不同海拔短肋羽藓叶绿素含量与光合效率研究
2021-12-14唐录艳陈泽宇庄李婕朱毛洁
唐录艳 ,陈泽宇,戴 尊,姚 雪,庄李婕,朱毛洁,彭 涛*,王 健
(1.贵州师范大学生命科学学院,贵州 贵阳 550025;2.杭州第二中学钱江学校,浙江 杭州 311215;3.华东师范大学生命科学学院,上海 200241)
植物的生长与分布受光照、水分及温度等环境因子直接或间接影响。与纬度梯度相比,海拔梯度因在短距离内包含了温度、湿度和光照等各种环境因子的剧烈变化,而成为研究物种分布格局及成因的重要区域[1],其环境因子变化比纬度梯度快 1000倍[2]。研究表明,海拔梯度上复杂的环境因子是限制植物分布的重要因素,随着海拔升高,气温、大气压及 CO2分压下降,UV-B辐射及光强增加,这些都将对植物的形态与生理产生重要影响[3—5]。在海拔梯度上,由于受不同环境因子的影响,同一物种长期生活在不同的小生境中,其生理生态特性也会发生变化,以形成适合自身生存的生理机制[3]。在海拔梯度上,一般海拔越高意味着植物生存的环境越恶劣,在此环境中生存的植物常形成抵御外界胁迫环境的特殊生理机制,主要表现在植物的光合特性、体内化学物质组成等方面[6]。
光合作用是植物叶片吸收温室气体的主要途径,也是植物生长和代谢活动的生理基础[7],同时也是最易受环境影响的重要生理过程,因此不同生态环境中的植物具有不同的光合特性[8]。苔藓植物部分种类在弱光下的叶绿素含量比强光下更高,能增强其在弱光环境中光捕获能力,从而具有更高的光合效率[9—11]。苔藓植物这种光合喜阴性被认为是对周围环境长期适应和演化的结果[12—13]。研究不同海拔苔藓植物的光合色素及光合作用效率,对于探讨和理解苔藓植物对海拔梯度的适应、分布及应对未来气候变化都具有重要意义。然而目前国内有关海拔梯度上植物的生理特性以及对气候的适应性研究大多集中于种子植物[14],对不同海拔苔藓植物光合作用效率的响应规律的研究及报道相对缺乏。本研究以高黎贡山百花岭的短肋羽藓(Thuidium kanedae)为材料,比较分析海拔梯度上的植株体叶绿素含量和实际光合量子产量变化,旨在揭示不同海拔苔藓植物适应生态环境的生理机制。
1 材料与方法
1.1 研究区概况
百花岭隶属云南省保山市芒宽乡境内,紧靠高黎贡山自然保护区南段东坡面,其中心点地理坐标为 98°42′40″E,25°17′17″N,海拔 760~3100 m[15—16]。境内山势陡峻,具有深大的断裂纵谷,地势高差大,垂直立体气候明显,从河谷到山顶依次出现热带、南亚热带、中亚热带、北亚热带、暖温带、中温带和寒温带的气候带[17]。森林植被茂盛,海拔1000~1800 m之间分布着季风常绿阔叶林,1800~2800 m之间分布着中山湿性常绿阔叶林,2700~3100 m之间分布着寒温性灌丛、云南铁杉(Tsuga dumosa)林和山顶苔藓矮林[18]。
1.2 方法
1.2.1 材料采集
以自然生长于百花岭海拔 1500~3100 m的短肋羽藓为实验材料,在2019年7月植物生长旺季进行采集。参照该地区不同林型,将采样段分为低海拔(1500~1800 m)、中海拔(1900~2700 m)和高海拔(2800~3100 m)三个区段。从海拔1500 m处开始,每隔100 m为一个海拔梯度,共17个梯度,每个梯度分别采集3份适量的新鲜短肋羽藓标本,每份标本取3份样品进行实验,测定生理指标。
1.2.2 叶绿素含量测定
参考张志良等[19]的方法,每个海拔梯度的标本称取0.5 g样品于研钵,加5 mL丙酮以及适量石英砂充分研磨后,转至离心管中以2000 r·min-1转速离心3 min,吸取上清液定容至8 mL。采用分光光度法分别于 663 nm和 645 nm处测得光密度值(OD值)。使用普析通用公司 T6新世纪光密度计,计算提取液的叶绿素a、b含量和叶绿素a/b比值及叶绿素总含量。
1.2.3 光合效率测定
参考张伟等[12]的方法使用 Junior-PAM 调制荧光仪测定各海拔梯度短肋羽藓新鲜叶片实际光化学量子产量 Y(Ⅱ),以反映其实际光合效率及开放的PS Ⅱ反应中心原初光能捕获效率[20]。
1.3 数据分析
采用 SPSS 26软件对不同海拔短肋羽藓叶绿素含量和实际光合量子产量Y(Ⅱ)做单因素方差分析;采用R语言Corrplot包进行相关性分析;采用局部加权回归(Loess regression)对测定指标的海拔分布规律进行拟合分析,比较叶绿素含量、叶绿素 a/b比值和实际光合效率随海拔升高的变化趋势。
2 结果与分析
2.1 不同海拔短肋羽藓叶绿素含量和实际光合量子产量
考虑在100 m海拔梯度间短肋羽藓叶绿素含量和实际光合量子产量的差异不显著,故以200 m为一梯度进行统计。由表1可知,在百花岭海拔1500~3100 m内,不同海拔段短肋羽藓的总叶绿素含量范围在 0.227~0.450 mg·g-1,在海拔 1900~2000 m 中所采集的短肋羽藓总叶绿素含量较高;各海拔梯度的短肋羽藓叶绿素a含量整体高于叶绿素b含量,其中叶绿素a含量在海拔1700~2000 m内较高,而叶绿素b含量则在海拔1900~2000 m内较高;叶绿素a/b比值范围为2.200~7.083,在海拔1700~1800 m内比值最大,而其他海拔梯度内的叶绿素 a/b比值差异不显著;实际光合量子产量 Y(Ⅱ)代表了实际光合效率,其范围为0.229~0.276,在海拔2900~3100 m内较大,表明其实际光合效率较高。

表1 不同海拔短肋羽藓的叶绿素含量和实际光合量子产量Table 1 Chlorophyll content and actual photosynthetic quantum yield of Thuidium kanedae at different altitude
2.2 不同海拔短肋羽藓叶绿素含量和实际光合量子产量的相关性
图1表明短肋羽藓叶绿素含量与实际光合量子产量Y(Ⅱ)之间的关系,从中可看出总叶绿素含量与叶绿素a、叶绿素b及叶绿素a/b比值呈正相关关系,相关系数分别为0.980、0.578和0.512。叶绿素a/b比值与叶绿素a和叶绿素b分别呈正、负相关关系,相关性系数分别为0.678和-0.391。实际光合量子产量Y(Ⅱ)与叶绿素总含量、叶绿素a、叶绿素b含量及叶绿素 a/b比值之间都呈负相关关系,相关系数分别为 -0.383、-0.342、-0.346和 -0.073。

图1 短肋羽藓叶绿素含量与实际光合量子产量的相关性Fig. 1 Correlation between chlorophyll content and actual photosynthetic quantum yield of Thuidium kanedae
2.3 不同海拔短肋羽藓叶绿素含量和实际光合量子产量的变化特征
由图2可看出,叶绿素a含量和叶绿素总含量的变化趋势相似,在海拔 1500~2500 m范围内呈“单峰”分布,然后随着海拔的继续升高变化不明显。叶绿素b含量在海拔1500~2500 m范围内也呈现“单峰”分布模式,但之后随着海拔升高又呈明显上升趋势。叶绿素a/b比值在1500~2100 m海拔范围内呈现“单峰”分布模式,随后随着海拔继续升高而缓慢下降。实际光合量子产量 Y(Ⅱ)在海拔1500~2700 m范围内呈缓慢上升趋势,随后随着海拔继续升高而出现一个急剧上升趋势(图3)。
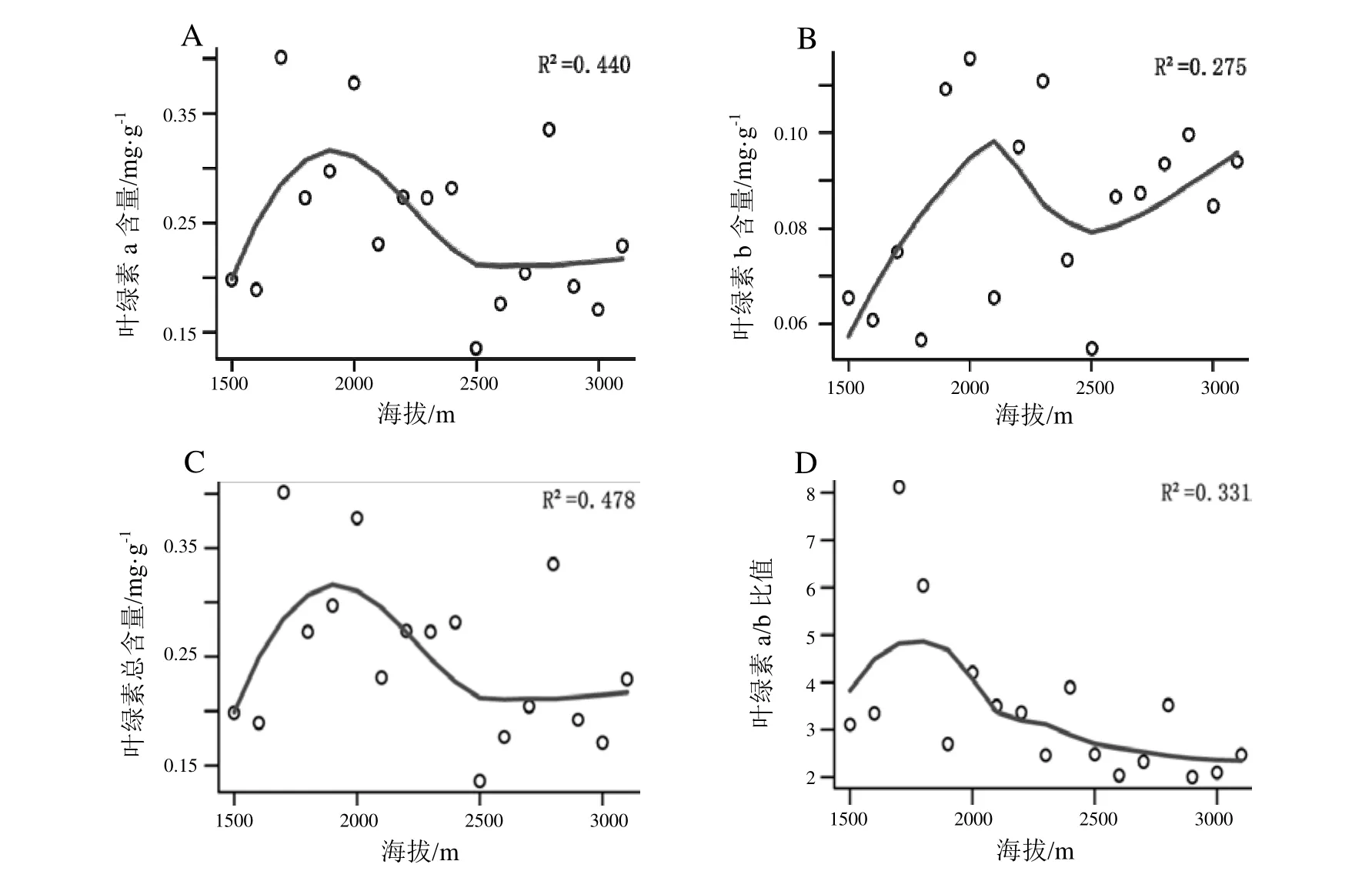
图2 不同海拔短肋羽藓叶绿素含量的变化特征Fig. 2 Variation characteristics of chlorophyll content of Thuidium kanedae at different altitude

图3 不同海拔短肋羽藓实际光合量子产量的变化特征Fig. 3 Variation characteristics of actual photosynthetic quantum yield of Thuidium kanedae at different altitude
3 讨论
光合作用是植物物质积累及生理代谢的基本过程和储存太阳能的唯一途径,同时也是对逆境反映最为敏感的生理过程之一[21]。光合色素反映了植物利用光能的能力,可用于判断植物光合生理能力的强弱,反映环境胁迫的状况等[10]。一般而言,苔藓植物较高的叶绿素含量强化其在弱光下的光捕获能力,从而使其具有更高的光合效率[22]。但张伟等[12]研究表明,苔藓植物叶绿素含量与其光合作用效率之间并非简单的线性相关关系,更多反映出苔藓植物配子体表面的水分与光照之间的平衡[23]。本研究发现,短肋羽藓实际光合量子产量Y(Ⅱ)与叶绿素a、叶绿素b含量及叶绿素总含量、叶绿素a/b比值之间都呈负相关关系,进一步支持张伟等[12]研究的结论。
在种子植物中,关于叶绿素含量与海拔之间的关系有两种观点。一种观点表明,随着海拔的升高,植物叶片内的叶绿素a、叶绿素b及叶绿素(a+b)含量都降低[3,24—26],另一种观点是植物叶片叶绿素含量随海拔升高呈先上升后下降的趋势[3,27]。本研究与后者相似,即短肋羽藓的叶绿素总含量、叶绿素 a含量和叶绿素b含量均表现出在海拔1500~2500 m范围内随着海拔升高呈先升高而后下降的“单峰”分布模式。但随着海拔的进一步升高,叶绿素 a+b含量和叶绿素a含量变化不明显,而叶绿素b含量表现出明显的上升趋势。
叶绿素a/b比值是衡量植物耐阴性的重要指标。研究表明,叶绿素 a/b比值低有利于提高植物对远红光的吸收,有利于植物在低光度条件下吸收较多的光照以提高光合效率,是植物在弱光条件下形成的一种生理适应[28]。在维管植物中,可能由于高海拔地区的低温和强辐射导致叶绿素b比叶绿素a降解得更快的缘故,叶绿素 a/b比值一般随着海拔的升高而增加[26]。相对于维管植物,苔藓植物具有较低的光饱和点、光补偿点及叶绿素 a/b比值,表现出典型的阴生植物特性[22,29—30]。本研究发现短肋羽藓植物体内叶绿素a/b比值在海拔1500~2100 m范围内呈现“单峰”分布模式,随后表现出随海拔继续升高而缓慢下降的趋势。与维管植物不同,短肋羽藓叶绿素 a/b比值整体表现出随海拔升高而下降的趋势,而实际光合量子产量Y(Ⅱ)整体表现出随海拔升高而增加的趋势,并在高海拔处(2700~3100 m)出现急剧上升。故可看出高黎贡山百花岭地区短肋羽藓对海拔梯度有着相应的光合生理适应机制。
全球气候变暖带来的影响已不容忽视,并将会在未来的很长一段时间持续影响我们。随着全球气候的变暖,物种为了获得合适的生存环境,将向更高纬度或更高海拔迁移[31]。相对于纬度梯度,海拔梯度因为在较小的地理范围内具有巨大的环境变化而成为开展生物多样性分布规律及成因研究的“天然实验室”[1,32]。在众多解释物种多样性海拔分布假说中,与环境相关的水分-能量动态假说被讨论得最多[33],该假说主要基于水分对生物体能量利用过程的影响,直接影响植物光合作用和呼吸作用[34]。与其他高等植物相比,苔藓植物作为一类以配子体世代为主要世代的变水植物,对外界环境的水分和能量变化更为敏感[23]。在我国亚热带山地,温度一般随海拔升高而逐渐降低,而水分的有效峰值一般出现在降雨量和土壤水分保持量高且蒸发量低的中海拔地区[35],这主要是因为低海拔地区的高温导致蒸发量超过降水量而变得干燥,高海拔地区的较浅土壤层和较大径流量都将降低水分的有效性。本研究中短肋羽藓实际光合量子产量 Y(Ⅱ)随海拔的变化趋势,表明低温相对于水分的有效性对其生长更具促进作用,这与前人研究结论相一致,即苔藓植物的生长和分布普遍表现出对低温的适应[36—37]。而气候变暖主要对物种的海拔分布下缘(即低海拔)产生重要影响[38—39],这将对分布于低海拔处的短肋羽藓产生严重的威胁。建议针对苔藓植物更多种类开展类似研究,以了解其在海拔上的分布及生长情况,为其应对未来的气候变化研究提供科学依据。
