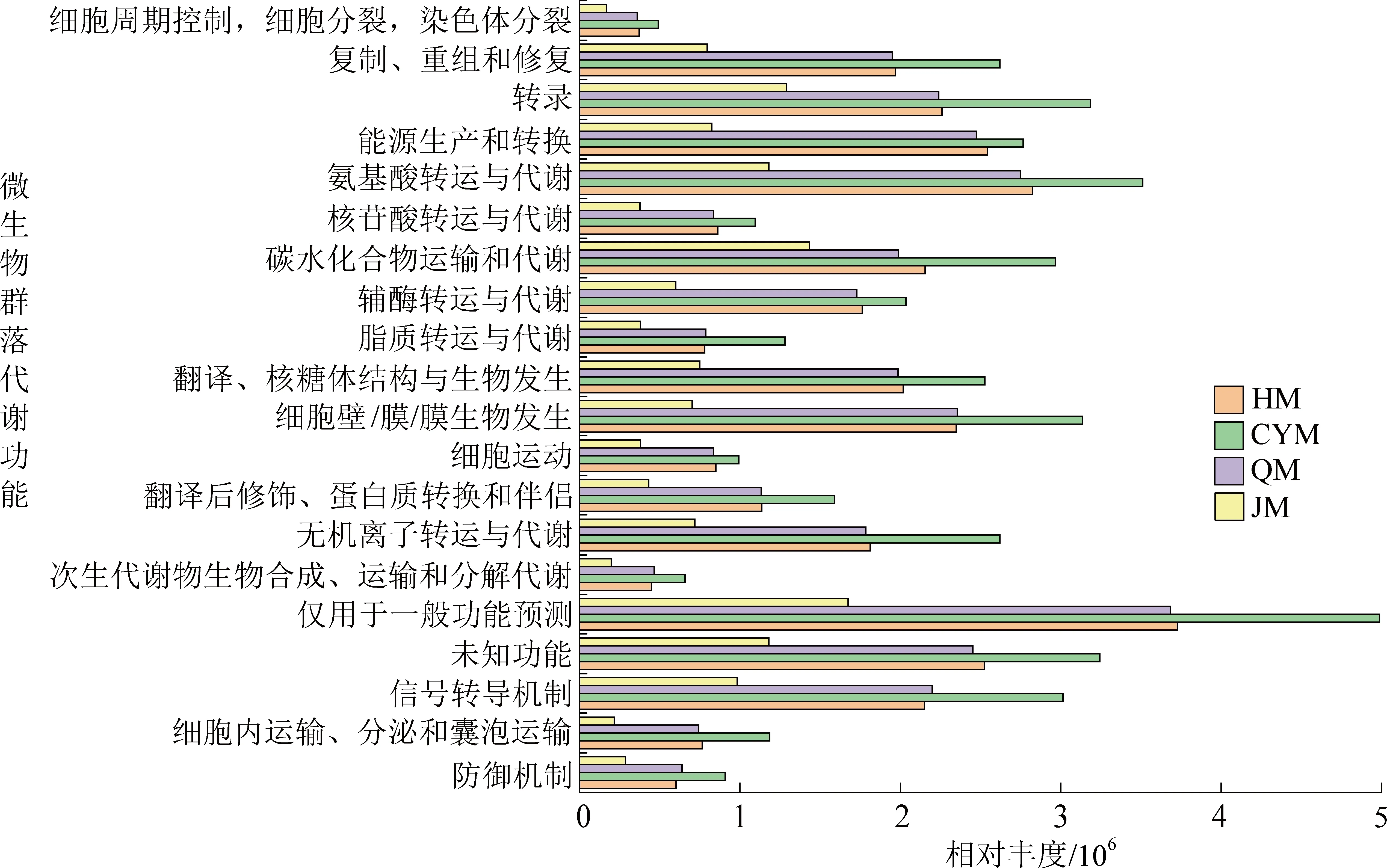
图2 基于KEGG数据库的微生物群落代谢功能
研究分析了主要参与甲烷生成的4种代谢功能(次生代谢物生物合成、运输和分解代谢;辅酶转运与代谢;碳水化合物运输和代谢;氨基酸转运与代谢)丰度与最大甲烷生成潜力之间的线性关系(图3)。从图3可以看出,参与辅酶转运与代谢和碳水化合物运输和代谢这2种代谢功能与最大甲烷生成潜力之间存在良好的线性关系(R2=0.727,R2=0.945)。为了更准确地反映这些代谢功能的差异性,对其相关的表达基因进行了对比分析。
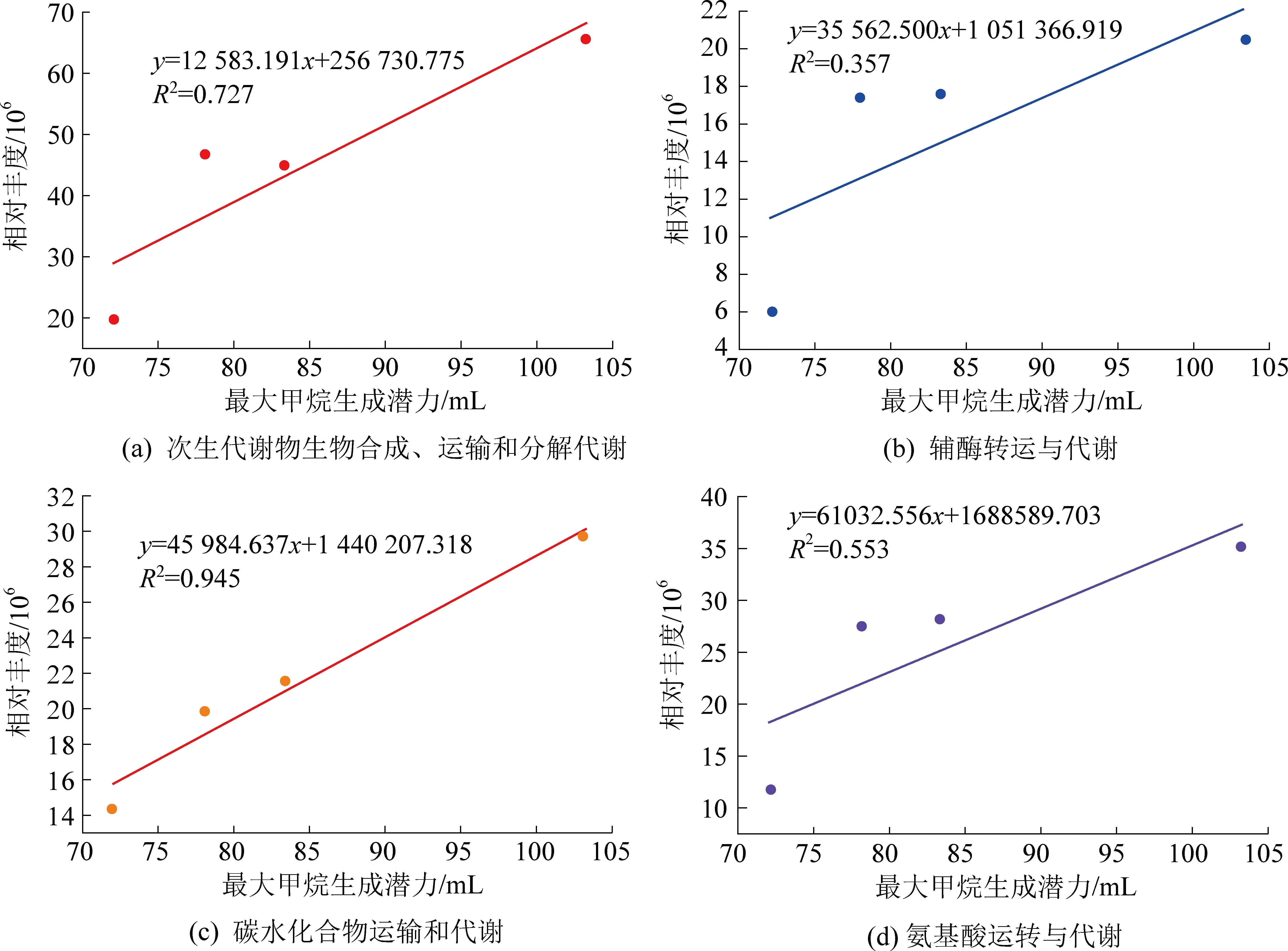
图3 代谢功能与最大甲烷生成潜力之间的线性关系
2.3.1 氨基酸代谢
通常氨基酸的代谢是指其形成和转化。因此,通过根据氨基酸的形成和转化相关基因的相对丰度来进行预测分析,相关功能基因描述见表3,分析对比结果如图4所示。

表3 氨基酸代谢相关功能基因描述
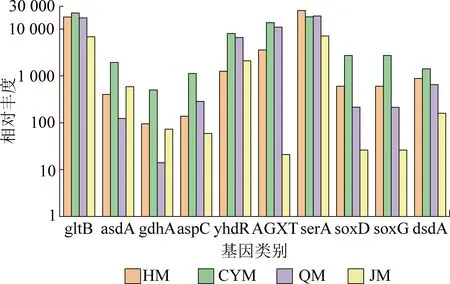
图4 氨基酸形成与转化相关功能基因丰度对比
不同煤阶煤发酵过程中与氨基酸形成相关的酶的基因丰度差异较大(图4)。CYM中参与谷氨酸合成的gltB的相对丰度最高[29]。此外,CYM中能够将含氮基团转移到L-丙氨酸的asdA[30]的相对丰度比HM、QM和JM分别高15.69倍、4.79倍、3.27倍。另一方面,与氨基酸转化相关的酶的基因在不同底物条件下也发生了变化。gdhA基因在4组试验组中的整体丰度较低,该基因与谷氨酸脱氢酶有关[31],谷氨酸脱氢酶能将谷氨酸脱氢酶转化为α-酮戊二酸,这有利于丙酮酸合成,其整体变化趋势与甲烷产量成正比关系。CYM中较高的编码天冬氨酸氨基转移酶和参与草酰乙酸合成的yhdR基因和aspC基因丰度,有利于乙酸和丙酸的产生,为产甲烷菌提供更充足的营养源。CYM中同时也存在较高丰度的AGXT基因(参与丙酮酸的直接合成),为VFA和H2/CO2的生产提供了更多的发酵前体物。
如图4所示,对氧化还原酶和磷酸甘油酸变位酶(糖酵解酶)至关重要的以及有助于丝氨酸形成的serA基因相对丰度与煤阶几乎成反比关系,即煤阶越低,则serA基因丰度越高,这可能与低阶煤中含有较高残余糖类物质有关。在CYM中有较丰富的soxD基因和soxG基因,其编码与甘氨酸形成相关的氧化还原酶,并作用于供体化合物的氨氮基团。显然,这2个基因的丰度与甲烷测试结果成正相关关系。此外,编码能够将D-丝氨酸水解为丙酮酸的D-丝氨酸脱水酶的dsdA基因丰度在CYM中有强的活跃性,进而促进发酵体系中脂肪酸的生成。
2.3.2 脂肪酸生物合成
在有机物厌氧发酵过程中,脂肪酸生物合成途径与VFA的形成密切相关。因此,进一步分析了脂肪酸生物合成途径中与VFAs产生相关的功能基因,相关功能基因描述见表4。fabG基因在整体功能占比最大,该基因已被报道与酰基还原酶的编码有关,酰基还原酶以烟酰胺腺嘌呤二核苷酸(NAD+)或烟酰胺腺嘌呤二核苷酸磷酸(NADP+)作为电子受体[32],并且在脂质代谢和脂肪酸合成中起重要作用。

表4 脂肪酸生物合成相关功能基因描述
乙酰辅酶a羧化酶对应于4个同源基因accA、accB、accC、accD和bccA。其中,accA、accB、accC、accD的相对丰度处于较高水平,并且在CYM中的相对丰度高于其他试验组(图5)。KRIVORUCHKO等[33]得出,碳水化合物、蛋白质和脂肪等主要底物被乙酰辅酶a通过常见的代谢途径降解,它也是脂肪酸生产的前体物质。accA、accB、accC和accD参与了代谢模块中的碳水化合物代谢、能量代谢和脂质代谢,accA和accD可以生成C—C键,而accC可以在化学连接模块中生成C—C和C—H键。然而,与其他乙酰辅酶a羧化酶相比,bccA的相对丰度较低,它们在煤代谢过程中的贡献率相对较低,可以推断底物煤的加入可能会破坏这2种基因的表达。
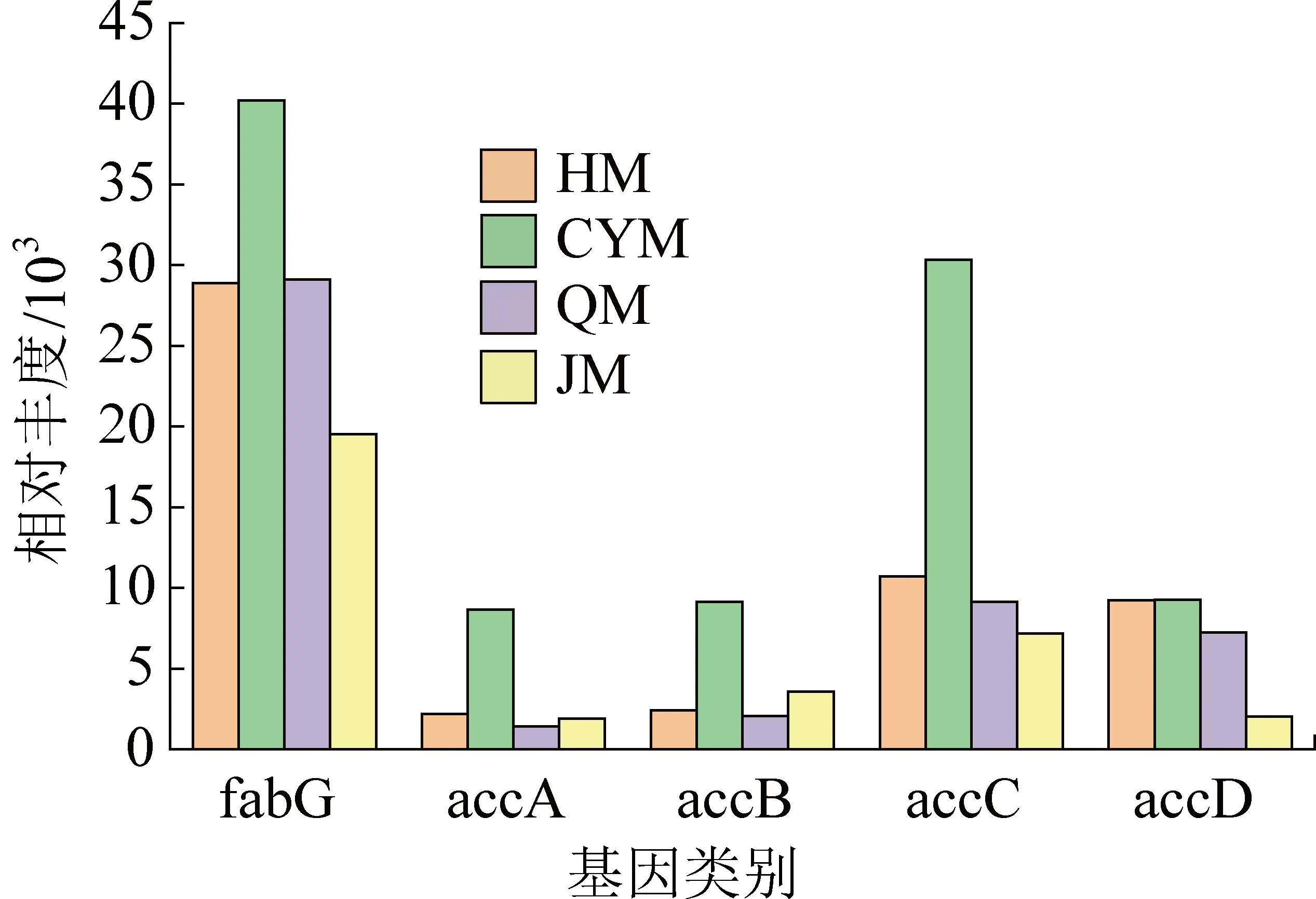
图5 脂肪酸生物合成相关功能基因丰度对比
2.3.3 细胞膜合成
此外,在CYM中的群体感应基因的相对丰度最高。群体感应是一种调节系统,可以展示细菌的细胞密度信息,并表达相应的调节基因。群体调控包括毒力因子产生、抗生素合成、细胞运动、孢子形成和生物膜形成[34-35]。本研究中,与胞外多糖合成和生物膜形成相关的crp基因[36]相对丰度在CYM中显著增加(图6),该基因描述见表5。生物膜的形成对于保护细胞免受外界侵袭,提高细胞对环境压力的抵抗力和耐受性非常重要,同时更多的胞外多糖表明有更多的底物为产酸细菌提供了产生挥发性脂肪酸的途径,这也是CYM中甲烷产量较多的主要原因之一。
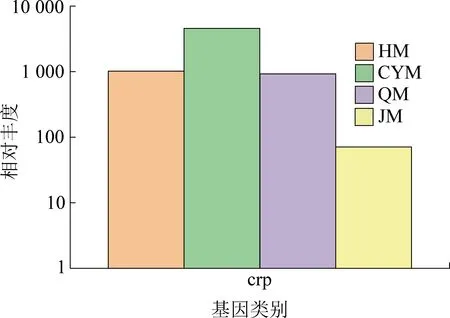
图6 生物膜合成相关功能基因丰度对比

表5 细胞膜合成相关功能基因描述
2.3.4 不同煤阶煤制生物甲烷代谢功能的差异讨论
在酶水解过程中,水解厌氧菌和兼性细菌,如梭状芽孢杆菌、肠杆菌和链球菌[37],产生胞外水解酶(糖苷酶、肽酶或酯酶)[38],它们破坏煤的大分子基质[39],释放蛋白质、脂质、核酸和较小的水溶性有机单体(糖、肽、氨基酸、长链脂肪酸和甘油酯)。
随着煤阶的降低,煤中的木质素含量会呈现增加的趋势,然后木质素因其在厌氧条件下不可降解性(归因于胞外酶所需氧气的缺乏),降低了木质纤维素的比表面积,进而抑制其它有机组分的生物降解以及CH4形成[40-41]。褐煤中的木质素含量较其他煤阶有所增加,无法为厌氧微生物提供更充足的生存空间,使得产生的相关基因功能酶有所降低。另外,木质素分解产物,如酚醛和酚酸,也可能对产甲烷菌有毒[42]。
有研究证实,底物中所含有的可溶性Ca2+离子可占据特定的木质素表面官能团,防止了胞外酶吸附到木质素上,进而增加了厌氧发酵系统的高效性和稳定性[43-44]。褐煤中降低的Ca2+与较高的木质素含量,使得胞外酶更多的吸附于木质素表面,从而降低了胞外酶的生物利用性,这是褐煤煤制生物甲烷产量以及检测的代谢相关酶含量低于长焰煤的原因之一。
3 结 论
1)以长焰煤为底物的生物甲烷试验出现不随煤阶增加而产气总量减少的结果,其生物甲烷产量也呈现最大值。
2)随着底物煤阶的升高,滞后期也在逐渐的增加,即高煤阶煤制生物甲烷实验表现出较低的微生物适应反应环境的能力。
3)高的煤制生物甲烷产量,其参与微生物代谢功能的相对丰度也最高,且参与辅酶转运与代谢和碳水化合物运输和代谢这2种代谢功能与最大甲烷生成潜力之间存在良好的线性关系。
4)生物甲烷产量越高,发酵体系中参与氨基酸形成与转化、脂肪酸和生物膜合成相关的功能基因丰度也会提高。
