优质强筋小麦新品种郑品优9号的遗传基础解析
2021-12-08陈晓杰范家霖程仲杰杨保安张福彦焦学俭白鹤峰王嘉欢张建伟
陈晓杰,杨 科,范家霖,程仲杰,杨保安,张福彦,焦学俭,白鹤峰,王嘉欢,张建伟
(1.河南省科学院同位素研究所有限责任公司/河南省核农学重点实验室,河南郑州 450015;2.郑州市农业技术推广站,河南郑州 450006; 3.河南金苑种业股份有限公司,河南郑州 450001)
小麦是我国最主要的口粮作物之一,其产量和品质对保证粮食安全至关重要[1- 2]。优质是小麦育种的重要目标之一,然而我国小麦品质育种工作起步较晚,从“七五”开始,小麦品质育种才正式列入国家重点科技攻关项目,经过30多年的小麦品质研究和遗传改良,一批强筋小麦品种(如豫麦34、郑麦9023、藁城8901、新麦26、郑麦366、西农979、济麦44等)陆续被选育和推广,为我国优质强筋小麦生产和满足消费者需求做出了重大贡献。目前我国优质强筋小麦品种在高产稳产、抗倒春寒、抗病性、抗逆性等方面与高产小麦品种仍存在一定差距[3-4]。因此,从分子水平对优质强筋小麦新品种的遗传基础和重要性状功能基因进行解析,对于优质小麦品种的遗传改良具有重要的指导意义。
近年来,前人对不同时期小麦骨干亲本和主栽品种的遗传构成进行了较多研究,结果表明,骨干亲本和主栽品种中含有与产量、品质、适应性、抗病抗逆性相关的染色体区段,这些区段更容易被育种家选择,在后代衍生品种中均具有较大的遗传贡献率[5-10]。肖永贵等[5]利用921个DArT标记和83个SSR标记对周8425B及其衍生品种的遗传结构进行解析,发现周8425B对其衍生子1代、子2代和子3代的平均遗传贡献率分别为67.7%、63.6%和58.8%,且周8425B的4个抗条锈病基因在衍生品种中传递。亓佳佳等[6]利用SSR标记对小麦骨干亲本小偃6号及其衍生品种(系)进行遗传解析,发现小偃6号对其衍生子1代、子2代、子3代和子4代的平均遗传贡献率分别为50.32%、47.54%、46.35%和44.83%。邹少奎等[7]利用SSR标记发现,母本周麦13号对周麦23号的遗传贡献率为63.04%,并筛选到一个可用于鉴定周麦23号的特异引物。杨子博等[8]利用SSR标记发现淮麦33更多地继承了母本烟农19的遗传物质。李玉刚等[9]利用SSR和SNP两种分子标记,提示青农2号的基因组大部分遗传信息来自鲁麦14(SSR标记:54.11%;SNP 标记: 72.55%)。吴胜男等[10]利用小麦55K芯片,分析了陕农981和新麦18对陕农33的遗传贡献率,发现11个与农艺和品质性状有关的QTL中,有3个来源于新麦18,有8个来源于陕农981。
随着小麦参考基因组的完成,越来越多与小麦重要性状相关的基因被定位或克隆,为育种家了解各品种的重要性状基因组成提供了依据,也为将来分子设计育种提供了支持。功能标记是根据功能基因内部引起表型性状变异的多态性基序开发出来的一种新型分子标记,这类标记不需要进一步验证就可以在不同的遗传背景下确定目标等位基因的有无,是作物育种中最有价值的一类标记。功能标记的高通量检测技术的开发有助于提高分子标记辅助育种的效率。竞争性等位基因特异性PCR(kompetitive allele-specific PCR,KASP)技术具有高通量、稳定、准确、灵活等优势[11-12]。目前,KASP标记已在分子辅助育种、QTL定位、亲本品种鉴定及大规模样本筛选等研究上得到应用[13-16]。中国农科院作物科学研究所何中虎研究员团队开发的小麦50 K SNP育种芯片不仅标记数量多且分布较均匀,而且包括上百个与株高、籽粒质量、品质、春化、光周期、开花、抗病、抗逆等小麦性状相关基因等位变异的 KASP功能标记,在遗传改良和育种上具有更高的利用价值[12,17]。
郑品优9号是以半冬性多穗型、中早熟、优质强筋小麦品种郑麦366为母本[18],以春性、早熟、优质强筋小麦品种豫麦34为父本进行杂交[19],F0代种子经60Co-γ 射线(200GY)处理,在F2M2代中选育而成的优质强筋小麦新品种。该品种表现为半冬性、矮秆、早熟、高产稳产、优质强筋、品质稳定,具有较大推广潜力。因此,本研究利用小麦50K SNP育种芯片,对郑品优9号及其父母本进行分子检测,旨在明确该品种遗传基础和重要性状功能基因的组成,为其遗传改良和生产应用提供参考。
1 材料与方法
1.1 供试材料
供试材料为郑品优9号及其双亲郑麦366和豫麦34,均由河南省科学院同位素研究所核农学实验室小麦诱变育种团队提供。
1.2 基因组DNA提取
选取每个小麦品种的幼嫩叶片,采用CTAB法提取小麦基因组DNA,并用UV-9000型紫外分光光度计检测DNA样品浓度。
1.3 SNP芯片分析
委托博奥晶典生物技术有限公司利用小麦50K SNP育种芯片对郑品优9号及其双亲材料进行SNP分析及功能基因KASP标记分析。功能基因标记包括产量性状相关基因(矮秆、粒重、籽粒形态等)、品质性状相关基因(高低分子量亚基、硬度、面团色泽等)、适应性相关基因(光周期、春化、开花等)和抗性性状基因(抗旱抗逆、抗穗发芽、抗病等)。
1.4 遗传贡献率分析及基因型图谱的绘制
首先剔除杂合或缺失的SNP位点,保留亲本和子代中纯合的SNP位点。根据双亲间纯合差异SNP位点数计算双亲遗传物质对子代的遗传贡献率,某一亲本对后代的遗传贡献率为后代中同该亲本相同的特异位点数与双亲特异位点总数的百分比。
利用GGT 2.0软件[20],依据染色体位置信息的纯合SNP 标记绘制郑品优9号及亲本的SNP基因型图谱,根据亲本中SNP标记的差异赋予不同的颜色,郑品优9号与亲本标记的异同赋予相应的颜色。
2 结果分析
2.1 双亲对郑品优9号的遗传贡献率
通过小麦50K SNP芯片扫描分析,结果在郑品优9号及其亲本郑麦366和豫麦34中共检测到44 716个纯和的SNP位点,其中纯合差异SNP位点6 042个,表现为相同位点数远多于差异位点数。从表1可以看出,4 124个纯合差异SNP来源于亲本郑麦366,其余1 918个来源于豫麦34,郑麦366和豫麦34对郑品优9号的遗传贡献率分别为68.26%和31.74%。
从基因组水平来看,在A基因组上,郑麦366和豫麦34对郑品优9号的贡献率分别为 68.60%和31.40%,与整体遗传贡献率接近;在B 基因组上,郑麦366对郑品优9号的遗传贡献率达到90.08%,提供了该基因组绝大部分的遗传信息;在D基因组上,豫麦34对郑品优9号的遗传贡献率远大于郑麦366,达72.37%(表1)。
从染色体水平来看,郑品优9号来源于双亲的遗传位点在不同染色体间差异较大(表1和图1)。郑麦366在不同染色体上对郑品优9号的遗传贡献率范围为7.53%~99.43%,其中在2A、4A、5A、2B、3B、4B、5B、6B、7B和2D染色体上对郑品优9号的遗传贡献率均超过80%。豫麦34在不同染色体上对郑品优9号的遗传贡献率范围为0.57%~92.47%,其中在7A、1D、6D和7D染色体上对郑品优9号的遗传贡献率均超过80%。在6A、1D、3D和4D染色体上纯合差异SNP位点数较少,均不到100个,表明双亲在这些染色体上差异较小,多态性差。
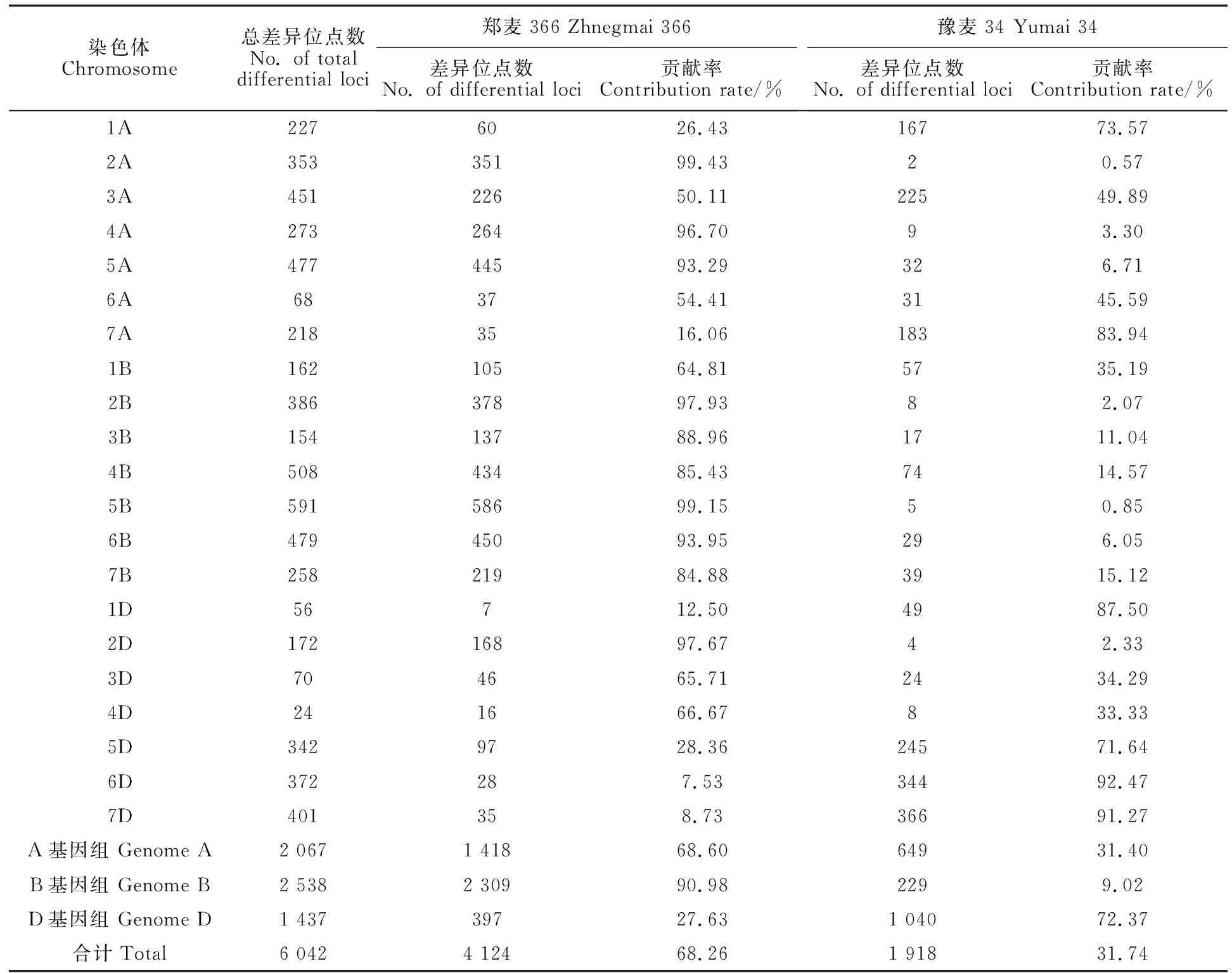
表1 郑麦366和豫麦34在不同染色体上对郑品优9号的遗传贡献Table 1 Genetic contribution of Zhengmai 366 and Yumai 34 to Zhengpinyou 9 on different chromosomes
2.2 郑品优9号来源于双亲的染色体区段
在检测到的44 716个纯和SNP位点中,有36 059个SNP位点有确切的染色体位置信息,其中30 379个SNP位点在郑品优9号和双亲间无差异,5 680个SNP在双亲中存在差异。依据这些SNP信息,利用GGT 2.0软件绘制了郑品优9号的SNP基因型图谱(图1)。在郑品优9号的21条染色体上,均检测到来自双亲的染色体区段,但比例不同。在2A、4A、5A、2B、3B、4B、5B、6B、7B、2D、3D和4D染色体上,郑品优9号检测到的差异区段主要来源于郑麦366;而在1A、7A、1D、5D、6D和7D染色体上,郑品优9号检测到的差异区段主要来源于豫麦34;在1B、3A、6A染色体上郑品优9号检测到的差异区段来源于双亲的比例接近。SNP基因型图谱分析结果与遗传贡献率分析结果具有较好的一致性。
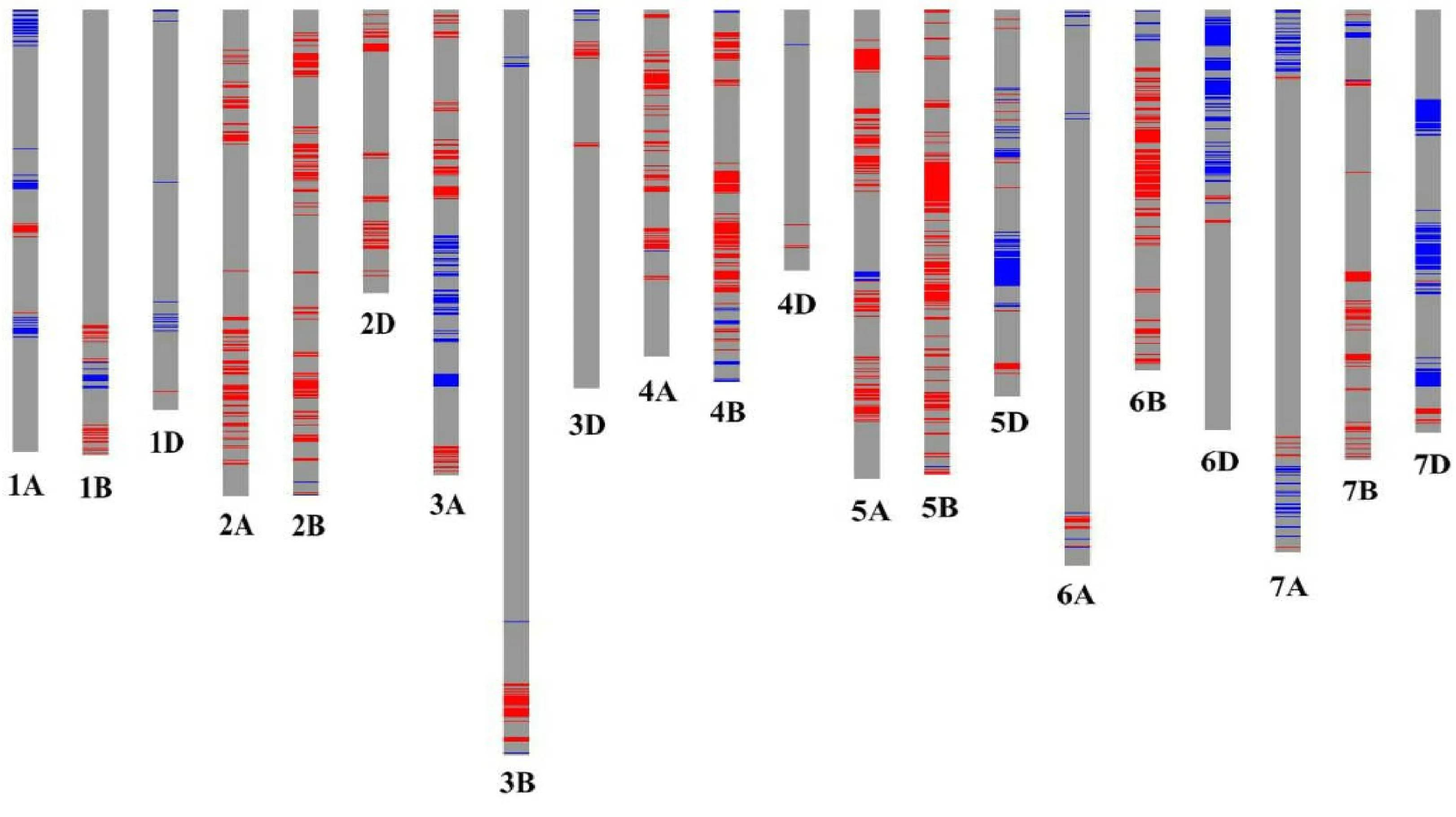
浅灰色表示三者相同区段,红色表示郑麦366区段,蓝色表示豫麦34区段。Light grey indicates identical fragment; Red indicates Zhengmai 366 fragment; Blue indicates Yumai 34 fragment.图1 郑品优9号的21条染色体SNP基因型图谱Fig.1 SNP genotype map on 21 chromosomes of Zhengpinyou 9
2.3 郑品优9号及其双亲重要性状的功能基因
利用小麦50K SNP育种芯片中的KASP标记对郑品优9号及其双亲的产量相关基因、品质性状相关基因、适应性相关基因和抗性相关基因进行分析,结果如下(表2)。
2.3.1 株高、籽粒等产量性状相关基因
郑品优9号及双亲株高的矮秆基因均为Rht-D1b基因,粒色为白粒TamybR_B1a基因,均含有芒基因AWN。共检测到13个与粒重或粒数相关的基因,郑品优9号与郑麦366在这13个基因的组成上完全一致,均聚合了8个高粒重基因(TaSus2-2A、TaSus2-2B、TaSus1-7A、TaSus1-7B、TaGW2-6B、TaGW6-2A、TaGS-D1和TaGS5)和1个高粒数基因 (TaMoc),与豫麦34只在TaGW2-6B基因存在差异。
2.3.2 品质性状相关基因
在郑品优9号及双亲间共检测到12个品质性状相关基因,3个品种的品质基因组成一致性很高。其中,高分子量谷蛋白亚基Glu-A1和Glu-D1位点均为优质亚基1和5+10,低分子量谷蛋白亚基Glu-A3和Glu-B3位点组成均为Glu-A3a和Glu-B3e,且均含有硬质基因Pina-D1b、高多酚氧化酶基因PPO-A2c、高黄色素含量基因Psy-B1c和Psy-D1g、高蛋白含量基因GPC-Hap-H以及能增加籽粒蛋白质积累的基因NAM-6A1b。此外,郑品优9号和郑麦366含有高黄色素含量的等位基因Pds-B1a,而豫麦34含有低黄色素含量的等位基因Pds-B1b。
2.3.3 春化、光周期、开花等适应性相关基因
在郑品优9号及双亲中共检测到6个春化基因、1个光周期基因和2个开花基因(表2)。其中郑品优9号和母本郑麦366在9个基因组成上完全一致,与豫麦34仅主效春化基因Vrn-B1存在差异。
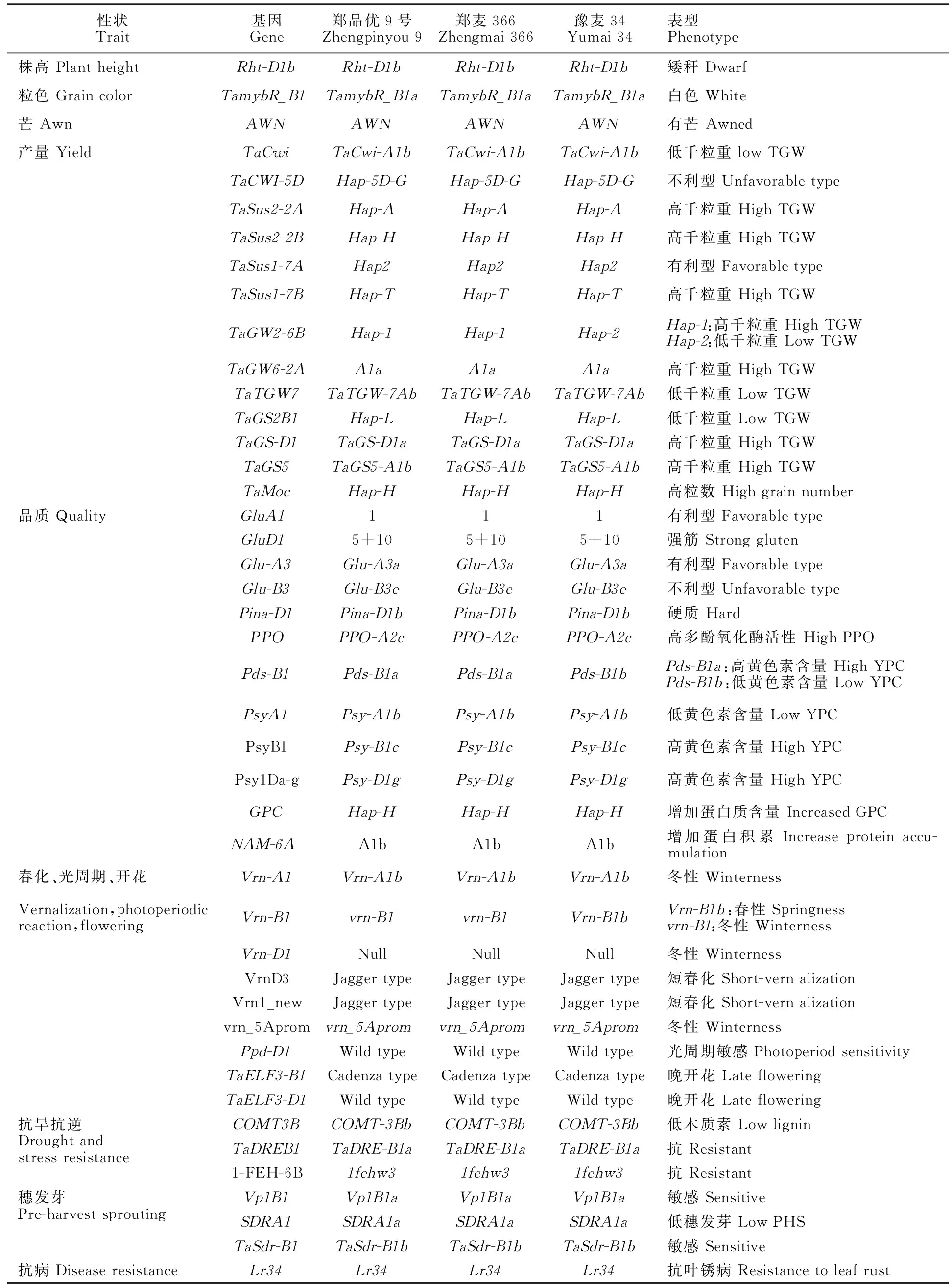
表2 郑品优9号及双亲重要性状相关基因分型Table 2 Genotyping of important traits in Zhengpinyou 9 and its parents
2.3.4 抗性相关基因
共检测到3个小麦抗旱、抗逆相关基因 (COMT3B、TaDREB1和1-FEH-6B)、3个抗穗发芽基因(Vp1B1、SDRA1和TaSdr-B1)和1个抗叶锈病基因(Lr34)。郑品优9号和双亲在这7个基因组成上完全一致。
3 讨 论
利用小麦50 K SNP育种芯片对郑品优9号及其父母本进行分子检测,揭示了郑品优9号的分子遗传构成,发现郑品优9号大部分的遗传物质来源于母本郑麦366(68.26%)。这与前人利用分子标记对小麦骨干亲本或主栽品种在后代中的遗传物质多发生偏亲现象一致[7-9,21-22]。如周麦23中63.31% 的遗传物质来源于母本周麦13(SSR标记)[7],淮麦33中73.9%的遗传物质来源于母本烟农19(SSR标记)[8],青农2号大部分遗传信息来源于鲁麦14(SSR 标记:54.11%;SNP 标记:72.55%)[9],周麦16中64.32%的遗传物质来源于周8425B[21]。Bernardo等[22]研究表明,单交亲本对后代的遗传贡献率为26%~74%,与本研究的结果一致。在单交后代中普遍出现子代遗传物质偏向于某一亲本的主要原因,可能与育种家的育种目标和亲本选用有关。育种家在配制单交组合时,常选择骨干亲本或当地主栽品种作为亲本之一,一般这些骨干亲本或主栽品种较另一亲本更适应当地区域生态条件,在后代选育时,选择符合当地生态区域条件的优良变异类型是重要的育种目标,因此后代品种保留了更多的骨干品种或主栽品种的遗传物质。前人研究中的郑麦366、烟农19、鲁麦14和周8425B等均是当地主栽品种或骨干亲本,较另一亲本具有更多的优良遗传位点,具有更好的适应性,经人工选育的后代品种遗传物质也偏向这类品种。
本研究发现,在基因组和染色体水平上,双亲对郑品优9号的遗传贡献率差异均较大。郑麦366对郑品优9号A、B和D基因组的贡献率分别为68.60%、90.98%和27.63%,而豫麦34的贡献率分别为31.40%、9.02%和72.37%;郑麦366对郑品优9 号在不同染色体上的遗传贡献率范围为7.53%~99.43%,在2A、4A、5A、2B、3B、4B、5B、6B、7B和2D染色体上均超过80%;而豫麦34在不同染色体上对郑品优9号的遗传贡献率范围为0.57%~92.47%,在7A、1D、6D和7D染色体上均超过80%。出现这种现象的原因可能也与育种家的育种目标选择有关。
本研究发现,郑品优9号聚合了多个优异基因,含有矮秆基因(Rht-D1b)[23-24]、高千粒重基因(TaSus2-2A、TaSus2-2B、TaSus1-7A、TaSus1-7B、TaGWb-2A、TaGS-D1和TaGS5[25]、高粒数基因(TaMoc)[25]、抗旱基因(TaDREB1a和1fehw3)[26-27]、低穗发芽基因(SDRA1a)[28]、抗叶锈病基因(Lr34)[29]、高蛋白含量基因(GPC-Hap-H)以及有利籽粒蛋白质积累的基因 (NAM-6A1b)[30-31]。正是由于以上基因的聚合效应,郑品优9号表现出半冬性、矮秆、稳产、优质强筋以及一定的抗逆抗病性和适应性等特点,与审定报告性状特征相一致。另外,郑品优9号含有高多酚氧化酶活性基因(PPO-A2c)和高黄色素含量基因 (Psy-B1c和Psy-D1g)[32-33],使得其面粉色泽白度不够,影响面制品的外观品质,因此降低黄色素含量是今后该品种改良的一个目标。本研究也发现,郑品优9号的有利抗病抗逆基因和产量基因仍不够多,表明其在产量水平和抗病、抗逆方面仍有很大的改良空间。
