A New Species of Odorous Frog Genus Odorrana (Anura,Ranidae) from Southern Guizhou Province,China
2021-12-30TaoLUOSiweiWANGNingXIAOYaliWANGandJiangZHOU
Tao LUO ,Siwei WANG ,Ning XIAO ,Yali WANG and Jiang ZHOU
1 School of Karst Science,Guizhou Normal University,Guiyang 550001,Guizhou,China
2 School of Life Sciences,Guizhou Normal University,Guiyang 550001,Guizhou,China
3 Guiyang Nursing Vocational College,Guiyang 550003,Guizhou,China
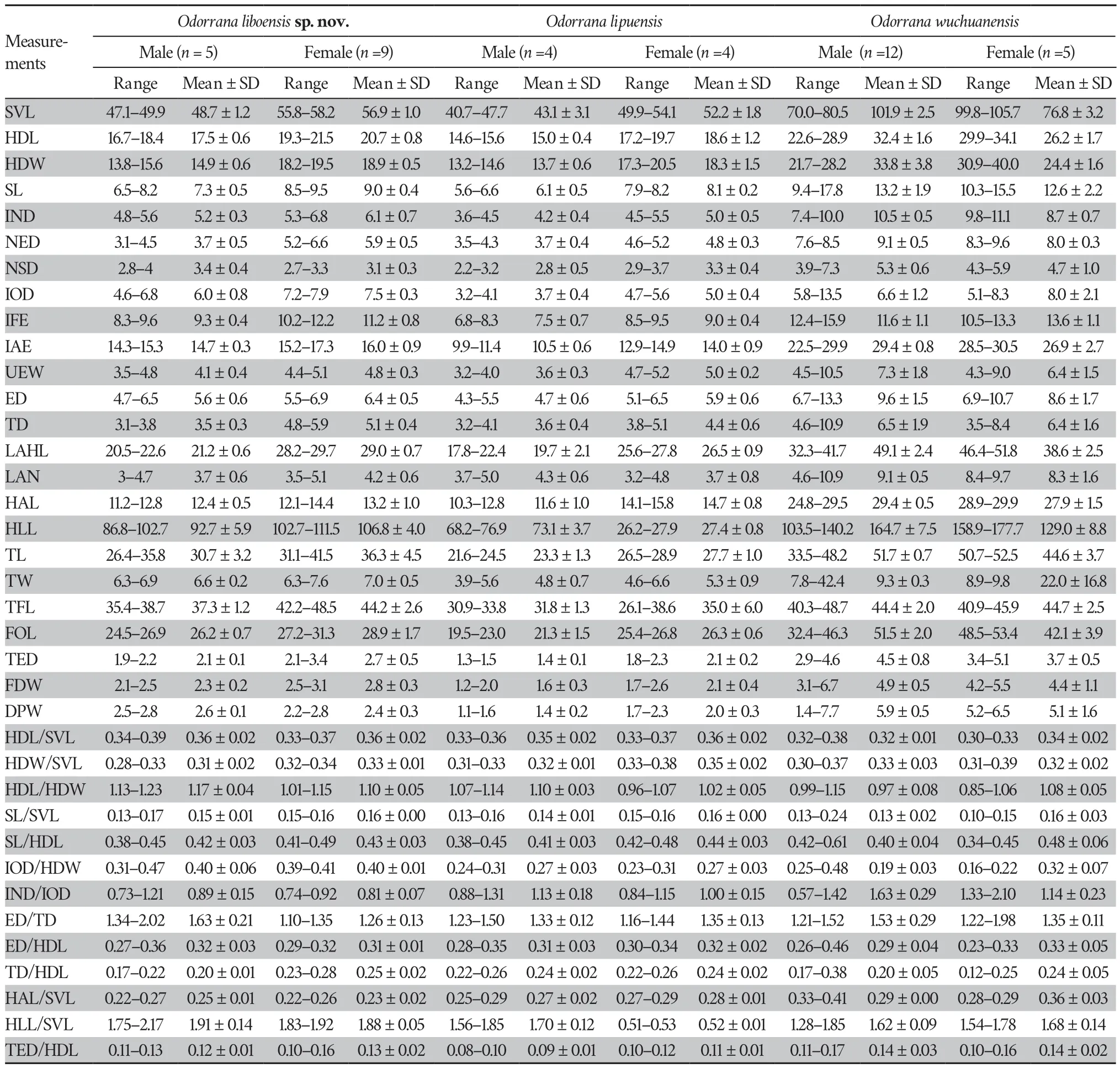
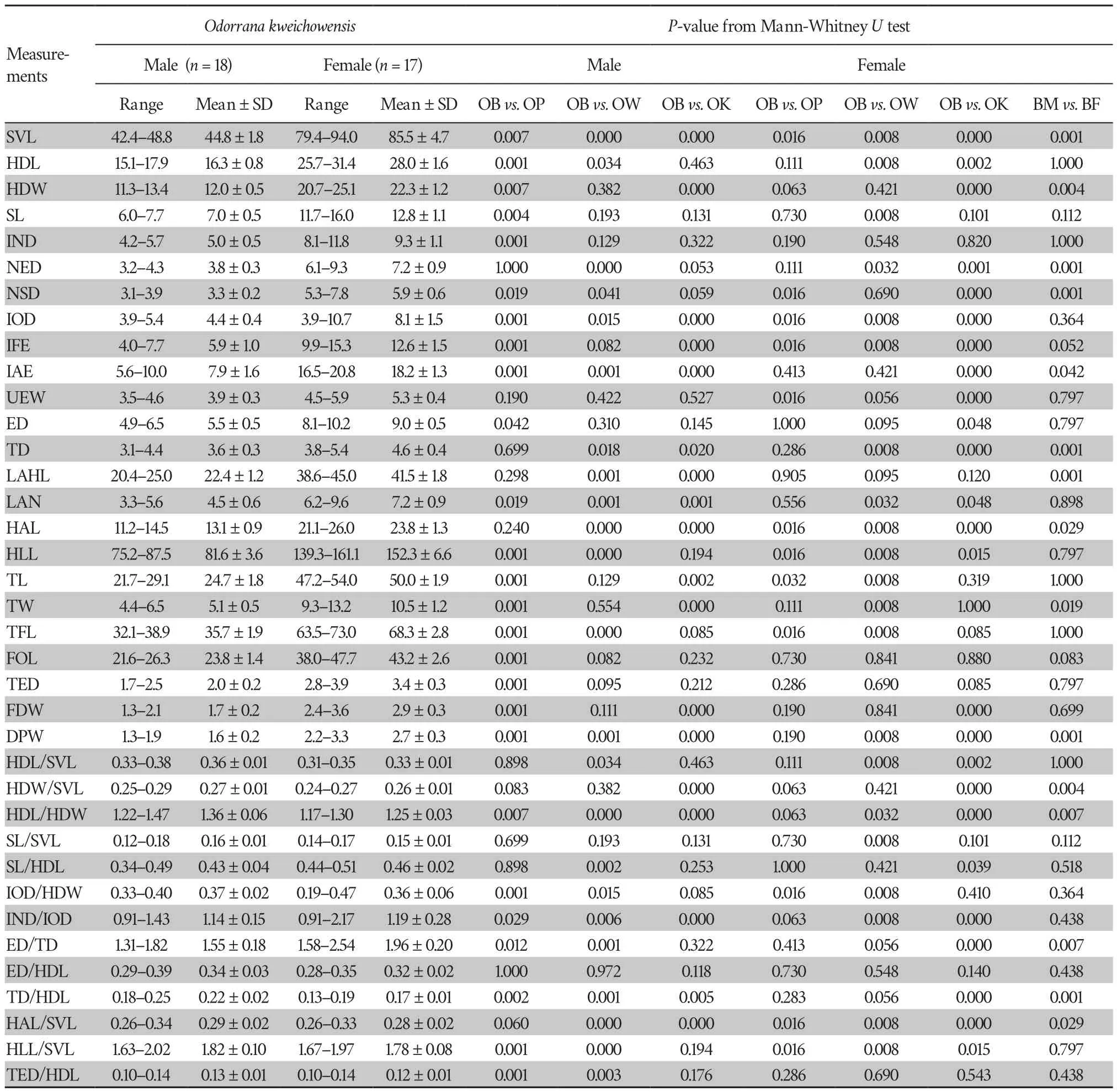
Abstract We describe Odorrana liboensis sp.nov., a new species from the Maolan National Nature Reserve,Libo County,Guizhou Province,China.Phylogenetic analyses based on DNA sequences of the mitochondrial 12S rRNA,16S rRNA,and ND2 genes supported the new species as an independent lineage,closely related to O.lipuensis.The uncorrected genetic distances between the 12S rRNA and 16S rRNA in the new species and its closest congener, O.lipuensis, were 6.06% and 5.19%,respectively.The new species is distinguished from its congeners by a combination of the following morphological characters:(1) having medium body size,with the snout-vent length (SVL) of adult females approximately 1.2 times as long as males at 56.9 ±1.0 (55.8–58.2 mm,n=9) in females and 48.7 ± 1.2 (47.1–49.9 mm,n=5) in males;(2) head length greater than width in males and females;(3) tympanum distinctly visible,greater than one-half the diameter of the eye;(4) eyes big and prominent,width of upper eyelid (UEW) approximately 3/4 of eye diameter (ED);(5) dorsolateral folds absent;(6)two metacarpal tubercles;(7) relative finger lengths:II
Keywords Taxonomy,morphology,Odorrana,Odorrana liboensis sp.nov., karst cave,Guizhou
1.Introduction
The genusOdorrana
was first recognized by Feiet al
.(1990)with the type speciesOdorrana margaretae
(Liu,1950).Despite previous controversy over classification (Dubois,1992;Matsui,1994),molecular phylogenetic analyses suggested thatOdorrana
forms a well-supported monophyletic group (Matsuiet al
.,2005;Ngoet al
.,2006;Caiet al.
,2007;Cheet al
.,2007;Stuart,2008;Chenet al
.,2013;Liet al
.,2018a).According to Frost (2021),Odorrana
contains at least 61 recognized species and is widely distributed in montane streams and rivers in the subtropical and tropical regions of East and Southeast Asia (Feiet al
.,2012;AmphibiaChina,2021;Frost,2021).The range of species in this genus covers the Ryukyu Archipelago,southern China,northeastern India,and the Thai-Malay Peninsula,and extends southwards to Sumatra and Borneo (Frost,2021).All species in the genus are associated with mountain streams except for
O.wuchuanensis
(Xu,1983) andO.lipuensis
Mo,Chen,Wu,Zhang &Zhou,2015,which occur in dark caves.Among theOdorrana
species,39 occur in China and 24 of these are endemic to China (Feiet al
.,2012;AmphibiaChina,2021;Frost,2021;Chen
et al
.,2020;Shen
et al
.,2020;Liu
et al
.,2021).Feiet al
.(1990) establishedOdorrana
,and later Ye and Fei (2001) suggested four species groups (O.andersonii
,O.kuangwuensis
,O.schmacheri
,and
O.livida
) withinOdorrana
based on a morphological phylogenetic study.Feiet al
.(2005)established two subgenera (Odorrana
andBamburana
) withinOdorrana
and recognizedOdorrana versabilis
as the type species ofBamburana
.Molecular phylogenetic studies support the division of species groups withinOdorrana
but not the two subgenera (Cheet al
.,2007).Subsequently,Fei
et al
.(2009) divided the ChineseOdorrana
species into six species groups (O.tormota
,O.taiwaniana
,O.graminea
,O.margaretae
,O.schmackeri
,andO.andersonii
;O.taiwaniana
is a synonym forO.swinhoana
) based on morphological characters.These divisions have been accepted by other researchers and applied to theOdorrana
species distributed outside China (Phamet al
.,2016;Liet al
.,2018a).However,recent studies have rejected the monophyly of theO.margaretae
,O.schmackeri
,andO.andersonii
species groups (Chenet al
.,2013).The phylogenetic relationships between these species groups,the species included in the species groups,and the species group affiliations of new species published in recent years are unclear.The montane river and stream habitats of mostOdorrana
may promote intraspecific divergence through geographical isolation,especially given the wide geographical range of the complex (Chenet al
.,2013;Liet al
.,2015).Indeed,many cryptic species ofOdorrana
have been discovered.For example,25 new species ofOdorrana
have been described since 2005 (See species list of the genusOdorrana
in Frost(2021)and AmphibiaChina(2021)).This indicates that further discoveries or potential taxonomic changes withinOdorrana
are likely.From 2016 to 2018,14 specimens of an unknown species ofOdorrana
were collected inside a completely dark karst cave in the Maolan National Nature Reserve,Libo County,Guizhou Province,China.Morphologically,these specimens most closely resembleO.lipuensis
andO.kweichowensis
Li,Xu,Lv,Jiang,Wei &Wang,2018 (Moet al
.,2015;Liet al
.,2018a),but differ fromO.lipuensis
,O.kweichowensis
,and all otherOdorrana
from China and adjoining countries.They inhabit a dark karst cave environment similar to that ofO.
wuchuanensis
andO.lipuensis
but are morphologically very different fromO.
wuchuanensis
andO.lipuensis
.To distinguish these specimens,we conducted phylogenetic analyses based on mitochondrial DNA and morphological comparisons.All of the analyses consistently indicated that the specimens from Maolan National Nature Reserve are a new taxon.We describe this taxon here as a new species.2.Materials and Methods
2.1.Sampling
A total of 72 specimens were collected in this study.Fourteen specimens were of the undescribed species from Maolan National Nature Reserve in Libo County,Guizhou Province;35 wereO.kweichowensis
from Lengshuihe Nature Reserve in Jinsha County,Guizhou Province;four wereO.yizhangensis
Fei,Ye &Jiang,2007 from Kuankoushui National Nature Reserve in Suiyang County,Guizhou Province,and Yuntai Mountain,Shibing County;one wasO.huanggangensis
Chen,Zhou &Zheng,2010 from Yueliangshan Nature Reserve in Conjiang County,Guizhou Province;one wasO.kweichowensis
from Maolan National Nature Reserve in Libo County,Guizhou Province;and 17 wereO.wuchuanensis
from Maolan National Nature Reserve,Libo County,Guizhou Province (Figure 1).All of the specimens were fixed in 10%buffered formalin and later transferred to 75% ethanol for preservation.The muscles used for molecular analysis were preserved in 95% alcohol at −20 °C.All of the specimens were deposited in Guizhou Normal University (GZNU),Guiyang City,Guizhou Province,China.2.2.DNA Extraction,PCR amplification,and sequencing
Genomic DNA was extracted from muscular tissue using a DNA extraction kit from Tiangen Biotech Co.,Ltd.(Beijing).All samples were sequenced for three mitochondrial genes:partial 12S ribosomal RNA gene (12S rRNA),16S ribosomal RNA gene (16S rRNA),and NADH dehydrogenase subunit 2 (ND2).The primers used for 12S rRNA were 12SF (5’-GGTTTGRTCCTRGCCTTAC-3’)and 12SR (5’-CCATGTTACGACTTGCCTCT-3’) following Chenet al
.(2013),the primers used for 16S rRNA were 16SF (5’-ACGAGCCTAGTGATAGCTGGTT-3’) and 16SR (5’-CGGTCTGAACTCAGATCACGT-3’) following Chenet al
.(2013),and the primers used for ND2 were Gln-LND2 (5’-CCCTTTGCACTTCCTTTATGC-3’) and Ala-HND2 (5’-GGCCTGAGTTGCATTCATG-3’) following Liet al
.(2015).PCR amplifications were performed in a 25 μL reaction volume with the following cycling conditions:an initial denaturing step at 95 °C for five min;36 cycles of denaturing at 95 °C for 40 s,annealing at 52 °C (for 12S rRNA)/49 °C (for 16S rRNA)/60 °C (for ND2) for 40 s and extending at 72 °C for 1 min,and a final extending step of 72 °C for 10 min.The PCR products were sequenced on an ABI Prism 3730 automated DNA sequencer in Chengdu TSING KE Biological Technology Co.Ltd.(Chengdu,China).All of the sequences have been deposited in GenBank (Table 1).Some homologous DNA sequences of voucher specimens of related species were downloaded from GenBank and incorporated into the phylogenetic analyses.2.3.Phylogenetic analyses
We used a total of 187 sequences (including 72 12S rRNA,79 16S rRNA,and 36 ND2 gene sequences) for molecular analyses.Three mitochondrial genes from 11 muscle tissues were sequenced and 154 sequences downloaded from GenBank from 48 species of the genusOdorrana
were used.These included the undescribed species from China that was named in this study (Figure 1).A total of 26 sequences were downloaded from GenBank as out-groups(Chenet al
.,2013).Detailed information on these materials is shown in Table 1.All of the sequences were assembled and aligned using the MUSCLE (Edgar,2004) module in MEGA 7.0 (Kumaret al
.,2016)with default settings.Alignments were checked by eye and revised manually if necessary.Trimming with the gaps partially deleted was performed in GBLOCKS 0.91b (Castresana,2000).Phylogenetic trees were constructed with both maximum likelihood (ML) and Bayesian inference (BI).The ML was conducted in IQ-TREE(Nguyenet al
.,2015) with 2000 ultrafast bootstrapping (Hoanget al
.,2018) and was performed until a correlation coefficient of at least 0.99 was reached.The BI was performed in MrBayes 3.2.1 (Ronquistet al
.,2012),and the best-fit model was obtained by the Bayesian inference criteria (BIC) computed with PartitionFinder 2 (Lanfearet al
.,2016).For this analysis,12S rRNA,16S rRNA,and ND2 genes were defined.The analysis suggested that the best partition scheme was 12S rRNA/16S rRNA/ND2 genes.We selected GTR+I+G as the best model for 12S rRNA and 16S rRNA and the TIM+I+G model as the best model for the ND2 gene.Two independent runs were conducted in BI analysis,each of which was performed for 2,000,000 generations and sampled every 1000 generations.The first 25% of the samples were discarded as burn-in,resulting in a potential scale reduction factor (PSRF) of <0.01.Nodes in the trees were considered well supported when Bayesian posterior probabilities (BPP) were ≥0.95 and the ML ultrafast bootstrap value(UFB) was ≥95%.Uncorrectedp
-distances (1000 replicates) based on 12S rRNA and 16S rRNA were calculated in MEGA 7.0 (Kumaret al
.,2016).
Figure 1 Sampling collection localities and distribution of the Odorrana liboensis sp.nov.,O.lipuensis,O.wuchuanensis,O.kweichowensis,O.huanggangensis,and O.yizhangensis in southwest China.1.Maolan National Nature Reserve,Libo County,Guizhou,China.2.Lipu County,Guangxi,China.3.Jiarong Town,Libo County,Guizhou,China.4.Maolan National Nature Reserve,Libo County,Guizhou,China.5.Lengshuihe Nature Reserve,Jinsha County,Guizhou,China.6.Yueliangshan Nature Reserve,Congjiang County,Guizhou,China.7.Yuntai Mountains,Shibing County,Guizhou,China.8.Kuankuoshui National Nature Reserve,Suiyang County,Guizhou,China.The base maps are from Standard Map Service website (http://bzdt.ch.mnr.gov.cn/index.html).
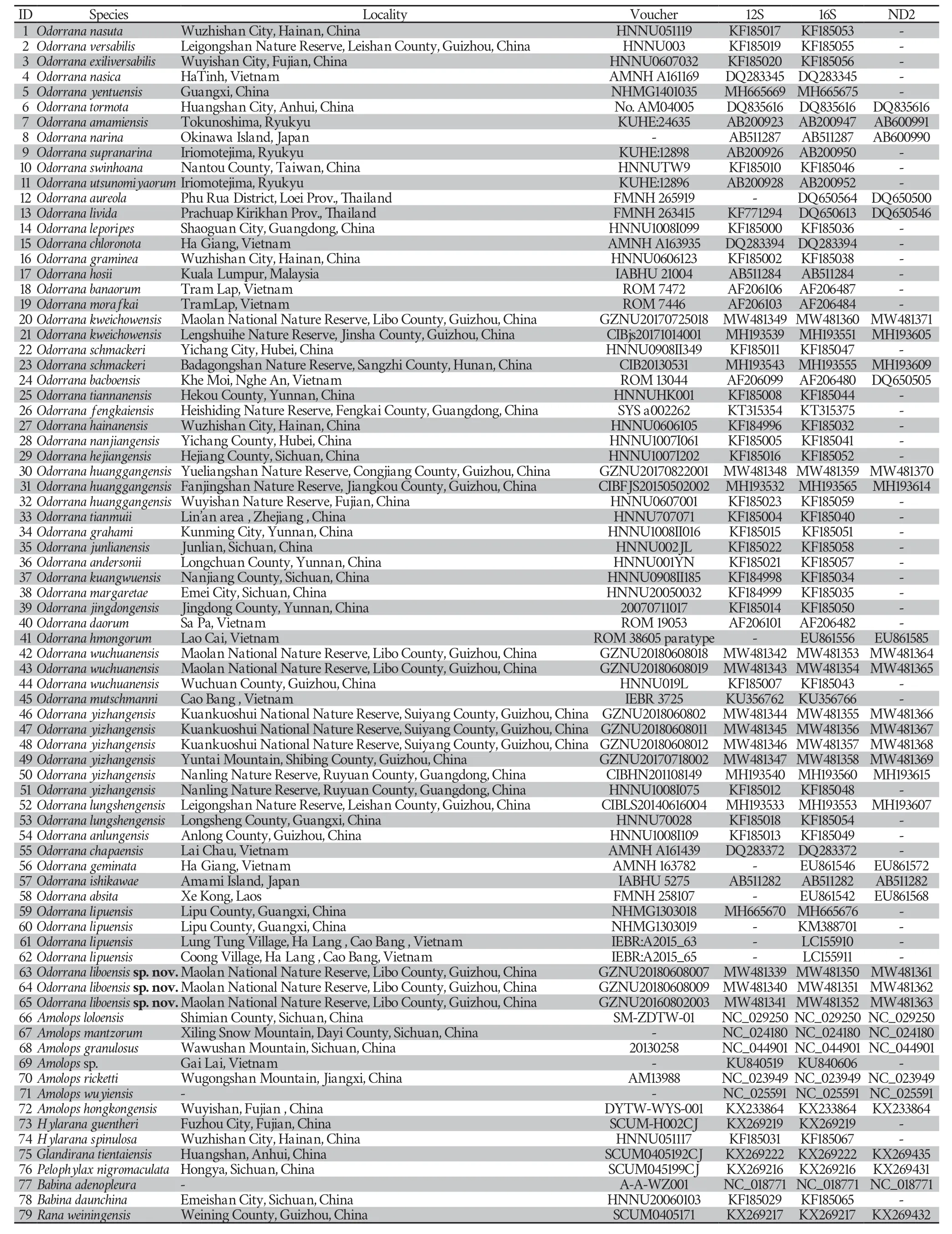
Table 1 Localities,voucher information,and GenBank numbers for all samples used in this study.
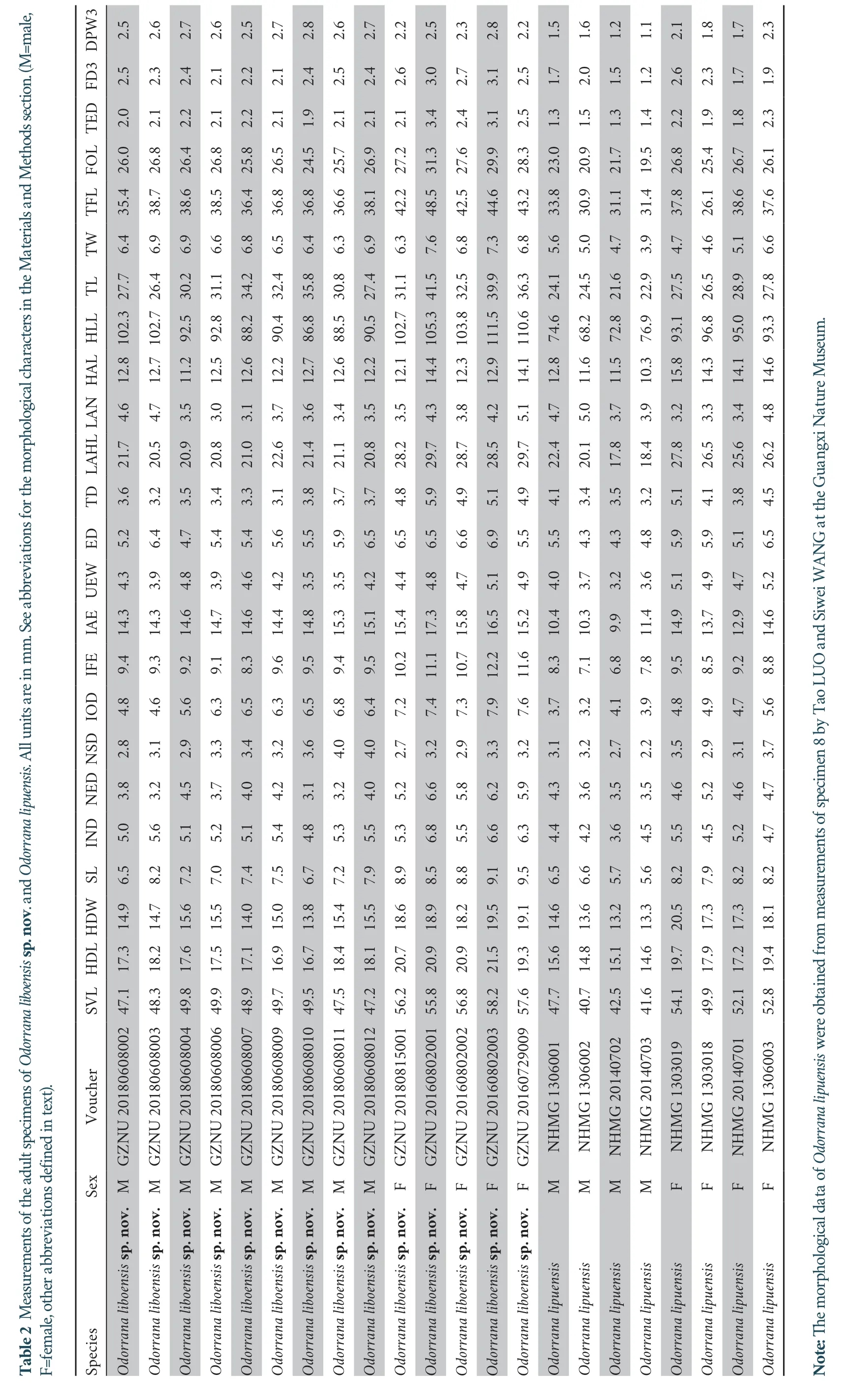
2.4.Morphological analysis
Morphometric data were taken from 74 well-preserved adult specimens (voucher information in Table 2 and Table S1).Measurements were recorded to the nearest 0.1 mm with digital calipers by Tao Luo following Feiet al
.(2009) and Liet al
.(2018a).A total of 24 morphological characteristics were measured.These measurements were as follows:SVL
=snout-vent length(distance from the tip of the snout to the posterior edge of the vent),HDL
=head length (distance from the tip of the snout to the articulation of jaw),HDW
=maximum head width (greatest width between the left and right articulations of jaw),SL
=snout length(distance from the tip of the snout to the anterior corner of the eye),IND
=internasal distance (minimum distance between the inner margins of the external nares),NED
=nasal to eye distance(distance between the nasal and the anterior corner of the eye),NSD
=nasal to snout distance (distance between the nasal the posterior edge of the vent),IOD
=interorbital distance (minimum distance between the inner edges of the upper eyelids),IFE
=distance between anterior corner of eye,IAE
=distance between posterior corner of eyes,UEW
=upper eyelid width (greatest width of the upper eyelid margins measured perpendicular to the anterior-posterior axis),ED
=eye diameter (distance from the anterior corner to the posterior corner of the eye),TD
=tympanum diameter,LAHL
=length of lower arm and hand (distance from the elbow to the distal end of the finger IV),LAN
=width of lower arm,HAL
=hand length(distance from the posterior end of the inner metacarpal tubercle to the distal tip of Finger IV),HLL=
hindlimb length (maximum length from the vent to the distal tip of the toe IV),TL=
tibia length (distance from knee to tarsus),TW=
maximal tibia width,TFL=
length of foot and tarsus (distance from the tibiotarsal articulation to the distal end of the toe IV),FOL=
foot length (from the base of inner metatarsal tubercle to the tip of fourth toe),TED
=tympanum-eye distance (from anterior edge of tympanum to posterior corner of the eye),FDW
=finger disk width (width at the widest part of the disk of finger III),andDPW
=distal phalanx width (maximal width of the distal phalanx of finger III).To reduce the impact of allometry,a size-corrected value from the ratio of each character to SVL was calculated for the following morphometric analyses.Principal component analysis (PCAs) of size-corrected variables and simple bivariate scatterplots was used to explore and reflect the morphometric differences between the new species andO.lipuensis
.Mann-WhitneyU
tests were used to test the significance of differences on morphometric characters between the new species andO.lipuensis
,O.kweichowensis
andO.wuchuanensis
.Mann-WhitneyU
tests were also conducted to test the morphometric differences between the males and the females of the new species.The statistical analyses were performed using SPSS 21.0 (SPSS,Inc.,Chicago.IL,USA),and differences were considered to be significant atP
<0.05.Sex was determined by direct observation of calling behavior and the presence of internal vocal sac openings for males,as well as the presence of eggs on the abdomen for females.The presence or absence of nuptial pads/spines was examined by optical microscopy.
We compared the morphological characters of the new taxon with other species ofOdorrana
.Comparative data were obtained from the literature for 61 species ofOdorrana
(Table 3).For comparison,we examined the type and/or topotype materials forO.lipuensis
,O.kweichowensis
,andO.
wuchuanensis
(Appendix I).3.Results
3.1.Phylogenetic analyses and genetic divergence
The maximum likelihood (ML) and Bayesian inference (BI)phylogenetic trees were constructed based on concatenated DNA sequences of the mitochondrial 12S rRNA (752 bp),16S rRNA (1006 bp),and ND2 (1030 bp) genes with a total length of 2788 bp.ML and BI analyses resulted in a largely identical topology (Figure 2).The new taxon is a sister taxon toO.lipuensis
with high node support values (1.00 in BI and 100% in ML;Figure 2).The smallestp
-distance divergences between the new lineage and other species ofOdorrana
were 6.06% in 12S rRNA (between new taxon andO.lipuensis
) and 3.90% in 16S rRNA (between new taxon andO.geminata
),which were at the same divergence level as those among recognized congeners (1.94% and 2.79% in 12S rRNA betweenO.
wuchuanensis
vs.O.mutschmanni
andO.kweichowensis
vs.O.schmackeri
,respectively;and 1.30% and 1.95%in 16S rRNA betweenO.lungshengensis
vs.O.yizhanggensis
andO.nanjiangensis
vs.O.hejiangensis
,respectively),indicating that the new taxon represents an independent evolutionary lineage(Tables S2–S3).3.2.Morphological analyses
The Mann-WhitneyU
tests indicated that males of the new species were significantly different fromO.lipuensis
andO.kweichowensis
in many morphometric characters (allP
values <0.05;Table 4).PCA extracted three and four principal component factors with Eigenvalues greater than two in males and females,respectively(Table S4).The first two principal components explained 61.34% and 63.28%,of the total variation in males and females,respectively.These differences were mainly influenced by limb and head characteristics (Table S4).The new taxon was distinctly separated fromO.lipuensis
andO.kweichowensis
on the two-dimensional plots of PC1vs.
PC2,in both males and females(Figure 3).3.3.Taxonomic account
sp.nov.(Tables 2 and 4,Figures 4–5)
Holotype.
GZNU20180608004 (Figure 4),adult male,collected by Tao LUO on June 8,2018 from Maolan National Nature Reserve (25.481711° N,108.078003° E,ca.715 m a.s.l.),Libo County,Guizhou Province,China.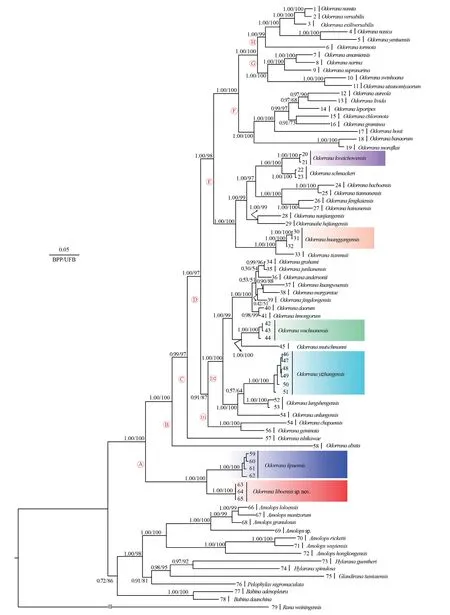
Figure 2 Phylogenetic tree based on mitochondrial 12S rRNA+16S rRNA+ND2 genes.In the phylogenetic tree,ultrafast bootstrap supports (UFB) from ML analyses and Bayesian posterior probabilities (BPP) from BI analyses were noted beside nodes.The scale bar represents 0.05 nucleotide substitutions per site.Numbers at the tips of branches correspond to the ID numbers in Table 1.
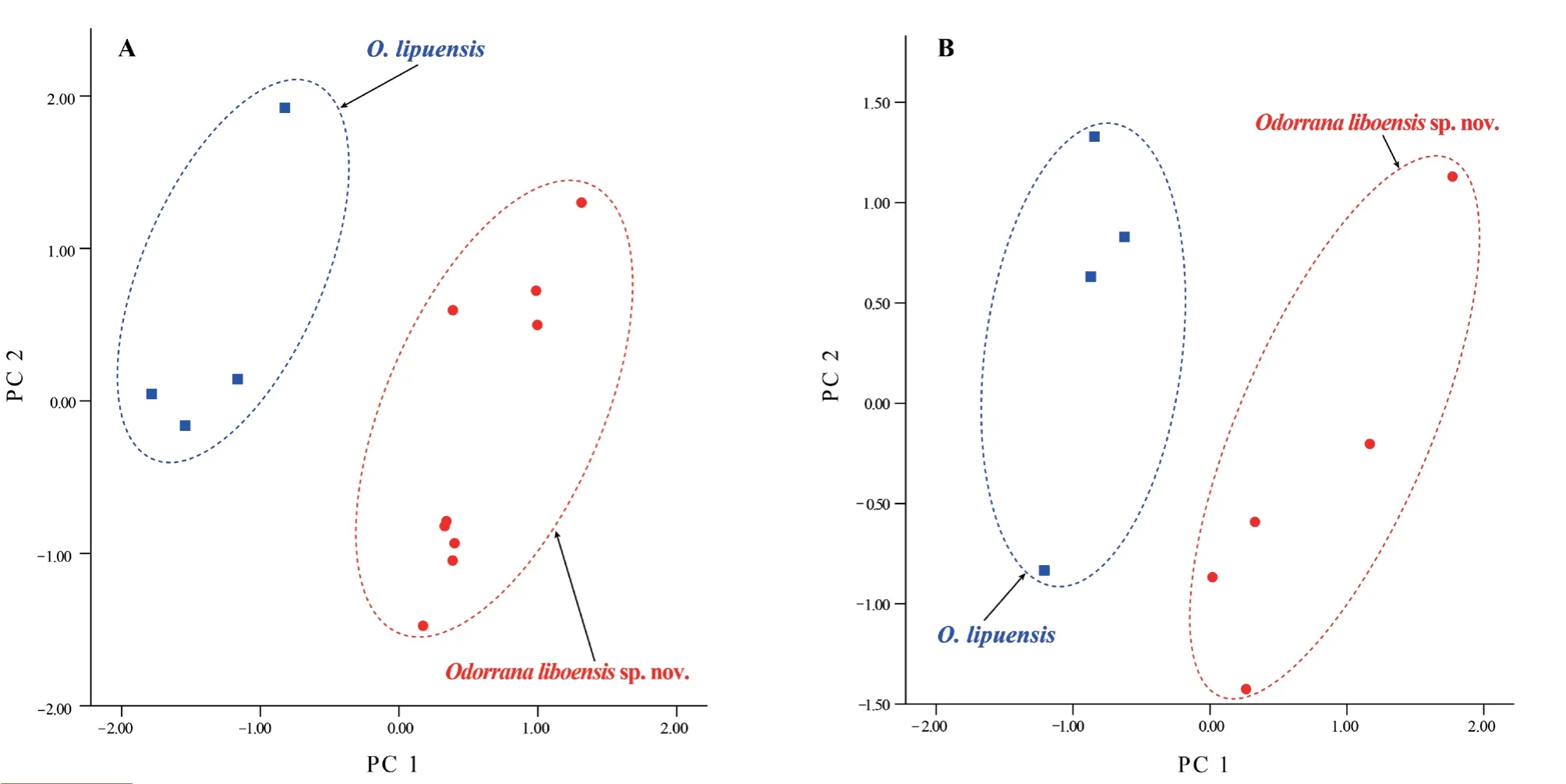
Figure 3 Plots of the first principal component (PC 1) versus the second (PC 2) for Odorrana liboensis sp.nov.and O.lipuensis from a principal component analysis.A,male.B,female.
Paratypes.
Thirteen individuals collected at the same locality as the holotype.GZNU20180608002,GZNU20180608003,GZNU20180608006,GZNU20180608007,GZNU20180608009,GZNU20180608010,GZNU20180608011 and GZNU20180608012,adult males collected by Tao Luo on June 8;GZNU20180815001(Figure 5),adult females collected by Xiang ZENG on August 15,2018;GZNU20160802001,GZNU20160802002,GZNU20160802003,and GZNU20160729009,adult females collected by Tao Luo on August 2,2016.Etymology.
The specific epithet“liboensis
”is in reference to the type locality of the new species:Libo County,Guizhou Province,China.We propose the common English name“Libo Odorous Frog”and Chinese name“Li Bo Chou Wa (荔波臭蛙)”.Differential diagnosis.
Odorrana liboensis
sp.nov.
is assigned to genusOdorrana
based upon molecular phylogenetic analyses and the following morphological characters:(1) dorsum is mostly green;(2) tips of digits dilated,tapering,disks with circum-marginal grooves or lateroventral grooves,and vertical diameter longer than horizontal diameter in the disks;(3)supernumerary tubercle below the base of fingers III and IV;(4) feet fully webbed to disks,without tarsal fold;(5) the first finger thick and nuptial pad distinct and (6) dorsal skin smooth,dorsolateral folds absent or fine (Feiet al
.,2009;Liet al
.,2018a).Odorrana liboensis
sp.nov.
is distinguished from congeners by a combination of the following characters:(1) having medium body size,with the snout-vent length (SVL) of adult females approximately 1.2 times as long as that of males at 56.9 ± 1.0(55.8–58.2 mm,n=
9) in females and 48.7 ± 1.2 (47.1–49.9 mm,n=
5) in males;(2) head length greater than width in males and females;(3) tympanum distinctly visible,greater than one-half the diameter of the eye;(4) eyes big and prominent,with the width of the upper eyelid (UEW) approximately 3/4 of the eye diameter (ED);(5) dorsolateral folds absent;(6) two metacarpal tubercles;(7) relative finger lengths:II Description of holotype.GZNU 20180608004 (Figure 4),adult male.Medium body size,SVL 49.8 mm;habitus slender;head length slightly larger than head width (HDL=17.6 mm;HDW=15.6 mm;HDL/HDW 1.12);snout short,rounded,and projecting beyond the lower jaw in dorsal view,longer than the diameter of the eye (SL/ED 1.30);nostril rounded,distinct,closer to the snout tip than the eye (NEL/NSL 0.52);internasal distance less than interorbital distance (IND/IOD 0.91) and greater than upper eyelid width (IND/UEW 1.06);pineal body invisible;tympanum distinct,rounded,75% of eye diameter,depressed relative to the skin of the temporal region,tympanic rim slightly elevated relative to tympanum;vomerine teethmoderately developed,on two oblique ridges;tongue cordiform,deeply notched posteriorly;and eyes large,slightly protuberant in dorsal view,eye diameter 26.70% of head length,pupils transverse;supratympanic fold absent.
Table 3 References for morphological characters for congeners of the genus Odorrana.
Forelimbs are slender and comparatively short,the length of the lower arm and hand is 41.97% of SVL;fingers slender,relative finger lengths:I Hindlimbs are slender (HLL/SVL 1.86);heels overlapping when thighs are positioned at right angles to the body;tibiotarsal articulation reaching the between eye to nostril when leg stretched forward;foot length less than tibia length(FL/TL 0.87);relative toe lengths I Figure 4 Morphological features of the live adult male holotype GZNU20180608004 of Odorrana liboensis sp.nov. and O.wuchuanensis.(A) Dorsal view;(B) Ventral view;(C) Dorsolateral view;(D) Tadpole;(E) Type locality of Odorrana liboensis sp.nov. in June,2018 (dry season);(F) Dorsolateral view of O.wuchuanensis in life. Skin is smooth on upper surfaces;venter smooth;tiny spinules on lateral body,upper edge of lid,temporal region,and anterior and posterior edge of tympanum;weak supratympanic fold from the posterior edge of the eye to the posterior edge of the tympanum;and dorsolateral fold absent. Coloration of holotype in life Preserved holotype coloration Table 4 Morphological comparison of Odorrana liboensis sp.nov.(OB),O.lipuensis (OP),O.kweichowensis (OK),and O.wuchuanensis (OW).All units are in mm.P-values are at 95% significance.Morphometric characters are explained in the methods section.BM and BF are the abbreviations for male and female from Odorrana liboensis sp.nov. (Continued Table 4) Variations. n= n= Sexual dimorphism. Comparisons. Odorrana liboensis sp.nov. Odorrana Odorrana liboensis sp.nov. O.absita O.anlungensis O.gigatympana O.huanggangensis O.khalam O.kweichowensis O.monjerai O.morafkai O.nasica O.orba O.tormota vs. Odorrana liboensis sp.nov. O.amamiensis O.andersonii O.aureola O.bacboensis O.cangyuanensis O.chapaensis O.geminata O.grahami O.hainanensis O.hosii O.ishikawae O.jingdongensis O.junlianensis O.kuangwuensis O.leporipes O.livida O.lungshengensis O.macrotympana O.margaretae O.mutschmanni O.nanjiangensis O.nasuta O.sinica O.splendida O.supranarina O.swinhoana O.tiannanensis O.trankieni O.versabilis O.wuchuanensis O.zhaoi vs. vs. Odorrana liboensis sp.nov. O.amamiensis O.andersonii O.anlungensis O.aureola O.bacboensis O.banaorum O.bolavensis O.chapaensis O.chloronota O.dulongensis O.fengkaiensis O.geminata O.grahami O.hainanensis O.hejiangensis O.hosii O.huanggangensis O.ishikawae O.jingdongensis O.junlianensis O.kuangwuensis O.kweichowensis O.livida O.lungshengensis O.macrotympana O.margaretae O.monjerai O.morafkai O.mutschmanni O.nanjiangensis O.narina O.nasuta O.orba O.rotodora O.schmackeri O.splendida O.supranarina O.swinhoana O.tianmuii O.tiannanensis O.tormota O.versabilis O.wuchuanensis O.yentuensi O.yizhangensis O.ichangensis vs. Odorrana liboensis sp.nov. O.absita O.amamiensis O.banaorum O.bolavensis O.exiliversabilis O.gigatympana O.graminea O.hosii O.indeprensa O.khalam O.leporipes O.livida O.monjerai O.narina O.nasica O.nasuta O.orba O.supranarina O.tormota O.trankieni O.utsunomiyaorum O.yentuensis O.zhaoi vs. Figure 5 Morphological features of the live adult female GZNU20180815001 of Odorrana liboensis sp.nov. (A) Dorsal view;(B) Dorsolateral view;(C)Ventral view;(D) Ventral view of right foot;(E) Ventral view of right hand;(F) The entrance habitat of the karst cave at the type locality of Odorrana liboensis sp.nov.;(G) Water outlet in the cave. Odorrana liboensis sp.nov. O.bacboensis O.jingdongensis O.lungshengensis O.margaretae O.mutschmanni O.nanjiangensis O.narina O.orba O.sinica O.swinhoana O.tormota O.yizhangensis vs. O.nasica O.nasuta vs. O.hainanensis vs. O.junlianensis vs. O.cangyuanensis O.exiliversabilis O.fengkaiensis O.gigatympana O.grahami O.graminea O.tiannanensis O.versabilis O.yentuensis O.zhaoi vs. O.amamiensis vs. O.amamiensis O.anlungensis O.huanggangensis O.kuangwuensis O.macrotympana O.wuchuanensis O.ichangensis vs. O.lipuensis O.splendida O.supranarina vs. O.rotodora O.trankieni vs. O.utsunomiyaorum vs. Odorrana liboensis sp.nov. O.andersonii O.anlungensis O.bacboensis O.bolavensis O.chapaensis O.dulongensis O.fengkaiensis O.geminata O.grahami O.hainanensis O.hejiangensis O.huanggangensis O.ishikawae O.junlianensis O.kuangwuensis O.kweichowensis O.lungshengensis O.margaretae O.mutschmanni O.nanjiangensis O.schmackeri O.splendida O.supranarina O.tianmuii O.tiannanensis O.wuchuanensis O.yizhangensis O.ichangensis vs. Odorrana liboensis sp.nov. O.absita O.amamiensis O.andersonii O.anlungensis O.aureola O.bacboensis O.banaorum O.bolavensis O.cangyuanensis O.chapaensis O.chloronota O.dulongensis O.exiliversabilis O.fengkaiensis O.geminata O.gigatympana O.grahami O.graminea O.hainanensis O.hejiangensis O.huanggangensis O.indeprensa O.ishikawae O.jingdongensis O.junlianensis O.khalam O.kweichowensis O.lungshengensis O.macrotympana O.morafkai O.nanjiangensis O.nasica O.nasuta O.orba O.sinica O.swinhoana O.tianmuii O.tiannanensis O.tormota O.trankieni O.utsunomiyaorum O.versabilis O.yentuensis O.yizhangensis O.ichangensis O.zhaoi vs. sp.nov. O.lipuensis O.dulongensis vs.

O.amamiensis
,andO.anlungensis
(vs.
IIvs.
IO.bacboensis
,O.banaorum
,O.bolavensis
,O.chloronota
,O.fengkaiensis
,O.gigatympana
,O.grahami
,O.jingdongensis
,O.junlianensis
,O.kweichowensis
,O.margaretae
,O.mutschmanni
,O.orba
,O.schmackeri
,O.swinhoana
,O.tiannanensis
,O.wuchuanensis
,andO.yentuensis
(vs.
IIvs.
IO.hainanensis
,O.hejiangensis
,O.kuangwuensis
,O.nasica
,O.nasuta
,andO.versabilis
(vs.
IIvs.
I≤II=IVO.supranarina
,andO.trankieni
(vs.
IVvs.
I=IVsp.nov.
differs fromO.andersonii
,O.fengkaiensis
,O.grahami
,O.graminea
,O.hainanensis
,O.huanggangensis
,O.jingdongensis
,O.junlianensis
,O.kweichowensis
,O.lungshengensis
,O.margaretae
,O.tianmuii
,O.yizhangensis
,andO.zhaoi
(vs.
presence of white pectoral spinules).By having two metacarpal tubercles on the base of the hand,Odorrana liboensis
sp.nov.
differs fromO.absita
,O.aureola
,andO.bacboensis
(vs.
lacking metacarpal tubercles),fromO.cangyuanensis
,O.dulongensis
,O.lungshengensis
,O.nanjiangensis
,O.orba
,O.schmackeri
,O.tiannanensis
,andO.yizhangensis
(vs.
one metacarpal tubercle),fromO.andersonii
,O.exiliversabilis
,O.fengkaiensis
,O.hainanensis
,O.hejiangensis
,O.huanggangensis
,O.jingdongensis
,O.junlianensis
,O.nasica
,O.nasuta
,O.tianmuii
,O.tormota
,andO.ichangensis
(vs.
three metacarpal tubercles).By having one metacarpal tubercle on the base of the metatarsus,Odorrana liboensis
sp.nov.
differs fromO.absita
,O.amamiensis
,O.exiliversabilis
,O.nasica
,andO.nasuta
(vs.
two metatarsal tubercles).
The congenersO.kweichowensis
andO.wuchuanensis
have a sympatric distribution withOdorrana liboensis
sp.nov.
The new species can be distinguished from these species by a series of morphological characters as follows.The new species differs fromO.kweichowensis
by having a larger body size (adult males with a length of 47.1–49.9 mmvs.
adultO.kweichowensis
males with a length of 36.2–43.3 mm) and having a lower ratio of IND/IOD at 0.89 in males and 0.81 in females (vs.
1.14 in males and 1.19 in females forO.kweichowensis
),lacks black bars on the lips (vs.
the presence of black bars),an absence of vocal sacs in males (vs.
the presence of vocal sacks),lacks large black spots on the dorsum (vs.
large black spots in the center of the dorsum),has thighs with four brown bands and tibias with three (vs.
thighs with five brown bands and tibias with six),lacks white pectoral spinules in mature males (vs.
the presence of white pectoral spinules),and lacks toes with entire webbing (vs.
full webbing).The new species differs fromO.wuchuanensis
by having a smaller body size (SVL 47.1–49.9 mm in adult males and 55.8–58.2 mm in adult femalesvs.
71.1–76.5 mm in adult males and 75.8–90.0 mm in adult females) and a lower ratio of IND/IOD 0.89 in males and 0.81 in females (vs.
1.63 in males and 1.14 in females inO.wuchuanensis
).The new species has a tibiotarsal articulation reaching to between the eye and the nostril when the leg is stretched forward (vs.
reaching the nostril),lacks black bars on the lips (vs.
the presence of black bars),lacks large black spots on the ventral surface (vs.
the presence of large black spots),has relative finger lengths of Ivs.
the presence of large white spines).Odorrana liboensis
sp.nov.
is phylogenetically closest toO.lipuensis
,and this new species could be distinguished from the latter by tibiotarsal articulation reaching to between the eye and the nostril when the leg is stretched forward (vs.
reaching the anterior corner of the eye);relative finger lengths Ivs.
in males 0.24–0.31,mean 0.27,and in females 0.23–0.31,mean 0.27);males with a higher ratio of TED/HDL (0.11–0.13,mean 0.12vs.
0.08–0.10,mean 0.09);the absence of small white spiny grains from the anterior corner of the eye along the eye and via the tympanic membrane below to the anterior part of the cloacal foramen (vs.
the presence of grains);indistinct brownish-black bands on limbs (vs.
distinct);the throat,chest,and abdomen lacking black spots (vs.
the presence of black spots);and the ventral surface of the limbs is pink (vs.
light purple).Distribution and ecology.
Odorrana liboensis
sp.nov.
is known only from the type locality,Maolan National Nature Reserve,Libo County,Guizhou Province,China,at elevations between 645 and 728 m.The new species has only been found in one cave in the area located far from the village.There is no light in the cave,and the new species was found in a small pool approximately 1.5 m wide and 79 cm deep,about 145 m from the entrance of the cave,where the water temperature is approximately 20 °C all year round.The tadpoles were collected on July 23,2016,but no pairs of male and female adults were found to hold them,and with four years of survey data,the adults were collected only from mid-July to mid-August.Therefore,we speculate that the breeding period begins in late June and continues until about mid-August.Inside this cave,Odorrana liboensis
sp.nov.
is sympatric withChinapotamon maolanense
,Sinocyclocheilus longibarbatus
,Hipposideros armiger,Aselliscus stoliczkanus
,Murina liboensis
,andLeopoldamys edwardsi
.Outside the cave,no adults and tadpoles of the new species were found during a herpetological survey in the vicinity.However,a number of amphibians and reptiles can be found outside the cave,includingTylototriton asperrimus
,Quasipaa boulengeri
,Bufo gargarizans
,Kurixalus eif fingeri
,Rhacophorus dennysi
,Goniurosaurus liboensis
,Sinomicrurus
macclellandi
,Lycodon flavozonatum
,andOrthriophis moellendorf fi.
4.Discussion
Most species ofOdorrana
live in montane streams.Previously,only two species (O.wuchuanensis
andO.lipuensis
) had been reported to live in cave environments(Fei
et al
.,2012;Moet al
.,2015).We discovered,and report on,a third species ofOdorrana
,Odorrana liboensis
sp.nov.
,that inhabits cave environments.Two of these species (includingO.wuchuanensis
) are found in karst caves in Guizhou.Phylogenetic analyses based on three mitochondrial genes suggested thatOdorrana liboensis
sp.nov.
belongs toOdorrana
but is distinct from its congeners.The genetic distances of 12S rRNA and 16S rRNA between theOdorrana liboensis
sp.nov.
and the closely relatedO.lipuensis
were 6.06% and 5.19%.The genetic distances of 16S rRNA was greater than 5%,and this is greater than the distance that typically represents differentiation,at the species level,in frogs (>3%)(Venceset al
.,2005;Fouquetet al
.,2007).The new species was different from its congeners on the basis of many morphological characters,and this supports its validity.Odorrana liboensis
sp.nov.
described here increases the number ofOdorrana
species to 62,with 39 recorded from China (Frost,2021;AmphibiaChina,2021).Based on 49 previously named species,one new species,and three mitochondrial genes,we conducted a phylogenetic study of the genusOdorrana.
The number of species covered here and the amount of data analyzed exceeds previously reported data(Chenet al
.,2013;Liet al
.,2018a).Eight highly supported major clades were identified inOdorrana
(Clades A–H).The topology of this tree differed significantly from previous studies,focusing mainly on the root evolutionary branches ofOdorrana
(Figure 2).In the phylogenetic tree,O.lipuensis
+Odorrana liboensis
sp.nov.
was the first species to diverge from the genusOdorrana
,whereas in other studies by Chenet al
.(2013) and Liet al
.(2018a),the first species to diverge wasO.chapaensis
.In our phylogenetic tree,O.chapaensis
+O.geminata
(Clade D1) serves as a sister taxonomic unit to Clade D2 consisting ofO.andersonii
as well asO.wuchuanensis
,O.mutschmanni
,O.yizhangensis
,O.lungshengensis
,andO.anlungensis
.In contrast,in previous studies,O.chapaensis
appeared as a sister taxon in a clade that included all otherOdorrana
(Chenet al
.,2013;Liet al
.,2018a),or as a sister taxon toOdorrana
other thanO.lipuensis
(He,2017).Ye and Fei(2001) suggested that the primitive taxa ofOdorrana
may have originated from the Hengduan Mountains and the plateau of western Yunnan.The Guizhou plateau may have been the center of differentiation ofOdorrana
(Ye and Fei,2001).O.lipuensis
andOdorrana liboensis
sp.nov.
appear to have diverged fromOdorrana
and formed the ancestral evolutionary branch of the genusOdorrana
.Therefore,we believe that the ancestral distribution ofOdorrana
may be the region south of Guizhou and northwest of Guangxi.However,this preliminary speculation needs to be supported by data from additional species and nuclear genes.Biodiversity conservation in southwestern China is a priority of the Chinese government (Ministry of Environmental Protection,2015).Biodiversity conservation programs in this region play an important role in maintaining the stability of mountain ecosystems as well as protecting biodiversity.In the past three years alone,16 new amphibian species have been described from Guizhou Province,China (Zhanget al
.,2017;Liet al
.,2018a,b;Liet al
.,2019a,b;Liet al
.,2020a,b;Lyuet al
.,2019;Wanget al
.,2019;Luoet al
.,2020;Liuet al
.,2020;Lyuet al
.,2020;Suet al
.,2020;Weiet al
.,2020;Wanget al
.,2020;Chenget al
.,2021).The discovery of these new species suggests that amphibian species diversity in this region is severely underestimated.In the context of global warming,there is an urgent need for a comprehensive,systematic,and in-depth survey of the impacts of climate change on terrestrial vertebrates to provide a basis for scientific decisions regarding amphibian conservation (IPCC,2014).Acknowledgements This study was supported by programs of the Biodiversity Survey and Assessment Project of the Ministry of Ecology and Environment,China(2019HJ2096001006),the Strategic Priority Research Program B of the Chinese Academy of Sciences (CAS) (No.XDB31000000),the National Natural Science Foundation of China (NSFC 31460091),the National Animal Collection Resource Center,China (No.2005DKA21402),the Application of Amphibian Natural Antioxidant Peptides as Cosmetic Raw Material Antioxidants (No.QKZYD [2020]4002).We thank Gang LI,Xiang ZENG,Jun ZHOU,Hongtao CUI,Tao LIU,and Jian CHEN for their help during specimen collection.We thank Mr.Yunming MO,Ms.Xiaowen LIAO,and Prof.Weicai CHEN for their help in reviewing the specimens.We thank LetPub (www.letpub.com) for its linguistic assistance during manuscript preparation.
Appendix
Specimens examined
Odorrana lipuensis
(n
=8):China:Guangxi:Lipu County (type locality):four males:NHMG 1306001,NHMG 1306002,NHMG 20140702,and NHMG 20 140703;four females:NHMG 1303019,NHMG 1303018,NHMG 1306003,and NHMG 20140701.Odorrana kweichowensis
(n
=35):China:Guizhou Province:Jinsha County (type locality).18 males:GZNU 20170717008–010,GZNU 20170717014,GZNU 20170717016–019,GZNU 20170717021–023,GZNU 20170717025,GZNU 20170717026,GZNU 20170717028–7030,GZNU 20170718003,and GZNU 2017 071800;17 females:GZNU 20170717001–007,GZNU 20170718001–002,GZNU 20170725001–007,and GZNU 20170725013.Odorrana
wuchuanensis
(n
=17):China:Guizhou Province:Libo County (topotype locality):12 males:GZNU 20160729001–006,GZNU 20160729008,GZNU 20160806001–005;five females:GZNU 20160729007,GZNU 20160809001,GZNU 20160809003,GZNU 20160809006,and GZNU 20160809011.About the front cover:Bronchocela cristatella
(Kuhl,1820) is a moderately slender arboreal lizard existing throughout Southeast Asia (Malaysia,Myanmar,Thailand,Singapore,India,Philippines,Indonesia,Papua New Guinea,Andaman and Nicobar Islands of India).Eggshell structure and composition of this lizard were reported by Kun GUOet al.
in this issue.This lizard was drawn by Timan YANG and Hanwen KANG.猜你喜欢
杂志排行

Asian Herpetological Research的其它文章
- Chemical Composition and Crystal Structure of the Eggshell of the Green Crested Lizard Bronchocela cristatella (Agamidae)
- Mitogenomic Conservation Genetics of the Critically Endangered Liaoning Clawed Salamander
- A New Species of Glyphoglossus Günther,1869 (Anura:Microhylidae) from Western Yunnan,China
- DNA Barcoding and Systematic Review of Minervaryan Frogs (Dicroglossidae:Minervarya) of Peninsular India:Resolution of a Taxonomic Conundrum with Description of a New Species