盐城沿海海堤林带柳莺属(Phylloscopus)鸟类秋季迁徙规律
2021-10-29安玉亭薛丹丹王立波鲁长虎
刘 彬,安玉亭,薛丹丹,王立波,许 鹏,鲁长虎,*
1 南京林业大学生物与环境学院,南京 210037 2 江苏省大丰麋鹿国家级自然保护区,盐城 224136
迁徙鸟类每年在繁殖地和越冬地之间往返,以应对季节性的环境变化,从而提高繁殖成功率和存活率[1]。但多数鸟类不能一次性飞达目的地,而需要在途中多次降落、停歇补充能量[2- 3]。因此鸟类到达中途停歇地的时间以及采取何种停歇策略,以确保完成迁飞过程就显得十分重要[4- 5]。学者关注小型雀形目鸟类在不同季节的迁徙节律、停歇时长、能量积累效率以及在不同年龄、不同性别、不同纬度之间的差异[6- 8]。研究表明中途停歇地的食物资源丰富程度及可获性往往决定小型雀形目鸟类停留时长:中途停歇地食物越丰富,越容易获得,则鸟类越倾向选择停留较长时间来补充能量,反之则倾向选择快速通过的策略[9- 10]。另一些研究发现,繁殖地或者越冬地的地理位置对小型雀形目鸟类迁飞途中的停歇规律具有影响:对春季莺类迁徙规律的研究表明,距离繁殖地越近则莺类通过中途停歇地的时间越早[11];对秋季途经墨西哥尤卡坦半岛的林鸟研究发现,繁殖地越靠北的森莺科鸟类越早到达中途停歇地[12];而对29种柳莺的迁徙节律研究表明,无论春季还是秋季,繁殖地越靠南的物种,其通过中途停歇地的时间越早[13]。迁徙时间最小化假说认为鸟类为了尽早到达繁殖地或者越冬地,会通过调整停歇策略来尽可能地缩短中途飞行或停歇时间,如提高迁飞速度以及减少在停歇地的逗留时间[14- 16]。在秋季从繁殖地到越冬地的迁飞过程中,鸟类为较快到达并占据质量较好的越冬地,会尽量减少在食物资源匮乏且不易获得的劣质停歇地逗留时间,以此平衡迁飞过程中能量消耗和能量补充之间的矛盾[17]。鸟类身体状况比如能量储备、脂肪量、飞行能力等的差异,也会导致到达中途停歇地时间的不同,经验越丰富且能量储备越高的个体,常常能够迅速到达停歇地[18]。
许多小型雀形目鸣禽如柳莺属鸟类具有长距离迁徙习性[19-20]。我国有柳莺属鸟类41种[21]。每年春季它们从越冬地向北迁飞,到我国东北、西伯利亚及北欧北部等地繁殖,秋季则从繁殖地飞往印度尼西亚、马来西亚等区域越冬[22- 24]。国内对柳莺属鸟类的研究报道总体较少,如对黄眉柳莺(Phylloscopusinornatus)、淡脚柳莺(P.tenellipes)繁殖生态学的研究[25- 26]。学者对黑龙江省嫩江高峰林区莺科鸟类春秋季迁徙期的种类、数量、迁徙规律的研究表明,黄眉柳莺为环志捕获优势种,相较于极北柳莺(P.borealis)和淡脚柳莺,秋季黄眉柳莺到达时间最晚[19]。盐城湿地位于江苏东部沿海,这里地形平坦开阔,缺乏成片的自然林地,沿海堤两侧栽植的海堤林是这里仅有的条带状林地,因此海堤林对依赖林地栖息或者迁徙的雀形目鸟类具有极其重要的作用。近年来通过监测发现,每年春季和秋季迁徙季节有很多鸟类经过江苏东部沿海海堤林[27]。条带状分布的海堤林为开展柳莺属迁徙规律研究提供了非常理想的场所。本研究中,我们采用雾网法对不同年份秋季途经海堤林的柳莺属种类、数量进行记录,分析优势种柳莺采取何种策略经过盐城海堤林,并对其身体状况随时间的变化进行对比分析,进而探讨盐城沿海海堤林带对小型雀形目鸟类迁徙的生态意义。
1 研究地点
研究地点位于江苏省大丰麋鹿国家级自然保护区(32°59′N—33°03′N,120°47′E—120°53′E)野放区附近的海堤林内。该保护区位于我国江苏黄海海滨,地处东亚—澳大利西亚鸟类迁徙路线,为典型沿海滩涂湿地。其气候特点为冬季干旱少雨,夏季高温降雨丰富。海堤林两侧地势开阔,生境类型包括互花米草(Spartinaalterniflora)盐沼、碱蓬(Suaedaglauca)草滩和潮间带光滩。保护区野放区生活着约1000头野生麋鹿(Elaphurusdavidianus),主要采食互花米草、芦苇(Phragmitescommunis)以及海堤林内灌木和草本植物。
所选海堤林始建于20世纪90年代,位于保护区野放区的西侧,南北长约8 km。海堤林内人工栽植乔木树种主要为刺槐(Robiniapseudoacacia)和少量加拿大杨(Populuscanadensis)、女贞(Ligustrumlucidum),高度9—15 m,因台风影响致使乔木密度较为稀疏;灌木层植被稀少,主要有蔷薇(Rosamultiflora)、枸杞(Lyciumbarbarum),另外还包含少量构树(Broussonetiapapyrifera)、苦楝(Meliaazedarach)等乔木树种的幼苗,呈斑块状分布,高度在1—4 m;草本层主要包括白茅(Imperatacylindrica)、葎草(Humulusscandens)、乌蔹梅(Cayratiajaponica)、野艾蒿(Artemisialavandulifolia)等。随着麋鹿采食压力增大,海堤林内灌木和草本植物数量明显减少。
2 研究方法
2.1 鸟类捕捉与测量
在获得盐城市自然资源和规划局审批许可之后(审批编号:盐林动审[2018]4号和盐林动审[2019]3号),采用雾网法捕捉途经海堤林的鸟类。沿海堤林每隔300 m设置一个网场,共设置4个网场。每个网场布设2张雾网,方向垂直于海堤林,雾网上下相连,每张雾网高2.6 m,长12 m,网眼大小36 mm2,雾网下纲线距地面0.1 m。选择无风晴朗天气, 6:00开始张网至17:00闭网,每间隔1 h巡网一次。
记录捕获鸟类的种类和数量、所在网号和入网时间,装入鸟袋送至附近的环志站进行测量,环志后放飞。用天平称量鸟类体重(精度±0.1 g),用直尺测量翅长(最大翅长法,±0.1 cm)和体长(cm)。采用8级脂肪度划分鸟类可见脂肪度,0级表示鸟类体内无可见脂肪,8级表示可见脂肪含量最高[28]。
去除大风、降雨等天气,2018—2019年秋季合计有效张网72 d,每年分别为37 d和35 d,历年网场布设地点和数量完全一致。2017年我们在研究地内进行了预实验,能够较为准确地掌握柳莺属鸟类最早到达和最晚离开的时间,并在2018年和2019年每月进行鸟类监测,准确掌握海堤林内鸟类迁徙的起止时间。2018年—2019年秋季,盐城海堤林带最早捕获到迁徙柳莺属鸟类的时间为9月3日,最晚日期为10月31日。因此我们把9月3日定义为秋季迁徙第1天,4日为第2天,依次类推10月31日为秋季第59天。
2.2 数据统计
(1)鸟类日捕获率
将4种优势柳莺属鸟类每日捕获数量按年份进行排列,以非参数检验(K-W test)比较同一年份内不同物种间平均日捕获率是否存在差异,以Mann-WhitneyUtest对同一物种的不同年份平均日捕获率进行比较。
(2)捕获数量随时间的变化
按年份将4种优势柳莺属鸟类捕获数量以日期顺序排列,把某种柳莺最早捕获日期至最晚捕获日期作为最短停留时间。用非参数检验(K-W test)对同一年份的4种优势柳莺属鸟类迁徙时间进行比较,并用Mann-WhitneyUtest对任意两种柳莺的迁徙时间进行比较。
(3)体重和可见脂肪度
采用鸟类体征参数CI(condition index)来表示鸟类相对体重大小[29]:
CI=M/W,其中M为体重(g),W为翅长(cm)
以日为单位,将每种柳莺的体征参数CI与可见脂肪度进行线性拟合,分析两者之间的关系;然后分析每种柳莺可见脂肪度大小与迁徙时间的关系。
本文数据处理、制图用SPSS 21.0和Origin 8.5软件完成,数据以平均数±标准差(Mean±SD)表示,显著度大小采用P=0.05水平。
3 研究结果
3.1 种类组成与数量
2018—2019年秋季共捕获雀形目柳莺科柳莺属鸟类8种,分别为极北柳莺、冕柳莺(P.coronatus)、淡脚柳莺、黄眉柳莺、双斑绿柳莺(P.plumbeitarsus)、黄腰柳莺(P.proregulus)、巨嘴柳莺(P.schwarzi)和褐柳莺(P.fuscatus)。其中前4种柳莺捕获数量较多,为秋季海堤林内的优势种(图1)。其数量分别为:极北柳莺数量最多,为245只,其次为冕柳莺、淡脚柳莺和黄眉柳莺,数量分别为131只、107只和94只。这4种优势柳莺捕获数量占柳莺属捕获总数的95%(图2)。

图1 秋季盐城海堤林带4种优势柳莺属鸟类Fig.1 4 dominant warbler species in Yancheng seawall forest belt in autumn
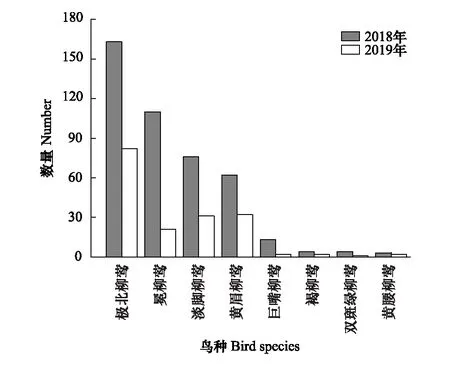
图2 秋季盐城海堤林带柳莺属鸟类组成及捕获数量 Fig.2 Species and captured number of warblers in Yancheng seawall forest belt in autumn
对4种优势柳莺日捕获率进行比较:2018年4种柳莺的平均日捕获率无明显差异(K-W test:χ2=0.723,df=3,P>0.05);2019年则存在差异(K-W test:χ2=8.794,df=3,P<0.05),进一步分析表明:2019年极北柳莺与淡脚柳莺(Mann-WhitneyUtest:Z=-2.030,P<0.05)、黄眉柳莺(Mann-WhitneyUtest:Z=-2.691,P<0.01)的平均日捕获率之间存在显著差异。2018年与2019年两个年份间4种柳莺的平均日捕获率无明显差异(Mann-WhitneyUtest,极北柳莺:Z=-0.318,P>0.05;冕柳莺:Z=-0.562,P>0.05;淡脚柳莺:Z=-1.150,P>0.05;黄眉柳莺:Z=-1.530,P>0.05)(表1)。
3.2 种群数量时序变化
4种柳莺捕获数量随迁徙时间呈现不同的变化规律:2018年4种柳莺都较早到达盐城海堤林内(9月4日—7日),其中极北柳莺、冕柳莺和淡脚柳莺50%的种群个体在9月中上旬到达,黄眉柳莺则较晚(10月9日)。冕柳莺的个体在9月23日前全部通过海堤林带,其他3种柳莺全部种群通过海堤林带延至10月下旬(图3)。2019年4种柳莺的迁徙时序与2018年基本相似,在9月上旬都已经出现在海堤林内,极北柳莺、冕柳莺和淡脚柳莺50%的种群个体在9月12日前到达,黄眉柳莺较晚(9月27日)。冕柳莺个体在9月16日前全部通过海堤林带,而淡脚柳莺、极北柳莺和黄眉柳莺全部个体则先后通过海堤林带(图3)。4种柳莺通过盐城海堤林带的时间在年间基本保持一致。

表1 不同年份秋季盐城海堤林带4种优势柳莺日捕获率差异

图3 不同年份秋季盐城海堤林带4种优势柳莺种群数量时序变化Fig.3 Number changes of 4 dominant warbler species with time in Yancheng seawall forest belt in autumn从9月3日算起的天数
极北柳莺为盐城海堤林带秋季常见鸟类,2018年和2019年秋季共捕获245只。在研究地极北柳莺最早在9月4日被捕捉到,10月24日之后再捕捉不到,估测秋季极北柳莺种群通过研究地的最短时间49天。年份间略有不同:2018年极北柳莺迁徙集中在秋季第3天到第21天,先后出现4个高峰,最高峰出现在秋季第3天,达到41只;2019年极北柳莺迁徙集中在秋季第3天到第15天,先后出现5个高峰,最高峰出现在秋季第10天,数量达到13只(图4)。不同年份间的秋季极北柳莺通过盐城沿海海堤林时,基本采取相对集中时间通过的模式。
2018年和2019年秋季共捕获冕柳莺131只。在研究地冕柳莺最早在9月4日被捕捉到,9月23日之后再捕捉不到,估测秋季冕柳莺通过研究地的最短时间为19天。不同年份间,冕柳莺迁徙节律基本相同,迁徙集中于9月上旬(4日—14日),数量最高峰出现在2018年的9月11日,为92只(图4)。冕柳莺通过盐城沿海海堤林采用相对集中时期通过模式。
2018年和2019年秋季共捕获淡脚柳莺107只。在研究地淡脚柳莺最早在9月4日被捕捉到,10月25日之后再捕捉不到,估测淡脚柳莺通过研究地的最短时间为52天,迁徙集中于9月6日至14日。年份间:2018年淡脚柳莺迁徙集中在秋季第4—10天(9月6—12日),2019年迁徙集中在秋季第9—12天(9月11—14日),最高峰出现在2018年的9月12日,达到30只(图4)。淡脚柳莺通过盐城沿海海堤林采用相对集中时期通过的模式。
2018年和2019年秋季共捕获黄眉柳莺94只。在研究地黄眉柳莺最早从9月7日被捕捉到,10月27日之后再捕捉不到,估测黄眉柳莺通过研究地的最短时间为51天。2018年迁徙集中在秋季第30天(10月2日)、第37—39天(10月9—11日)和第52—55天(10月24—27日);2019年迁徙集中在秋季第5—21天(9月7—23日)和第46—48天(10月18—20日)(图4)。虽然在不同年份间迁徙节律略有差异,但黄眉柳莺通过盐城沿海海堤林基本采取分段集中通过的模式。

图4 秋季盐城海堤林带4种优势柳莺捕获数量随时间的变化Fig.4 Timing of captured number of 4 dominant warbler species in Yancheng seawall forest belt in autumn
3.3 不同种类的迁徙时序
对不同年份4种优势柳莺的迁徙时序进行比较: 2018年秋季4种柳莺的迁徙时序存在极显著差异(K-W test,χ2=19.015,df=3,P<0.01)。进一步分析表明,黄眉柳莺在迁徙时序上要明显晚于其他3种柳莺(Mann-WhitneyUtest:①极北柳莺与黄眉柳莺,Z=-2.890,P<0.01;②冕柳莺与黄眉柳莺,Z=-3.672,P<0.01;③淡脚柳莺与黄眉柳莺,Z=-3.088,P<0.01),而冕柳莺略早于淡脚柳莺和极北柳莺,但是差异不明显(Mann-WhitneyUtest,P>0.05)(图5)。2019年4种柳莺的迁徙时序存在显著差异(K-W test,χ2=9.775,df=3,P<0.05),进一步分析,黄眉柳莺在迁徙时序上要明显晚于淡脚柳莺(Mann-WhitneyUtest,Z=-2.978,P<0.01),与极北柳莺(Mann-WhitneyUtest,Z=-1.273,P>0.05)、冕柳莺迁徙时序差别不大(Mann-WhitneyUtest,Z=-1.214,P>0.05)。冕柳莺在迁徙时间上要早于极北柳莺(Mann-WhitneyUtest,Z=-2.209,P<0.05)和淡脚柳莺(Mann-WhitneyUtest,Z=-2.304,P<0.05)(图5)。
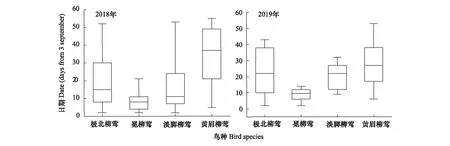
图5 不同年份秋季盐城海堤林带4种优势柳莺迁徙时序Fig.5 Migration timing of 4 dominant warbler species in Yancheng seawall forest belt in autumn
3.4 体重与可见脂肪度的变化
4种柳莺的可见脂肪度与体征指标CI之间都存在极显著正相关关系(极北柳莺:R=0.720,P<0.01;冕柳莺:R=0.703,P<0.01;淡脚柳莺:R=0.664,P<0.01;黄眉柳莺:R=0.791,P<0.01)。同一物种内,柳莺个体的可见脂肪度越大,鸟类体征指标CI值越大,即鸟类的相对体重越大,因此可见脂肪度的大小能够较好地反映柳莺的相对体重状况。
4种柳莺可见脂肪度含量在到达盐城沿海海堤林时普遍较低(极北柳莺:2.49±0.87,n=245; 冕柳莺:3.30±1.17,n=131;淡脚柳莺:3.23±0.88,n=107;黄眉柳莺:2.29±1.43,n=94)。随着时间的推移,柳莺的平均可见脂肪度呈显著下降的趋势(General Linear Model,P<0.05)(图6),即在秋季迁徙期内,后期到达研究地内的柳莺个体相对体重较前期到达的明显偏小,即身体状况也相对较差。

图6 秋季盐城海堤林带4种优势柳莺平均可见脂肪度的时间变化Fig.6 The relationship between migration date and average fat score of 4 dominant warbler species in Yancheng seawall forest belt in autumn
4 讨论
盐城东部沿海位于东亚—澳大利西亚鸟类迁徙路线上,每年春季和秋季都会有大量候鸟途经这里,并降落停歇补充能量,学者对迁徙期鸻鹬类鸟类的关注程度较高[30- 31]。许多小型雀形目鸟类也将盐城沿海作为中途停歇地或迁徙通道,生境单一但却稀有的林地生境—沿海海堤林带对其完成迁徙生活史具有重要意义。由于小型雀形目鸟类辨识难度大且行为隐蔽,人们对迁徙经过沿海海堤林带的小型雀形目鸟类的研究近乎空白。本文首次尝试用雾网法对秋季途经盐城沿海海堤林带的柳莺属鸟类进行研究,发现并捕获8种长距离迁徙的柳莺属鸟类,占我国柳莺属种类的20%。这说明盐城沿海海堤林带可能是雀形目柳莺属鸟类重要的迁徙途经路线。
对朝鲜半岛极北柳莺的迁徙规律研究发现,最早见到极北柳莺的时间为8月中旬,最晚为10月上旬[23]。这与我们的研究略有差异,秋季盐城沿海海堤林带最早见到极北柳莺的时间为9月上旬,最晚10月下旬离开,这比途经朝鲜半岛的极北柳莺要晚。这可能与两个研究地的地理位置有关:极北柳莺的繁殖地在北欧以北、北极圈附近,越冬地在东南亚、菲律宾和印度尼西亚等地[22-23]。江苏盐城比朝鲜半岛更靠南,秋季极北柳莺从北部繁殖地向南部越冬地迁徙,到达盐城时间要晚于朝鲜半岛。不同性别、年龄是否会对鸟类的迁徙节律产生影响,有的认为存在某种差异,比如雄性早于雌性,有的则认为没有差异[32- 33]。本文捕获的柳莺属鸟类迁徙节律的差异是否由于性别和年龄的差异所导致,需要在今后的研究中进一步细化和明确。
迁徙中途停歇地质量决定迁徙鸟类能否快速找到食物,进而迅速完成能量补充,因此栖息地内食物丰富程度、取食难易程度是决定鸟类选择不同停歇策略的关键[34]。当中途停歇地的食物资源不充足且难以获得时,鸟类通常会选择快速通过的策略,赶往下一站选择更加优质的停歇地[9]。盐城沿海海堤林带建群种为人工林,植被层次较为单一,且缺乏适宜昆虫栖息的灌木层[27],导致林内昆虫资源较少。调查发现秋季迁徙期盐城沿海海堤林内仅有少量的鳞翅目幼虫,对柳莺属鸟类来说,昆虫食物比较缺乏且不易捕食,因此海堤林带可能不是柳莺属鸟类最佳停歇地。通过对四种柳莺迁徙时间节律进行分析,4种柳莺大多数个体通过盐城沿海海堤林带的时间较短,采取集中或者分段集中快速通过的迁徙策略,从而减少在食物资源相对匮乏的盐城沿海海堤林内的逗留时间,以便能够快速的朝越冬地方向迁飞。这种迁徙策略符合鸟类迁徙时间最小化理论。研究期内4种优势柳莺的重捕率较低(0.9%),这侧面反映盐城沿海海堤林带可能仅是柳莺属鸟类的迁徙通道,而并非优质的停歇栖息地。
雀形目鸟类迁徙期主要以可见脂肪度的形式在身体内储存能量,其大小与鸟类的体重和翅长等身体指标相关[5]。我们的结果显示,四种优势柳莺的可见脂肪度都与体征指标CI呈正相关。对某些莺类迁徙规律进行研究发现,身体脂肪度储备较低的个体与较高的个体相比,会在优质的中途停歇地停留较长的时间,以补充足够能量,这导致前者会晚一些到达下一个中途停歇地[35- 37]。这与我们的结果相一致:身体能量储备较高的个体,凭借较强的飞翔能力和能量积累,能够较早到达中途停歇地,而能量储备较低的个体,由于需要在上一个适宜的停歇地花费大量时间补充能量,导致迁徙时间滞后。
综上可知,盐城沿海海堤林带一方面能够为秋季迁徙的柳莺属鸟类提供隐蔽地和通道,另一方面由于海堤林内植被结构单一,灌木层的缺乏使昆虫资源相对匮乏且难以获取。因此盐城沿海海堤林带可能仅是雀形目柳莺属鸟类迁徙的重要通道之一,而非优质的中途停歇地。
