茶树品种“紫娟”及其诱变后代遗传多样性分析
2021-12-14陈春林田易萍邓少春徐丕忠仝佳音尚卫琼
陈春林,田易萍,邓少春,徐丕忠,仝佳音,尚卫琼
(1.云南省农业科学院茶叶研究所,云南 勐海 666201;2.云南省茶树种质资源创新与配套栽培技术工程研究中心,云南 勐海 666201;3.云南省茶学重点实验室,云南 勐海 666201)
【研究意义】诱变是一种常规的农作育种方法,包括化学诱变和物理诱变。诱变具有应用方便、诱变后代稳定、特异性较强等优点,已广泛应用于农作物育种领域,目前对农作物诱变后代的多世代筛选、鉴定技术日益成熟,已经培育出具有生产利用价值的农作物新品种,在农业生产中产生了巨大的经济效益和社会效益[1-4]。“紫娟”属云南大叶种变种,因其叶、茎、花萼、花梗、茶汤均呈紫色而得名。“紫娟”的花青素含量较高,达到29.14 mg/kg,花青素含量远高于其他茶树品种且茶氨酸和儿茶素总量差异也很大[5-9]。大量研究表明,紫娟茶具有降压、保护肝脏、提高记忆力、影响脂类代谢及抗氧化等功效。因此,“紫娟”诱变后代的花青素含量具有较高的分析价值[10-11]。【前人研究进展】李谨等[12]以仙客来干种子为试材,利用“神州八号”飞船进行太空诱变处理,结果显示太空诱变对仙客来植株在花色和花形上与对照相比表现出明显的表观性状差异,所有的突变株DNA均发生了变化;姜振峰等[13]用0.04%的化学诱变剂叠氮化钠处理大豆,结果显示诱变后代有矮化或晚熟现象,植株性状特征发生了明显的变化。近年来,随着分子生物学的快速发展,微卫星标记在作物遗传多样性研究上被广泛应用,谢淑芹等[14]通过NaN3诱变小麦后再利用SSR分析,结果表明利用NaN3处理种子出苗率高,后代性状变异类型丰富,变异率高;孔佑涵等[15]人探索NaN3处理柑橘腋芽的诱变效应中采用SRAP分子标记检测发现一些新梢变异叶片的基因组DNA发生了变化。【本研究切入点】基于陈林波等[16-17]前期对“紫娟”的芽、第二叶、开面叶、成熟叶转录组高通量测序所得的242 757条Unigene,在确定的44对EST-SSR引物为数据的研究基础上开展研究,一是通过观测“紫娟”诱变后代的植物学性状,明确诱变后代的表型特征;二是分析诱变后代的主要生化组分差异,明确诱变后的生化成分变化;三是利用EST-SSR分析诱变后代的多态性。【拟解决的关键问题】通过上述研究,从形态特征、生化组分和分子水平上探讨太空和NaN3诱变对“紫娟”的诱变效果,为将来利用太空和NaN3诱变茶树育种提供理论基础。
1 材料与方法
1.1 材料
于2020年6月11日在云南省农业科学院茶叶研究所试验基地采集“紫娟”及后代材料的幼嫩叶片及新梢一芽二叶,按相对稳定的工艺技术制备蒸青样,并保存于-80 ℃的冰箱中。
1.2 方法
1.2.1 植物学性状观测 参照茶树种质资源描述规范和数据标准,观测“紫娟”及其诱变后代植物学性状,重点比较叶色、叶片大小、芽叶色泽、一芽三叶百芽重等。
1.2.2 内含成分测定 花青素测定方法采用分光光度计法,按照FB/LH 003—2010标准进行分析;水浸出物测定方法采用分光光度计法,按照GB/T8305—2013标准进行分析;茶多酚测定方法采用分光光度计法,按照GB/T 8313—2018标准进行分析;游离氨基酸测定方法采用分光光度计法,按照GB/T 8314—2013标准进行分析;咖啡碱测定方法采用分光光度计法,按照GB/T 8312—2013标准进行分析。
1.2.3 引物的设计合成 合成引物基本信息见表1。

表1 合成引物基本信息
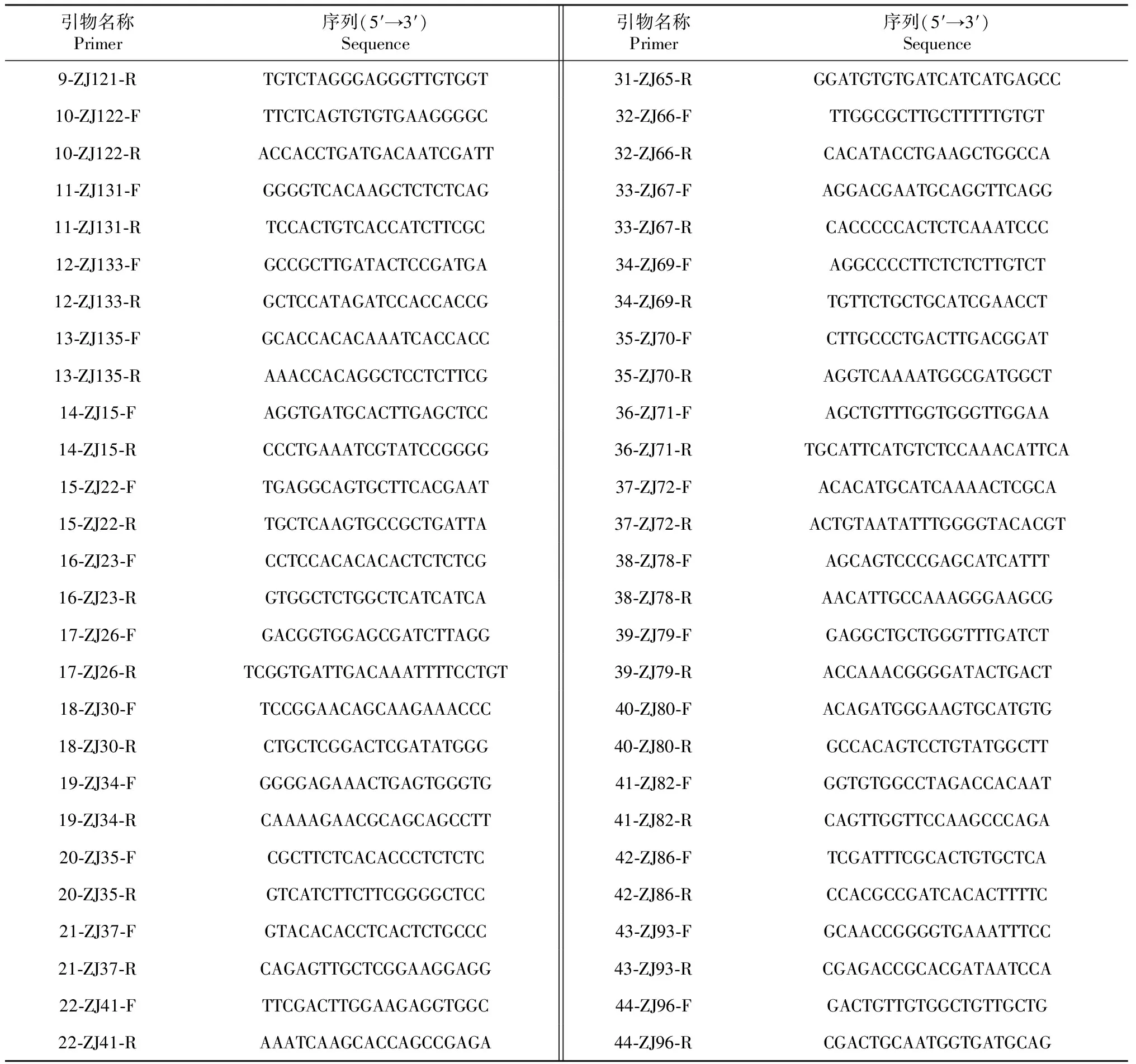
续表1 Continued table 1
1.2.4 DNA提取 取100 mg幼嫩叶片加入液氮充分研磨,利用CTAB的方法提取DNA,取2 μL DNA用于1.2%琼脂糖凝胶电泳检测,取2 μL DNA用于NanoDrop分光光度计测浓度。
1.2.5 PCR扩增 PCR反应体系10×Buffer I:1.5 μL;2.5 mmol/L dNTP 1.2 μL,5 μmol/L F/R 0.8 μL,DNA 1.0 μL,ddH2O 15 μL。PCR反应程序:95 ℃ 5 min,95 ℃30 s,60 ℃30 s,72 ℃30 s,72 ℃30 min,循环35次。
1.2.6 电泳检测 96孔板中每孔加入分子量内标和甲酰胺混合液(0.5∶8.5)9 μL,PCR产物1.0 μL;95 ℃变性3 min,上机检测。
1.3 数据分析
将检测得到的原始数据文件导入到分析软件genemapper ID 3.2中进行分析。
2 结果与分析
2.1 “紫娟”诱变后代的植物学性状
“紫娟”茶的主要特点为紫芽、紫叶、紫茎,从表2可知,有15份材料芽叶色泽与“紫娟”相近,且除ZJ-05、ZJ-06、ZJ-19以外,其余都花青甙显色,春季一芽三叶百芽重除ZJ-17,夏茶中除ZJ-01、ZJ-02、ZJ-03、ZJ-04外其余均超过“紫娟”;春季一芽二叶百芽重、一芽一叶百芽重均超过“紫娟”,夏茶中一芽二叶百芽重除了ZJ-02、ZJ-03、ZJ-04、ZJ-06和ZJ-17以外,其余均已超过“紫娟”,一芽一叶百芽重比“紫娟”低的有ZJ-02、ZJ-03、ZJ-09、ZJ-17、ZJ-19共5份材料;单芽百芽重中春季有2份材料与“紫娟”相当,分别为ZJ-17、ZJ-19,其余的比“紫娟”要高,夏茶有6份材料比“紫娟”低,分别为ZJ-02、ZJ-03、ZJ-07、ZJ-09、ZJ-16、ZJ-17。

表2 “紫娟”及其诱变后代的新梢性状
由表3可知,各材料成熟叶片的叶形只有ZJ-10和ZJ-11与“紫娟”相同,为披针形,其余的为椭圆形或长椭圆形,叶面积最大的为ZJ-01,达到37.01 cm2,ZJ-03和ZJ-04为小叶,其余的为中叶;叶色与“紫娟”相同的有9份材料,都为紫绿色。
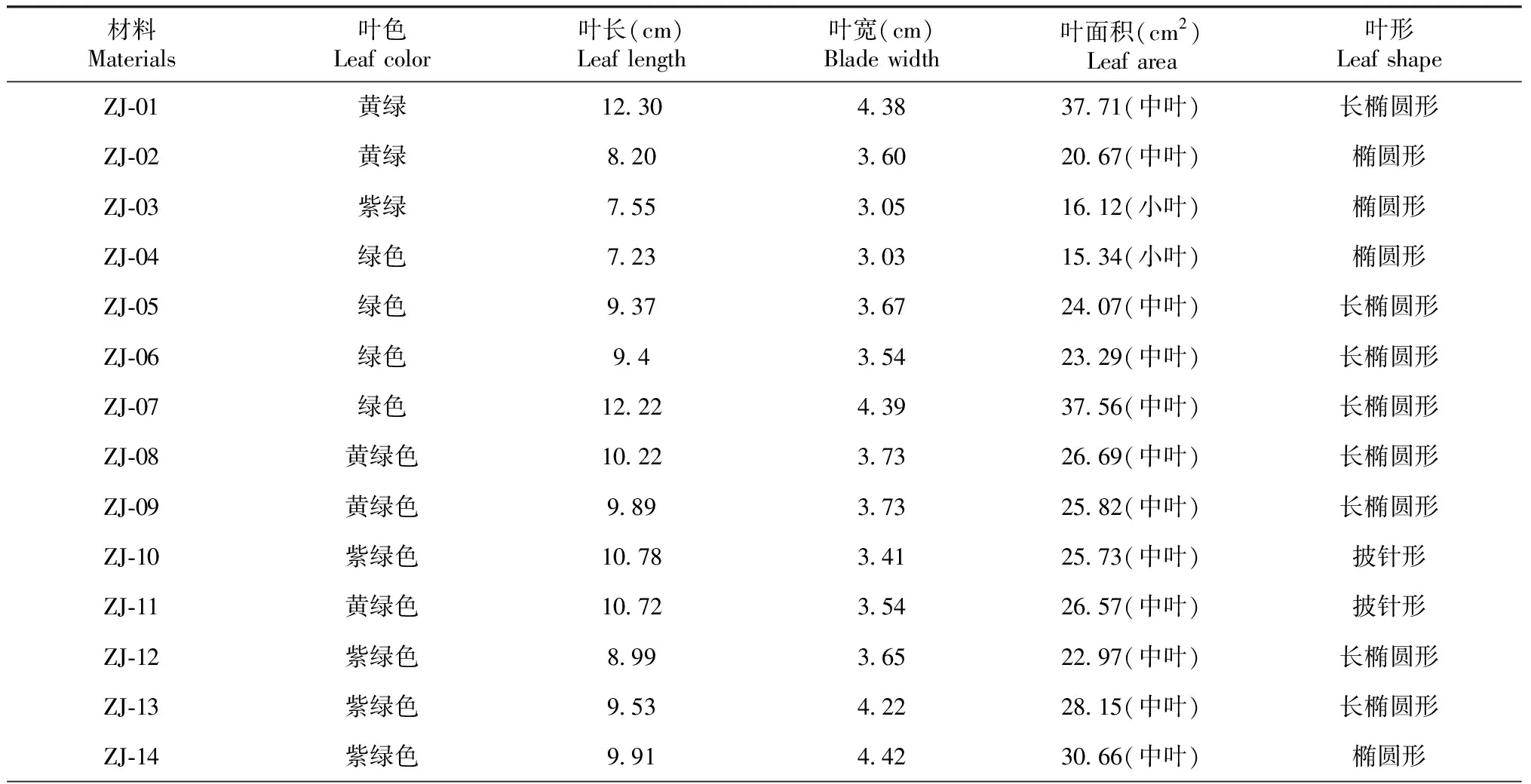
表3 “紫娟”及其诱变后代的叶片性状

续表3 Continued table 3
由表4可知,20份材料中花柱的长度、开裂数与“紫娟”基本一致,在裂位上,ZJ-02和ZJ-08为中裂,其余的为浅裂。
2.2 “紫娟”诱变后代的生化成分分析
由表5可知,春季花青素含量超过10%的有3份,其中ZJ-12花青素超过“紫娟”;春季水浸出物含量最低的为ZJ-01,最高的的为ZJ-15,超过“紫娟”的共有9份材料;茶多酚含量超过“紫娟”的有10份材料,最高的为ZJ-15、最低的为ZJ-11;游离氨基酸含量最高的为ZJ-11,最低的为ZJ-13,超过“紫娟”的有7份材料;咖啡碱含量超过“紫娟”的材料有6份材料。由表6可知,夏茶花青素含量超过10%的有5份材料,总体上夏茶的花青素含量要高于春茶;夏季水浸出物含量最低的为ZJ-18,最高的为ZJ-06,超过“紫娟”的共有9份材料;茶多酚含量超过“紫娟”的有7份材料,最高的为ZJ-15,最低的为ZJ-11;游离氨基酸含量最高的为ZJ-20,最低的为ZJ-05、ZJ-06及ZJ-16,超过“紫娟”的有4份材料;咖啡碱含量超过“紫娟”的材料只有ZJ-06。
春、夏两季进行比较,花青素总量的变异系数最大,春季为72.16%,夏季为77.86%,表明诱变后代的花青素具有较为丰富的多样性,遗传改良的潜力较为巨大。春季游离氨基酸总量的变异系数大于20%,但夏季的变异系数才7.6%,这可能与春季游离氨基酸含量整体偏高有关。

表4 “紫娟”及其诱变后代的花器官性状

表5 春季 “紫娟”及其诱变后代的生化成分

表6 夏季 “紫娟”及其后代的生化成分
2.3 “紫娟”及其后代的DNA质检
21个样品中提取的DNA电泳质检合格,结果见图1。
2.4 “紫娟”及其后代的SSR遗传多样性分析
通过毛细血管电泳,使用genemapper ID 3.2软件分析对44对SSR引物的扩增结果进行分析,分析结果见表7,44对引物共扩增到165个等位基因,每个位点扩增到2~10个等位基因,平均等位基因7.5个,各位点多态信息量(PIC)变化范围0.233~0.846;有效等位基因(Ne)在1.368~7.221,平均值为2.562,与观察到的等位基因数存在着较大差距,表明等位基因在群体内分布不均匀;观测杂合度(H0)变化范围在0.048~1.000,平均值为0.629,期望杂合度(He)变化范围在0.269~0.862,平均值为0.565,不同位点的杂合度差异较大,平均观测杂合度高于期望杂合度,说明群体内杂合度较高;Shannon信息指数最高的为2.083,43个微卫星标记有18个为高度多态性(多态信息量>0.5),25个为中度多态位点(0.25<多态信息含量<0.5),表明“紫娟”及其后代遗传多样性较为丰富。

M1500为1500 bp marker,ZJ01~ZJ20为“紫娟”的后代 M1500 is 1500 bp marker;ZJ01-ZJ20 are Zijuan’s mutagenesis progeny图1 DNA电泳图Fig.1 DNA electrophoretogram

表7 43个SSR位点的遗传多样性和变异性
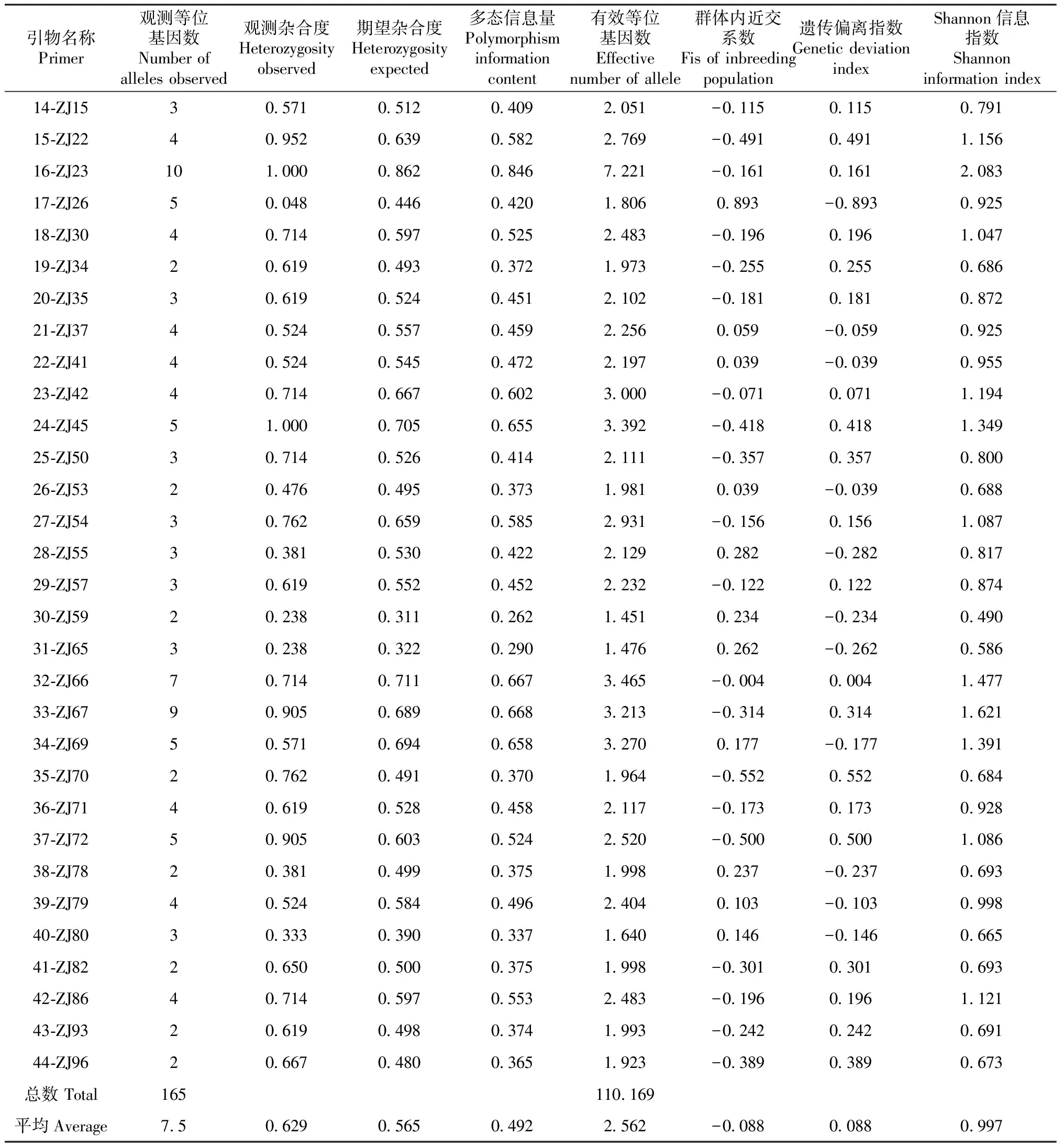
续表7 Continued table 7
3 讨 论
试验选定的20份“紫娟”诱变后代材料在叶色、芽色、叶形等外部形态特征上具有明显的差异性,通过春、夏两季花青素组分分析,证实花青素是“紫娟”叶片呈紫色的主要原因,验证了前期的推论,花青素含量多少影响着芽叶色泽,选定的诱变材料在生化成分上具有较高的遗传多样性。但是试验也有不足之处,茶树属于异花授粉作物,可能在自然杂交过程中会有自然突变的情况发生,这就造成诱变后代是因为化学或太空造成的突变还是自然环境造成的突变不得而知,故在今后的诱变后代分析上,首先可以采用扦插苗或穗条进行诱变,减少自然因素的干扰;再者分析群体时,可以把自然杂交群体和诱变群体进行一个遗传多样性分析,找出遗传多样性的差异,从而判断诱变效果。此外由于20份“紫娟”的诱变后代材料属于单株群体,长势各有不同,在进行春、夏理化成分分析时,蒸青样很难采够20 g的分析量,故最后春季只能分析15份,夏季只能分析14份,造成分析结果不够全面,数据不够完整,在今后的研究过程中,可以采用扩繁的方式,来弥补数据的缺失。
新梢叶色有15份材料芽叶色泽与“紫娟”相近;成熟叶片的叶形只有2份材料与“紫娟”相同,为披针形,其余的为椭圆形或长椭圆形,叶色与“紫娟”相同的有9份材料,都为紫绿色,有7份材料叶色为黄绿色,有4份材料为绿色;花器官中的花萼有8份材料与“紫娟”相同,花萼花青甙显色;20份材料中花柱的长度、开裂数与“紫娟”基本一致,在裂位上,ZJ-02和ZJ-08为中裂,其余的为浅裂。
春、夏季生化成分的变异系数中,变异系数最高的为花青素,春茶达到72.16%,夏茶达到77.86%,花青素含量变化夏季大于春季,这个趋势与谭晓琴[18]的研究较为一致;两季中的茶多酚、水浸出物、游离氨基酸、咖啡碱变异系数都在30%以下,最低的为夏季游离氨基酸变异系数,达到7.60%。春季游离氨基酸的平均值为3.23%、夏季游离氨基酸的平均值为2.37%,春季茶多酚平均值为20.09%,夏季为24.34%,这个变化趋势与周顺珍等人[19]研究基本一致。
多态信息含量(PIC)反应SSR位点的变异程度,数值越大,变异越多,“紫娟”诱变后代的43个微卫星标记有18个为高度多态性(多态信息量>0.5),25个中度多态位点(0.25<多态信息含量<0.5),平均值为0.492,已接近0.50的高多态性标准值,说明诱变后代具有一定的变异程度。
等位基因数与有效等位基因数相差,群体间的差异性越小,选育空间越小[20]。通过数据表明等位基因平均数为7.5,有效等位基因数为2.562,两指标差异较大,说明诱变后代的群体差异较大,遗传多样性非常丰富。本研究的多态信息量指数(PIC)平均为0.492,低于自然杂交的,但是自然杂交的Shannon信息指数I又远低于诱变后代的0.997。这与杨军等[16]的研究结果一致。从群体的角度来说,通过诱变的子代群体遗传多样性较为丰富。
4 结 论
诱变后代生化成分变异系数最大的是花青素,变异系数均高于70%,说明诱变后代在花青素这个单一指标上具有很高的遗传多样性。诱变后代的某些表型特征与花青素具有一定的相关性,当花青素含量高甚至超过紫娟时,表型特征上芽叶色泽为紫绿色或紫色、芽叶花青甙显色、叶片色泽也呈紫色或紫绿色。这些生化成分或表型特征呈现出特异性的诱变后代是遗传研究和品种创制的物质基础,有待进一步的跟踪鉴定。本研究通过44个微卫星标记对20份“紫娟”后代的遗传多样性进行分析,其中引物3-ZJ10-F扩增不出外其余的都成功扩增出清晰条带,通过数据分析得出“紫娟”诱变后代的平均多态信息量为0.492,不同位点的杂合度差异较大,平均观测杂合度高于期望杂合度,群体内杂合度较高;Shannon信息指数最高的为2.083,43个微卫星标记有18个为高度多态性,25个为中度多态位点,表明“紫娟”及诱变后代遗传多样性较为丰富。
