南渡江下游海南长臀鮠生物学参数估算
2021-12-24梁智策孙安俯蔡杏伟郭志强
刘 飞,梁智策,潘 帅,孙安俯,蔡杏伟,郭志强
南渡江下游海南长臀鮠生物学参数估算
刘 飞1,梁智策1,潘 帅1,孙安俯1,蔡杏伟2,郭志强1
(1. 海南大学海洋学院//南海海洋资源利用国家重点实验室,海口 570228;2. 海南省海洋与渔业科学院,海口 571126)
【】开展海南长臀鮠()的生物学及种群资源评估,为南渡江下游的海南长臀鮠的种群保护和资源恢复提供基础依据。本研究从2019年10月至2020年9月在南渡江下游进行逐月采样,共采集677尾海南长臀鮠样品。利用FiSAT Ⅱ软件中的ELEFAN I 法对南渡江下游海南长臀鮠的种群参数进行估算,利用Beverton-Holt动态模型对南渡江下游海南长臀鮠的资源动态进行评估。南渡江下游海南长臀鮠体长,体质量关系为:= 0.000 0073.16(2= 0.97;< 0.05);海南长臀鮠von Bertalanffy 生长方程各参数为渐近体长∞= 299.25 mm、生长参数= 0.41,起点年龄0= -0.09 龄;根据生长方程推算出生长拐点年龄为3龄,拐点体长约为226.38 mm,拐点体质量为164.45 g。利用Pauly经验公式计算出自然死亡系数()为0.51,体长变换渔获曲线法分别计算出总死亡系数()为1.23,捕捞死亡系数()为0.72,捕捞开发率()为0.58;动态综合模型估算南渡江下游海南长臀鮠的开捕年龄为1.47 龄,对应开捕体长为149.5 mm。海南长臀鮠的渔业资源处于过度开发状态。
海南长臀鮠;生物学参数;过度捕捞;南渡江
海南长臀鮠()隶属于鲇形目(Siluriformes),长臀鮠科(Cranoglanididae),长臀鮠属(),为海南岛特有淡水鱼类,主要分布于海南岛昌化江及南渡江水系[1]。近10 a以来,海南岛淡水水域生态环境受到的影响不断加剧,淡水水生生物的栖息环境受到破坏[2,3]。此外,尼罗罗非鱼()、云斑尖塘鳢()等外来物种的入侵,也对海南长臀鮠生存造成了巨大威胁[2]。目前,根据《中国脊椎动物红色名录》,海南长臀鮠的濒危等级为濒危(EN)[4]。有关海南长臀鮠的研究大多集中在分类及遗传方面[5-7],有关海南长臀鮠生物学及资源动态的研究未见报道。鱼类的生长与死亡参数不仅是鱼类生物学研究的基础,同时还是研究鱼类种群结构变动的重要依据[8]。鱼类的硬组织(鳞片、耳石等)被广泛应用于鱼类年龄与生长的研究[9],但热带地区的年轮存在不清晰的缺点,年龄鉴定较为困难[10]。而体长频率分析法在估算鱼类资源量方面,具有操作方便及无需年龄数据而被广泛应用于生长与死亡参数的估算[11-13]。此外,鱼类准确资源量评估是开展渔业资源保护的重要依据[14]。Beverton和Holt建立的单位补充量渔获量的模型(B-H模型)被广泛用来分析过度捕捞对渔业资源造成的许多影响[15,16]。
海南岛地处热带,气候高温多雨,丰富水资源和复杂的生态环境使得海南岛具有丰富内陆鱼类资源[1,17-18]。本研究以南渡江下游的海南长臀鮠为研究对象,通过计算该物种体长组成,体长-体质量关系、以及利用体长频率方法估算海南长臀鮠的生长及死亡参数,利用B-H模型对其种群资源进行评估,为海南长臀鮠的种群资源保护提供理论依据。
1 材料与方法
1.1 研究区域与样品采集
本研究样品采集于南渡江下游(图 1),采样时间为2019年10月至2020年9月,每月采样时间定为10号至15号。每月利用3条多网目复合刺网(长:30 m; 高:1.5 m;网目(2/cm)分别为1.0、6.0和12.5)和地笼(长:4 m;网目:2= 0.4 cm)。将样品收集起来,测量南渡江下游海南长臀鮠的体长(精确到1 mm)和体质量(精确到0.01 g)等生物学参数。每月捕捞海南长臀鮠数量见表1。
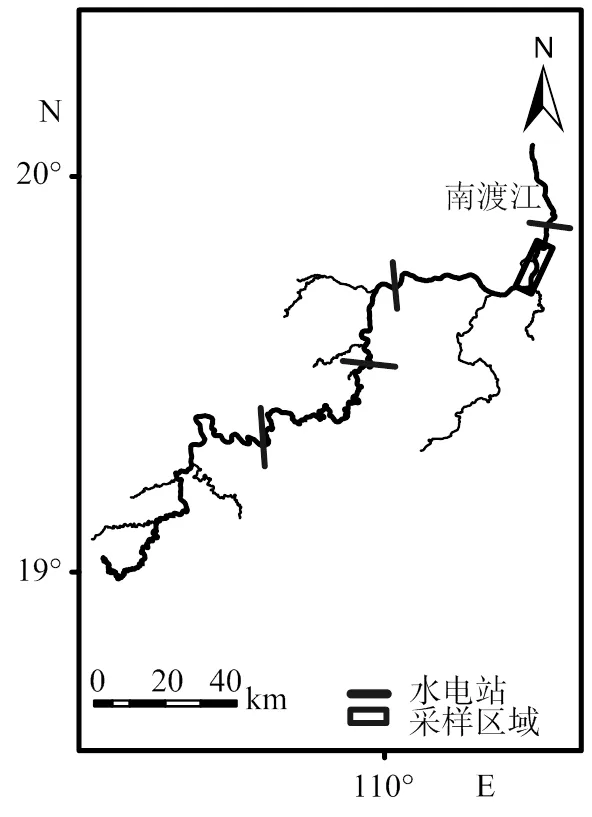
图1 南渡江下游海南长臀鮠采样位点
1.2 研究方法
1.2.1 体长与体质量关系 利用=aL对样品体长与体质量关系进行拟合,关系式中为样品体质量(g),为肥满度,为关系式系数,为鱼体体长(mm)。
1.2.2 生物学参数的计算 对样品体长以10 mm为组距进行分组[19],共50 ~ 59 mm,60 ~ 69 mm,270 ~ 279 mm,280 ~ 289 mm 等25组,并计算每组频率。输入海南长臀鮠体长频率数据,利用FiSAT Ⅱ软件中的 ELEFAN Ⅰ版块估算海南长臀鮠生长系数()和渐近体长(∞)[20]。然后根据Pauly经验公式,输入和∞计算其理论生长起点年龄0[21]:
ln (-0) = -0.3922 – 0.2752 ln∞- 1.038 ln,(1)
海南长臀鮠总死亡系数()利用体长变换渔获曲线法计算,自然死亡系数()利用自然死亡系数估算模块估算[19,22]。
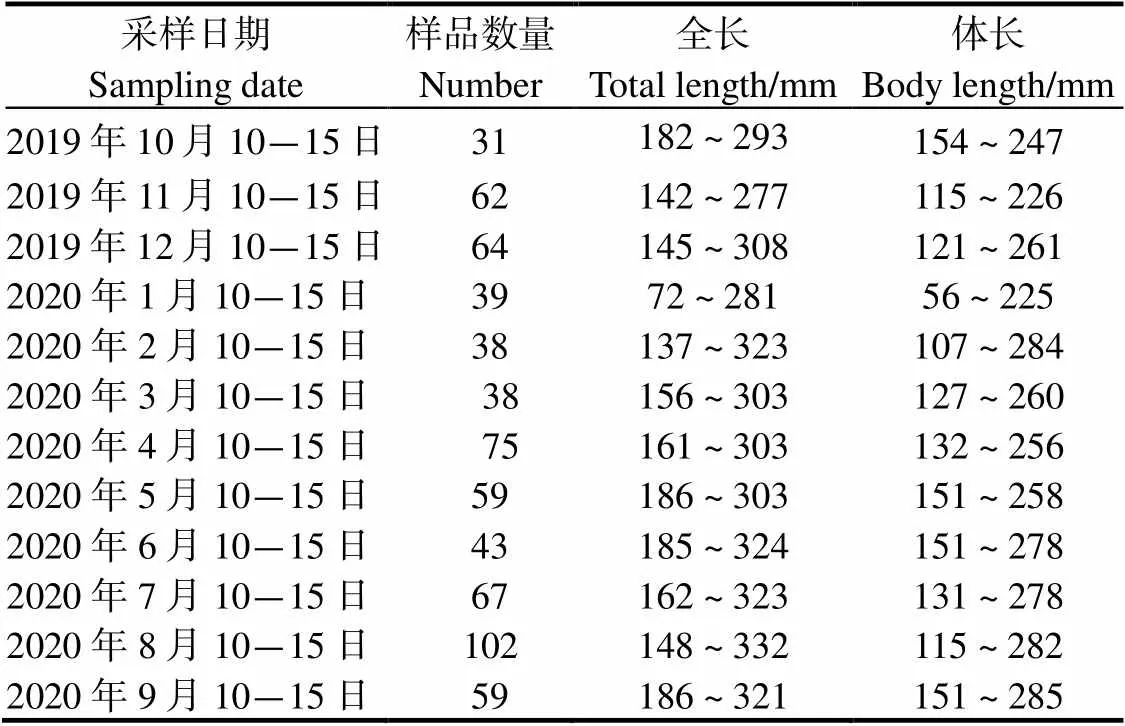
表1 南渡江下游海南长臀鮠样本数量和规格
1.2.3 生长特性指标 基于von Bertalanffy 生长方程[23],输入0(理论生长起点年龄)、L(体长)、m(体质量)等参数,拟合海南长臀鮠的体长、体质量生长方程:
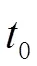
对海南长臀鮠的体长,体质量生长方程的进行一阶和二阶求导,计算其体长、体质量的生长速度和加速度曲线方程。
1.2.4 鱼类单位补充量渔获量
1.2.4.1 估算平均选择体长 利用体长变换曲线对总死亡系数()进行计算:ln (/△) =+t,其中为第个体长组的数量(尾),△为其他体长组与第个体长组的年龄间隔,为常数,t为第个体长组的相对年龄,-即为总死亡系数()[24]。然后计算各点观察与期望的比值及其累计率,当累计率达50.0%时所对应的体长即为海南长臀鮠的开捕体长()[25]。
1.2.4.2 相对单位补充量渔获量分析 Beverton和Holt提出动态综合模型(dynamic pooled model)以计算鱼类种群参数变化和渔获量之间的关系[15]。Pauly对模型进行了改进以分析相对/(单位补充量渔获量)和相对单位补充量生物量/(相对单位补充量生物量)[26]。在FiSAT Ⅱ软件中,开发率一般有三种:1)max,是最大渔获量时的开发率[27];2)0.5,为资源量下降到原始水平50.0%时的开发率[27];3)0.1,为/边际增长减少10.0%时的开发率[27]。
2 结果
2.1 体长分布
本研究在南渡江下游共采集海南长臀鮠677 尾,体长分布为56 ~ 285 mm,平均体长为 194.73 mm,南渡江下游海南长臀鮠的优势体长为200 ~ 290 mm,占全部体长组的84.47%(图 2)。
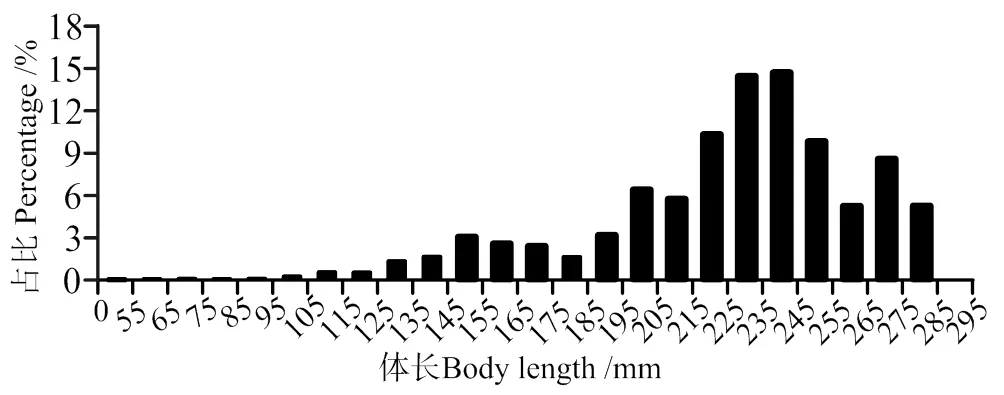
图2 海南长臀鮠体长分布
2.2 体长与体质量关系
通过对南渡江下游海南长臀鮠的体长和体质量进行拟合,海南长臀鮠体长和体质量拟合的关系式为:
= 0.000 0073.16(2= 0.97;< 0.05;= 677),(4)
其体长体质量关系的幂指数为3.16,与3具有显著性差异(< 0.05),因此海南长臀鮠为正异速生长(图 3)。
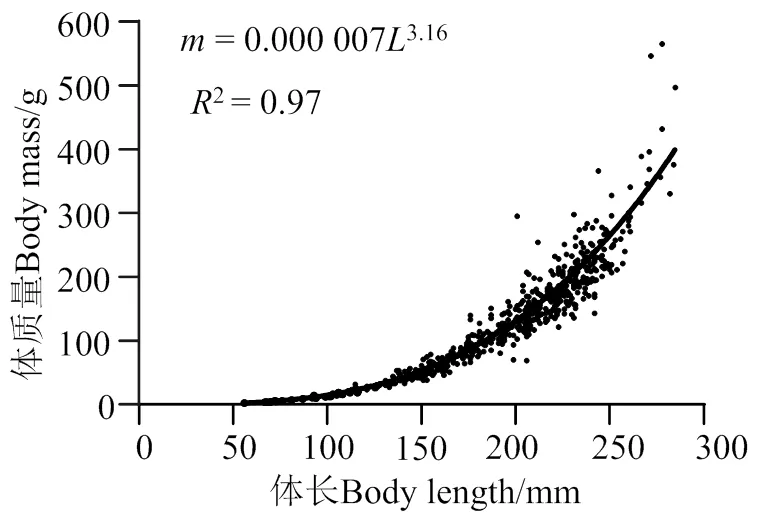
图3 海南长臀鮠体长与体质量关系式
2.3 生长方程及生长参数
利用FISAT Ⅱ软件中的ELEFAN Ⅰ方法中扫描功能计算海南长臀鮠的渐近体长(∞)为299.25 mm,生长参数为0.41。然后根据Pauly提出的用于计算理论生长起点年龄的经验公式计算出0为-0.09,因此海南长臀鮠的龄时的体长与体质量理论生长方程分别为:
L= 299.25 [1 - e-0.41 (t + 0.09)], (5)
m= 467.05 [1 - e-0.41 (t + 0.09)]3.16, (6)
海南长臀鮠年龄与体长、体质量拟合曲线见图4。
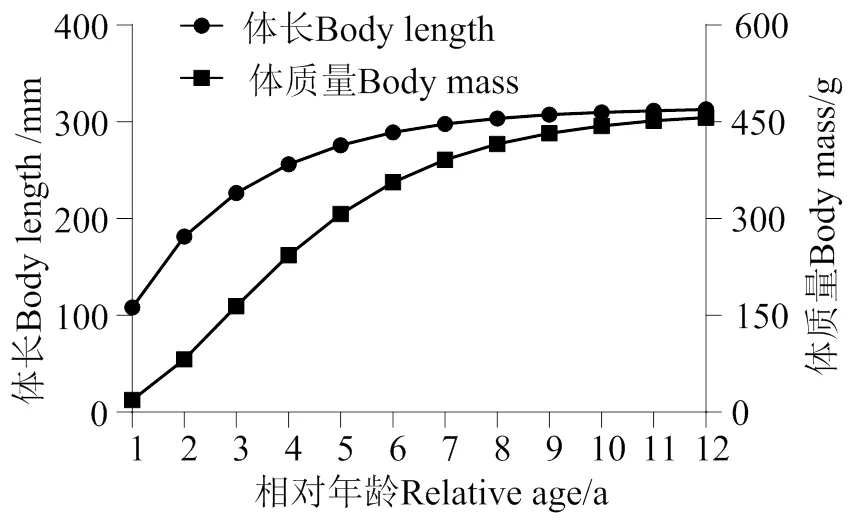
图4 海南长臀鮠von Bertalantly生长曲线
利用体长变换渔获物曲线法计算总死亡系数(),并对其进行拟合(图5):
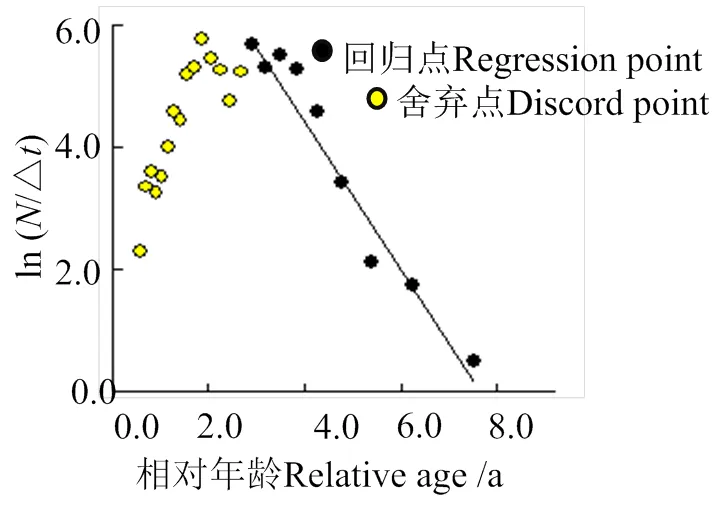
图5 体长变换渔获曲线估算南渡江下游海南长臀鮠总死亡系数(Z)
ln() = -1.23+ 9.482 (2= 0.96), (7)
式中,为相对年龄,为数量。
拟合曲线的系数为-1.23。因此,海南长臀鮠的总死亡系数()为1.23,总死亡系数()的95%置信区间为0.99 ~ 1.47。
将南渡江下游海南长臀鮠的渐近体长∞(299.25 mm)和生长参数(0.41)及采样点年平均水温 26.8 ℃代入Pauly[22]经验公式计算出0为-0.09,。利用体长变换曲线计算自然死亡系数为0.51、总死亡系数为1.23、捕捞死亡系数得为0.72,开发率为为0.58。
2.4 生长特性与拐点年龄
通过对南渡江下游海南长臀鮠的生长方程进行求导得海南长臀鮠的生长速度及生长加速度,由图 6、7可见海南长臀鮠的生长变化过程。
体长生长速度dL/d= 122.69 e-0.41 (t +0.09)。
体长生长加速度d2L/d2= -50.30 e-0.41 (t +0.09)。
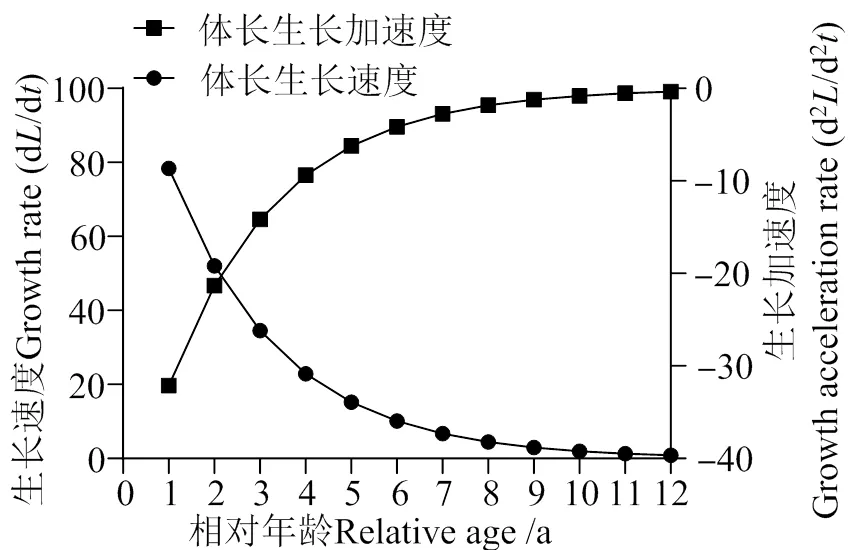
图6 海南长臀鮠体长生长速度与加速度变化曲线
体质量生长速度dm/d= 574.47 e-0.41 (t + 0.09)×[1- e-0.41(t + 0.09)]2。
体质量生长加速度d2m/d2= 235.53 e-0.41 (t +0.09)×[1- e-0.41 (t + 0.09)] × [3e-0.41 (t + 0.09)- 1]。
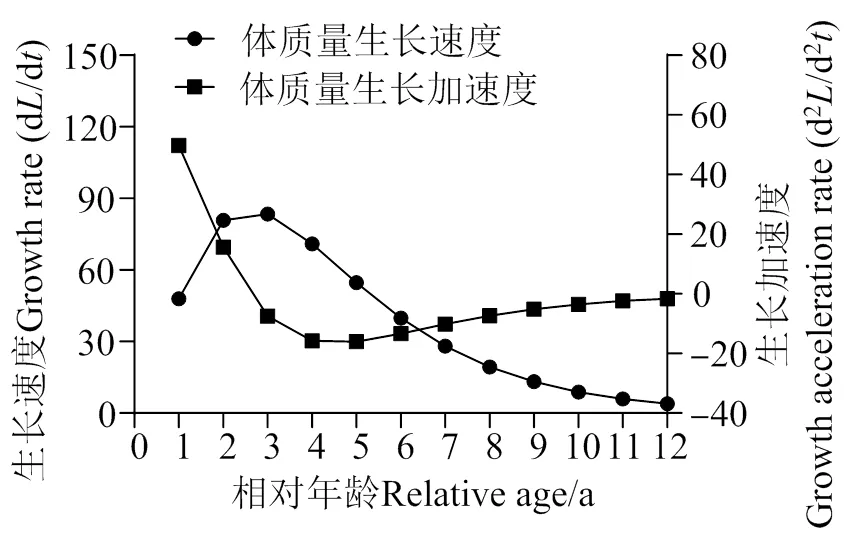
图7 海南长臀鮠体质量生长速度与加速度变化曲线
南渡江下游海南长臀鮠体长的增长随年龄的增长而减小,在高龄时期逐渐平稳趋于0(图 6)。海南长臀鮠的体质量的生长速度呈现出先上升后下降的趋势。其中在3龄之前,海南长臀鮠的体质量的生长速度随年龄的增长而增长,3龄后随年龄增长而下降,因此推测南渡江下游的海南长臀鮠的体质量的最大生长速度出现在2 ~ 3龄之间。通过计算得海南长臀鮠的生长拐点年龄为3 龄(体长226.38 mm,体质量164.45 g)。
2.5 单位补充量渔获量
2.5.1 估算平均选择体长利用FISAT Ⅱ软件对开捕体长进行估算,当累计率达到50%时对应的体长为(c)为149.5 mm,开捕体长(c)对应的年龄为1.47。
2.5.2 计算相对单位补充量渔获量 通过FISAT Ⅱ软件中的B-H模型计算,南渡江下游海南长臀鮠单位补充量渔获量(/)与开发率()的关系如图所示(图8)。其中,= 1.24,c/L= 0.50,估算出max= 0.689,0.1= 0.604,0.5= 0.363。
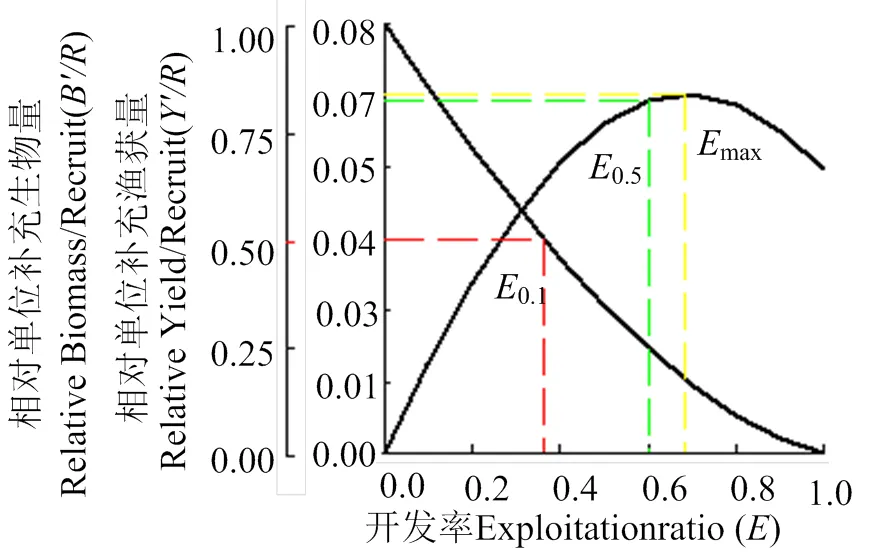
图8 海南长臀鮠相对单位补充量渔获量、生物量与开发率的关系
通过分析南渡江下游海南长臀鮠的相对单位补充量等值曲线,可得/与开捕体长c和开发率之间的动态变化趋势(图 9)。
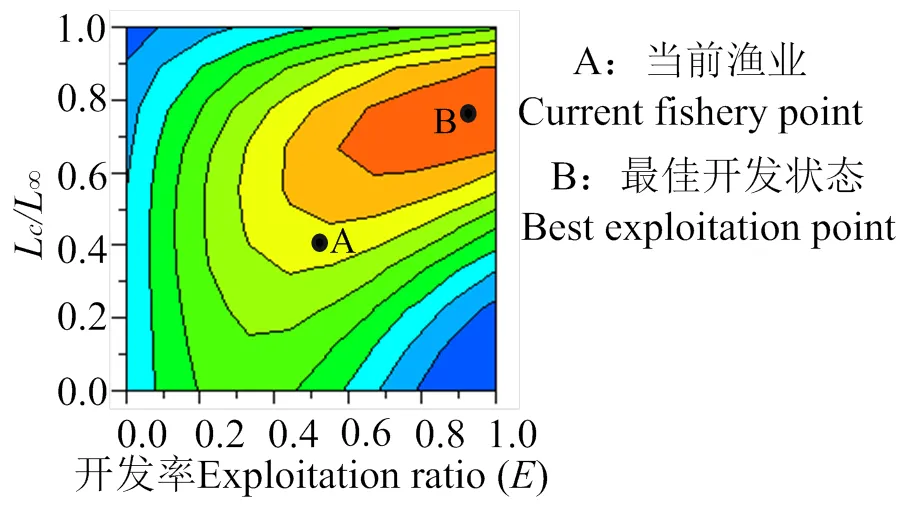
图9 海南长臀鮠相对单位补充量与开发率和开捕体长的关系
如图9所示,当前南渡江下游海南长臀鮠渔业点为A点,且开发率()为 0.58,c/L为 0.50[即开捕年龄(c)为 1.47,对应的开捕体长(c)为 149.5 mm]。南渡江下游海南长臀鮠理论上的最佳开发状态为B点,在/为 1.24 不变的情况下,单位补充量渔获量/达到最大值 0.077,其对应的开发率约为1.00,c/L约为 0.80。若从A点调到B 点,单位补充量渔获量/将达到最大值,其对应的开发率增大72%及c/L增大60%。目前海南长臀鮠开捕体长为149.5 mm,而生长拐点对应的体长为226.38 mm,若增加开捕体长,/呈现先增大后减小的趋势。当c/L为0.7,即开捕体长为209.45 mm时,/达到最大值0.06。
3 讨论
3.1 参数估算与资源量评估
有关鱼类年龄的鉴定的常用材料包括鳞片、耳石等硬组织材料[28]。海南长臀鮠体表无鳞片,因此无法利用鳞片进行年龄的鉴定。而利用耳石进行年龄的鉴定存在耗时长的问题,且在进行年龄的鉴定过程中需要较高技能,不同的鉴定者对于同一个耳石材料得出的年龄的鉴定结果一致性的概率小于90%[29]。而长度频率法是利用鱼类体长频率对生长参数进行估算,避免利用硬组织鉴定年龄过程中的主观性误差,因此在渔业资源的评估中得到了广泛应用[29-30]。但是由于对于不同研究对象和采样数据误差会影响模型本身的适用性。因此在利用体长频率法(LCA)进行评估时需要大量的样品以及足够的月份,且尽可能获得大个体样本[31]。而本研究利用地笼以及多网目复合刺网进行逐月采样,所采集样本体长频率分布合理,且大个体样本较多(图2),保证了体长频率法估算海南长臀鮠的准确性。
鱼类生长参数和自然死亡系数是渔业资源评估中的两个重要参数[32]。自然死亡系数受到环境水温、鱼类生长状况等诸多因素的影响[33]。鱼类生长参数与鱼类的渐近体长L具有负相关关系,与鱼类自然死亡系数具有正相关关系[26]。当e-< 1时,表明von Bertalanffy生长方程能够反映该鱼类的生长特性[33-34]。而本研究中= 0.41,e-= 0.66 < 1,表明von Bertalanffy可以用来拟合海南长臀鮠的生长。根据Gulland[35]提出资源最佳开发率为0.5,本研究中南渡江下游海南长臀鮠的资源开发率为0.58,表明海南长臀鮠处于过度捕捞状态。根据Pauly[21]等提出最佳开发率水平(= 0.4),即= 0.16,同样表明南渡江下游海南长臀鮠处于过度开发状态。
3.2 南渡江下游海南长臀鮠资源利用与保护
鱼类种群大小,开捕年龄及捕捞强度等因素会影响渔业资源的状态,进而影响渔业资源的可利用性[36]。其中,捕捞死亡系数与初次开捕体长是渔业管理中两个重要的人为控制方式,通过对这两个因素的控制可以实现最大可持续产量[19]。本研究通过B-H动态综合模型对南渡江下游海南长臀鮠渔业状态进行分析,结果表明南渡江下游的海南长臀鮠的野生资源利用具有不合理性(图 9),主要为南渡江下游海南长臀鮠的捕捞强度偏大,而开捕年龄则较小。由图8 可看出保持当前开发率= 0.58不变,增大开捕体长至209.45 mm,相对单位补充量渔获量可达到0.06,/增长1%,与最佳状态下的0.077基本持平;而降低南渡江下游的海南长臀鮠捕捞强度,则会导致/降低。通过对增大开捕体长与增大捕捞强度两种方式比较,可以得出增大开捕体长比减小捕捞强度更能达到保护资源的同时又合理利用资源的目的。
[1] 中国水产科学研究院珠江水产研究所. 海南岛淡水及河口鱼类志[M].广州:广东科技出版社, 1986: 1-406.
[2] 申志新, 李高俊, 蔡杏伟, 等. 海南省淡水野生鱼类多样性演变及保护建议[J]. 中国水产, 2018(11): 56-60.
[3] 蔡杏伟, 申志新. 海南长臀鮠[J]. 生物资源, 2020, 42(3): 366.
[4] 蒋志刚, 江建平, 王跃招, 等. 中国脊椎动物红色名录[J]. 生物多样性, 2016, 24(5): 501-551.
[5] 杨家坚. 海南长臀鮠人工繁殖与胚胎发育的研究[J]. 上海水产大学学报, 2008, 17(6): 684-688.
[6] 谢少林, 吕子君, 周爱国, 等. 基于线粒体Cytb和COⅠ基因对珠江、海南和红河长臀鮠群体遗传结构分析[J]. 生态科学, 2016, 35(3): 65-72.
[7] 乐小亮, 赵爽, 刘海林, 等. 基于线粒体细胞色素b基因全序列的海南长臀鮠南渡江种群遗传变异分析[J]. 生态科学, 2010, 29(3): 247-250.
[8] GILPIN J A, CHADWICK N E. Life-history traits and population structure of pederson cleaner shrimps[J]. The Biological Bulletin, 2017, 233(3): 190-205.
[9] ASSEFA W W, WONDIE A, ENYEW B G. Population dynamics and exploitation patterns ofin Lake Tana, northwest Ethiopia[J]. Lakes & Reservoirs: Research & Management, 2019, 24(4): 344-353.
[10]LOWE-MCCONNELL R H. Ecological studies in tropical fish com-munities[M]. London: Cambridge University Press, 1987.
[11] 高春霞, 麻秋云, 田思泉, 等. 浙江南部近海小黄鱼生长、死亡和单位补充量渔获量[J]. 中国水产科学, 2019, 26(5): 925-937.
[12] 郭峻宏, 李军, 沈忱, 等. 闽江口龙头鱼()的生物学参数及资源评估[J]. 广东海洋大学学报, 2019, 39(5): 56-64.
[13] 叶金清, 柳圭泽, 袁秀堂, 等. 基于生物学特征的辽河口鮻资源开发与保护[J]. 大连海洋大学学报, 2020, 35(2): 302-307.
[14] 吴金明, 娄必云, 赵海涛, 等. 赤水河鱼类资源量的初步估算[J]. 水生态学杂志, 2011, 32(3): 99-103.
[15] BEVERTON R J H, HOLT S J. On the dynamics of exploited fish populations[M]. Dordrecht: Springer Netherlands, 1993.
[16] RICKER W E. Handbook of computations for biological statistics of fish populations [J]. Can Fish Res Board Bull, 1958: 119-300.
[17] 中国水产科学研究院珠江水产研究所, 华南师范大学, 暨南大学, 上海水产大学, 湛江水产学院. 广东淡水鱼类志[M]. 广东: 广东科技出版社, 1991: 1-622.
[18] 郑慈英. 珠江鱼类志[M]. 北京: 科学技术出版社, 1989: 1-452.
[19] 詹秉义. 渔业资源评估[M]. 北京: 中国农业出版社, 1995: 1-353.
[20] GAYANILO F C, SPARRE P, PAULY D. FAO-ICLARM Stock Assessment Tools Ⅱ (Fisat Ⅱ): User’s Guide (Revised Version) [M]. Rome: FAO Computerized Information Series (Fisheries), 2005: 1-152.
[21] PAULY D. Length-converted catch curves and the seasonal growth of fishes[J]. Fishbyte, 1990, 8(3): 24-29.
[22] PAULY D, Morgan G. Length-based Methods in Fisheries Research [M]. World Fish1987: 5-89.
[23] VON BERTALANFFY L. A quantitative theory of organic growth (Inquires on growth laws.Ⅱ) [J]. Human Biology, 1938, 10: 181-213.
[24] SPARRE P, WILLMANN R. BEAM 4, a bio-economic multi-species, multi-fleet, multi-plant, multi-area extension of the traditional forecast model[M]. Copenhagen: ICES, Statistical Committee. 1992.
[25] 何宝全, 李辉权. 珠江河口棘头梅童鱼的资源评估[J]. 水产学报, 1988, 12(2): 125-134.
[26] PAULY D, SORIANO M L. Some practical extensions to Beverton and Holt’s relative yield-per-recruit model[J]. First Asian Fisheries Forum, 1986(5): 26-31.
[27] 田辉伍, 岳兴建, 陈大庆, 等. 怒江东方墨头鱼资源量和死亡参数的估算[J]. 生态学杂志, 2012, 31(1): 235-240.
[28] 张学健, 程家骅. 鱼类年龄鉴定研究概况[J]. 海洋渔业, 2009, 31(1): 92-99.
[29] 高春霞, 田思泉, 戴小杰. 淀山湖刀鲚的生物学参数估算及其相对单位补充量渔获量[J]. 应用生态学报, 2014, 25(5): 1506-1512.
[30] CUNHA N L, CATELLA A C, KINAS M A. Growth parameters estimates for a small fish of the Pantanal, Brazil:(Characiformes; Characidae)[J]. Brazilian Journal of Biology, 2007, 67(2): 293-297.
[31] HOENING, J. M., CSIRKE, J. SAN DE RS, M. J, et al. Data acquisition for length-based stock assessment: report of Working Group I [C]. Italy, International Centre for Living Aquatic Resources Management, 1987:343-352.
[32] 朱国平, 宋旗. 运用脂褐素鉴定甲壳类年龄的研究进展[J]. 生态学杂志, 2016, 35(8): 2225-2233.
[33] STERGIOU K I, VOULGARIDOU P. Trends in various biological parameters of the European sardine,(Walbaum, 1792), in the Eastern Mediterranean Sea[J]. Scientia Marina, 2003, 67(S1): 269-280.
[34] 费鸿年, 张诗全. 水产资源学[M]. 北京: 中国科学技术出版社, 1990: 1-787.
[35] GULLAND J A. fish stock assessment: a manual of basic methods [M]. Chichester: Wiley Interscience, 1983.56-76.
[36] 王继隆, 刘伟, 鲁万桥, 等. 黑龙江中游乌苏里白鲑资源现状评估[J]. 生态学杂志, 2019, 38(6): 1824-1829.
Estimation of Biological Parameters ofin the Downstream of Nandu River
LIU fei1, LIANG Zhi-ce1, PAN Shuai1, SUN An-fu1, CAI Xing-wei2, GUO Zhi-qiang1
(1.//,570228,; 2.,571126,)
【】To perform the biology and population resource assessment ofand to provided basic information for the population protection and resource recovery ofin the downstream of Nandu River. 【】A total of 677 fish samples were collected from the downstream of Nandu River from October 2019 to September 2020 monthly. The population parameters ofwere calculated by the ELEFAN I part of FiSAT Ⅱ software, and the population dynamic ofwas estimated by Beverton-Holt dynamic model.【】The length()-mass() relationship ofin the downstream of Nandu River was=0.000 0073.16(2= 0.97;< 0.05); Von Bertalanffy growth equation parameters ofwere asymptotic length∞= 299.25 mm, curvature parameter= 0.41,0(theoretical age when the theoretical length of fish was 0) =-0.09years. Calculated by growth equation, the inflection age was 3 years, inflection body length was226.38 mm, inflection mass was164.45 g for.The natural mortality calculated by Pauly’s empirical function was 0.51, the total mortality() coefficient, fishing mortality () and exploitation rate() estimated by length-converted catch curve method were 1.23,0.72, and 0.58,respectively. The capture age and body length ofcalculated by dynamic pool model were 1.47 years and 149.5 mm, respectively.【】in the downstream of Nandu River is overexploited.
; biological parameters; overfishing; Nandu River
S932.4
A
1673-9159(2021)06-0131-07
10.3969/j.issn.1673-9159.2021.06.016
刘飞,梁智策,潘帅,等. 南渡江下游海南长臀鮠生物学参数估算[J]. 广东海洋大学学报,2021,41(6):131-137.
2021-06-30
2020年海南省重点研发计划(ZDYF2020193);国家自然基金(31872574, 41866003, 31960731)
刘飞(1999-),男,硕士研究生,主要研究方向为渔业资源保护。E-mail: liufei@hainanu.edu.cn
郭志强,E-mail: guozq@hainanu.edu.cn
(责任编辑:刘岭)
