秦山核电站邻近海域生态环境长期变化
2021-10-24叶文琪刘翰仁李尚鲁廖一波陈悦汤雁滨陈全震寿鹿杜萍
叶文琪 ,刘翰仁 ,李尚鲁 ,廖一波 ,陈悦 ,汤雁滨 ,陈全震,寿鹿,杜萍
(1.自然资源部第二海洋研究所自然资源部海洋生态系统动力学重点实验室 杭州 310012;2.浙江省海洋监测预报中心 杭州 310007)
0 引言
在生态文明约束下,清洁能源在电力供应中占比不断提高[1]。核电作为一种清洁能源,是控制世界温室气体排放的最重要措施之一。按照《核电中长期发展规划(2005—2020 年)》,到2020 年,我国总投产核电容量将达到4 000万k W,年发电量达到2 600亿k W·h,占全国发电量6%以上[2]。目前,我国核电正在加速建设,滨海核电运行对海洋生态环境的影响备受关注。
正常条件下,核电的环境影响主要还是温排水对水生生物的影响以及冷却水取水导致的生物卷吸效应[3]。美国从20 世纪60 年代开始监测温排水和杀生剂对水生生物的影响,分析取水撞击和夹带导致的生物损失,结果显示,闭式循环冷却水系统因温排水量小而对水生生物的负面影响小[4];直流冷却水系统会改变扩散条件差的受纳水体(封闭、半封闭的海湾、浅滩等)或低纬度海域的水温环境,水体温度升高或昼夜热变化规律改变将导致生物群落多样性、丰度和生态系统功能改变[3,5-6]。我国滨海核电厂均采用直流冷却水系统。2000年以来国内开展了大量温排水生态效应的实验和原位短期研究,而长期研究仅见于大亚湾核电站[7]。魏新渝等[7]及所引用文献认为,大亚湾核电温排水导致邻近海域浮游植物群落结构变化,春、冬季中华哲水蚤密度降低,大型底栖动物生产力水平显著下降,斑和鲷科鱼类产卵地迁移等。然而,王友绍等[8]依据30 余年生态监测数据,综合多种因素认为,大亚湾核电站温排水对周围海域生态环境存在一定影响,但不会影响大亚湾生态系统的变化趋势,无机氮磷比(TIN∶P)是关键驱动因子。可见,基于长期数据的综合评估核电生态效应对于正确判断核电的生态影响至关重要。
秦山核电站是我国运行最早、规模最大的核电站,濒临强潮河口杭州湾,水交换条件良好;大亚湾为半封闭海湾,稀释扩散条件较差。研究秦山核电站邻近海域生态环境长期变化,与大亚湾核电站对比,更有利于为我国核电的选址和设计提供参考。同时,杭州湾海域生态环境健康发展事关大湾区建设和国家长三角一体化发展,摸清电厂的生态效应是研究杭州湾海域演变机制的重要组成部分。但秦山核电站邻近海域生态环境长期变化罕有研究,本研究比较1989年和1990年、1995年和1996年[9-25]、2010年和2012年[26-27]邻近海域和潮间带的生态调查结果,探讨核电运行对生态环境的长期影响。
1 材料与方法
1.1 研究区域及电厂概况
秦山核电站是中国自行设计、建造和运营管理的第一座核电站。一期工程机组装机容量32万k W,1994年4 月投入商业运行,与大亚湾核电站投运(1994年2月)几乎同时。秦山核电站二期1号-4号机组装机容量均为65 万k W,分别于2002 年4月、2004年5月、2010年10月和2011年12月投入商业运行。三期1 号和2 号机组装机容量均为72.8万k W,分别于2002年12月和2003年7月投入商业运行。目前,秦山核电基地(含方家山核电工程)共9台机组投产发电,总装机容量达656.4万k W,年发电量约500亿k W·h,成为国内核电机组数量最多、堆型最丰富、装机最大的核电基地。秦山核电的温排水排放达到520 m3/s,超过中国多数河流的径流量;排水口温排水温升达到8℃[26]。
秦山核电站坐落于杭州湾北岸的浙江省嘉兴市海盐县。杭州湾属强潮河口,平均潮差531 cm,最大可能潮差985 cm;潮汐为非正规浅海半日潮,涨落潮将钱塘江径流水和湾外海水充分混合;受S型往复流的作用,海水盐度一直维持着河口低盐水平;平均水深8~10 m;具低水色、低透明度、高悬浮体浓度、高营养盐以及低生物生产力等生态特征[11,16,28]。受亚热带季风影响和海洋气流调节,杭州湾的温盐具有明显的季节和年际变化,降雨主要集中在4-9月,占年降水量69%,多年平均降雨量在900~1 500 mm[28]。海底底质以粉砂为主,多为坚实的“铁板砂”,由于潮流的往返冲刷,沉积环境极不稳定,底栖生物贫乏[20]。
1.2 数据来源与分析
秦山核电站运行前期(1989—1990年)和运行期间(1995—1996年和2010—2012年),自然资源部第二海洋研究所(原国家海洋局第二海洋研究所)在厂址邻近海域和潮间带开展了生态环境调查(图1)。海域调查时间分别为1989年4月(春季)、7月(夏季)、10月(秋季)、1990年1月(冬季);1995年10月、1996年1 月、1996 年4 月、1996 年7 月、2010 年10月、2012年5月。潮间带调查时间分别为1985年4月、9月和2010年10月、2012年5月大潮期间。海域生态环境调查内容包含叶绿素a、网采(水采)浮游植物、大中型浮游动物和大型底栖生物,并同步测定海水温度、盐度、p H 和悬浮物、溶解氧、营养盐浓度,以及沉积物有机质含量等理化参数。叶绿素a测定用淬灭荧光法。网采浮游植物采用浅Ⅲ浮游生物网(网口内径37 cm,网口面积0.1 m2,网长140 cm,筛绢孔径为77μm)自底至表作垂直拖网采集,水采浮游植物采集500 mL表层海水,显微镜鉴定和计数。大中型浮游动物采用浅I型浮游生物网(网口内径50 cm,网口面积0.2 m2,网长145 cm,筛绢孔径为505μm)从底至表层垂直拖网采集,体视镜和显微镜下鉴定和计数,天平称量。大型底栖生物使用0.1m2的采泥器采样,经0.5 mm 孔径套筛冲洗出生物样品,体视镜和显微镜下鉴定和计数,天平称量。
本研究利用3个年代的海域调查数据分析秦山核电站邻近海域海洋生物长期变化;利用2个年代的潮间带调查数据分析核电站邻近潮间带生物长期变化。
2 结果与讨论
2.1 理化环境变化
2.1.1 水体环境变化
秦山核电邻近海域的温盐影响因素多、波动大(表1、表2)。夏、秋、冬季的温度年际差异较小,春季有升高趋势,但难以判定电厂温排水对春季温度的影响。因为2012年春季调查时间是5月,其他年份是4月,故2012年春季水温明显偏高,而且1996年春季调查期间连续阴雨,温度偏低,故无法确定温升是温排水的影响。盐度受径流、降雨和海水入侵影响,径流影响最明显[11]。1996年春季连续阴雨,径流和降雨较大,盐度较低;其他季节降雨少,外海水入侵势力增强导致盐度高于1989年春季同期[24];杭州湾5月降水明显高于4月[28],因此2012年春季盐度低于其他年份。该海域海水混浊、悬浮泥砂含量高的特征未变化(表3);高溶解氧含量和海水弱碱性环境也基本稳定(表4和表5)。

表1 海水温度历史变化[9-11,29] ℃

表2 海水盐度历史变化[9-11,29] ‰

表3 海水悬浮物历史变化[9-10,12-13,29] mg/L

表4 海水溶解氧浓度历史变化[9-10,12-13,29] mg/L

表5 海水p H 历史变化[9-10,12-13,29]
秦山核电邻近海域具有高营养盐浓度、高氮磷比(N/P)特征[30]。3个年代数值对比,硝酸盐、磷酸盐和硅酸盐浓度持续升高,亚硝酸盐和铵盐浓度呈先升高、后下降趋势(表6至表10)。无机氮主要来源于工农业及生活污水,磷酸盐来源多样,硅酸盐主要来源于土壤风化。受环杭州湾区工农业发展影响,近30年,湾内外营养盐浓度升高明显,其中无机氮增幅最大,主要与沿岸农田大量使用氮肥有关[30]。同时,杭州湾营养盐浓度年际变化也受降雨、钱塘江径流和长江冲淡水影响。
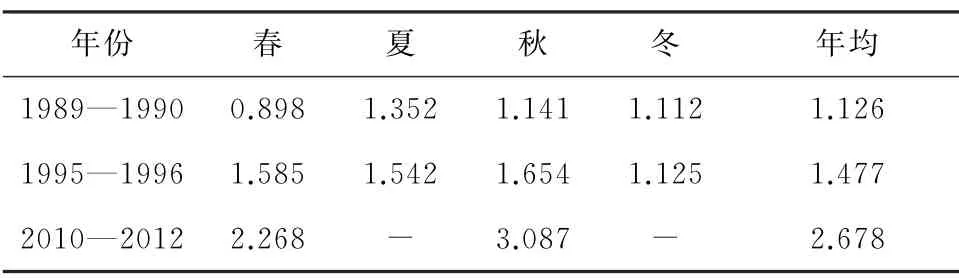
表6 海水硝酸盐历史变化[9-10,12,14,29] mg/L

表7 海水磷酸盐历史变化[9-10,12,14,29] mg/L

表8 海水硅酸盐历史变化[9-10,12,14,29] mg/L

表9 海水亚硝酸盐历史变化[9-10,12,14,29] mg/L
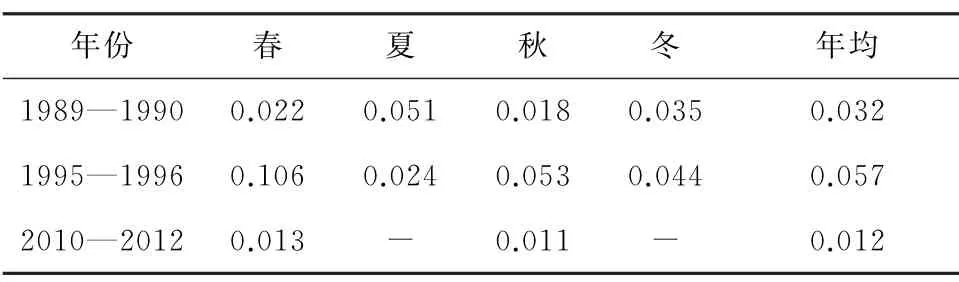
表10 海水铵盐历史变化[9-10,12,14,29] mg/L
2.1.2 沉积物环境变化
秦山核电邻近海域1989—1990 年和1995—1996年沉积物有机碳年均含量分别为0.46%[9]和0.39%[10];2010—2012年沉积物有机碳含量均值为0.62%[29]。可见,该海域沉积物中有机碳含量低,但略呈上升趋势。
2.2 叶绿素a和浮游植物
叶绿素a浓度是浮游植物细胞密度的反映,受温、盐、光照和营养盐浓度直接影响。杭州湾海域营养盐丰富,溶解氧充足,但由于水体透明度低及往复潮流强烈冲刷,初级生产力和叶绿素a含量低[16]。1995—1996年降雨量小、径流弱,调查区盐度高于1989—1990年(除春季),携带高悬浮泥沙含量的海水进入,使透光率下降,限制浮游植物的生长繁殖,导致1995—1996年叶绿素a浓度和浮游植物细胞密度(水样)明显低于1989—1990年;同时,春季连绵阴雨,温度低也是原因之一。与1989—1990年相比,2012 年春季温度高、盐度和悬浮物浓度低,硝酸盐和磷酸盐浓度为1989—1990年同期2倍以上,促使叶绿素a浓度明显高、浮游植物细胞密度(网样)约为1989—1990年的9倍;2010年秋季温度稍高、悬浮物浓度稍低,硝酸盐为1989—1990 年同期的2.7 倍,叶绿素a与1989—1990 年基本持平、浮游植物细胞密度约为1989—1990年的3 倍(表11 至表13)。可见温度、盐度和光照是研究海区浮游植物密度年际波动的主要限制因素,但当温度、盐度和光照条件良好时,高营养盐会促进浮游植物旺发。陈悦等[27]发现2010—2012 年春、秋季电厂邻近海域网采浮游植物丰度分别为杭州湾1981—1982年同季的27倍和71倍,认为营养盐增加是主要原因。
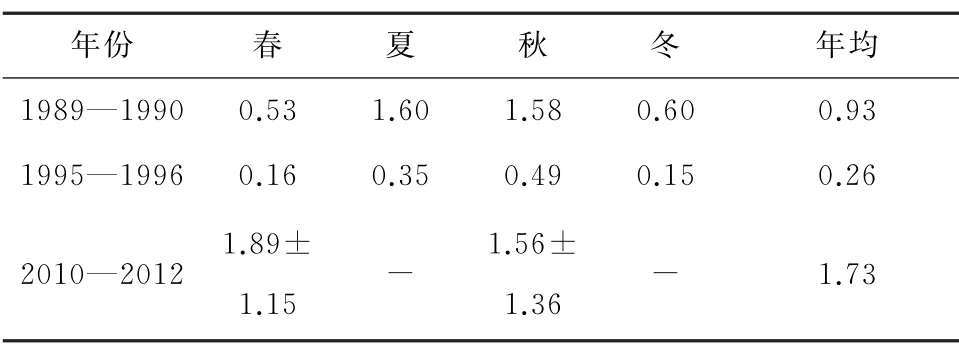
表11 表层叶绿素a浓度历史变化[9-10,16-18,23,27,29] μg/L

表12 浮游植物细胞密度历史变化[9-10,16-18,23,27,29] 万个/m3

表13 浮游植物种类数历史变化[9-10,16-18,23,27,29]
秦山核电邻近海域浮游植物种类主要为广布性近岸种、半咸水种和淡水种,也有少量暖水性外海种。浮游植物物种数年际波动大,基本与温度正相关、与盐度负相关,总种类数主要取决于淡水种的增减[10]。与1989年夏季相比,1996年夏季降雨量小、径流弱,淡水种减少60余种[10],导致总种类数(水样)下降明显。1995—1996年其他3季和2010年秋季、2012年春季物种数(网样)较1989—1990年同期均有不同程度的增加(表4),这与杭州湾海域1992—2012年的浮游植物种类数明显增加趋势一致[31]。
优势种也有变化。与1989—1990 年相比,2012年春季(网样)和1996年夏季(水样)具槽帕拉藻(Paralia sulcata)优势度提高;2010 年秋季(网样)弓束圆筛藻小型变种(Coscinodiscus curvatulus var.minor)增加。两者均为小型硅藻,小型硅藻占比增加的趋势也在大亚湾核电站附近海域观察到[32],这可能与海水升温有关[33-35]。此外,研究显示温升会提高暖水种琼氏圆筛藻(Coscinodiscus jonesianus)的细胞分裂速率[36],秦山核电邻近海域春、秋季其网采丰度分别由1989—1990 年的32.5万 个/m3和25.3万个/m3增长到 2010—2012年的47.7万个/m3和32.6万个/m3[9,27],且丰度与温度呈正相关,热羽区较高[27,29],由此推测,温升可能促进暖水种增加。
2.3 大中型浮游动物
浮游动物生物量和丰度年际波动明显(表14至表16)。浮游动物丰度受温度和盐度影响,通常与温度呈正相关、与盐度呈负相关。优势种变化改变浮游动物平均大小,从而与丰度共同影响生物量。1996年春季调查期间水温偏低,浮游动物丰度和生物量均较低;1995—1996年夏、秋、冬3季盐度明显高于1989—1990年同期,导致浮游动物丰度低,但因夏、秋季优势种长额刺糠虾个体较大,且有较多个体大的球型侧腕水母,导致夏、秋的生物量较1989—1990年反而略有上升[24]。2010年秋季浮游动物丰度、生物量和优势种变化完全符合以上规律;2012年春季优势种也符合以上规律,但2012年春季虽温度高、盐度低,浮游动物丰度依然最低,可能与理化环境的长期变化有关。象山港宁海电厂邻近海域多年研究发现,排水口附近的大中型浮游动物丰度较低,与温排水冲击导致原优势种迁移和死亡有关[37],也与长期高温胁迫导致浮游动物变小有关[38]。基于此推测,秦山核电站邻近海域春季浮游动物丰度降低可能与温排水有关——受到温排水长期影响,浮游动物个体变小,导致大中型浮游动物数量下降。

表14 浮游动物生物量历史变化[9-10,19,24,29] mg/m3

表15 浮游动物丰度平均值历史变化[9-10,19,24,29] ind/m3

表16 浮游动物物种数历史变化[9-10,19,24,29]
秦山核电邻近海域浮游动物优势种为半咸水河口种(虫肢歪水蚤、华哲水蚤、火腿许水蚤和江湖独眼钩虾)和近岸低盐种(长额刺糠虾和真刺唇角水蚤)。优势种组成及优势度的年际变化受盐度影响,随盐度升高,优势种由咸水河口种向近岸低盐种转变。与1989—1990年相比,1996年和2012年春季盐度较低,半咸水河口种虫肢歪水蚤优势度较高、近岸低盐种真刺唇角水蚤优势度较低;1995—1996年夏、秋、冬季盐度较高,半咸水河口种虫肢歪水蚤优势度较低、近岸低盐种长额刺糠虾和真刺唇角水蚤优势度较高;2010 年秋季优势种与1989—1990年一致。因此,浮游动物优势种基本稳定。
2.4 大型底栖动物
秦山核电邻近海域底质以粉砂为主,由于潮流的往返冲刷,沉积环境极不稳定,多为坚实的铁板砂,有机质含量少,底栖动物生物量和栖息密度均属较低水平[20]。但3个年代的大型底栖动物物种数、生物量和栖息密度持续增加(表17 至表19)。1995—1996年与1989—1990年的优势种均为多鳃齿吻沙蚕和焦河蓝蛤,2010—2012 年优势种中,焦河蓝蛤未变,但是多鳃齿吻沙蚕不再是优势种。大型底栖生物种类、数量和钩虾增加与人类活动有关,围填海、码头建设等工程改变海域水动力环境,从而影响底质类型和环境;同时,海域微藻旺发、沉降,导致沉积物有机质含量升高,为钩虾提供了丰富的食物来源[39]。
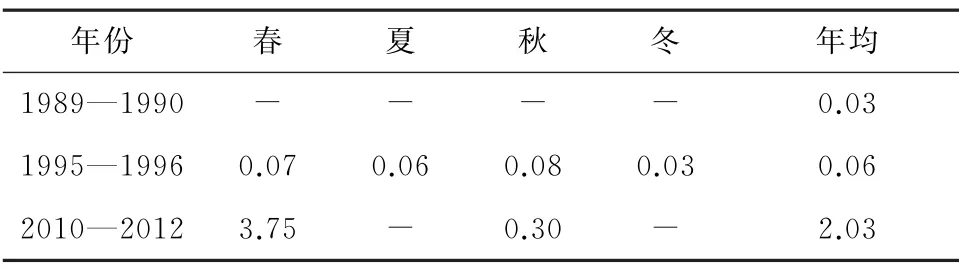
表17 大型底栖生物生物量历史变化[9-10,20,25,29] g/m2

表18 大型底栖生物密度历史变化[9-10,20,25,29] 个/m2

表19 大型底栖生物物种数历史变化[9-10,20,25,29]
2.5 潮间带生物
1985年潮间带调查的6条断面中,D 断面是岩滩,其余5个断面为软质粉砂滩[21];2010—2012年的3条断面分别为T1岩滩、T2泥滩、T3沙滩[29]。为有效比较2个年代的潮间带生物变化,将底质分为硬相(岩滩)和软相(泥滩、沙滩、粉砂滩)。2010—2012年潮间带生物平均密度和生物量均较1985年增加,软相底质与硬相底质的变化趋势相同。1985年软相底质潮间带生物的年均密度和生物量 分 别 为44.93 个/m2和19.73 g/m2,低 于2010—2012 年 的217.97 个/m2和34.61 g/m2;1985年硬相底质潮间带生物的年均密度和生物量分别为46.48 个/m2和59.93 g/m2,低于2010—2012年的95.83个/m2和444.80 g/m2。潮间带生物密度和生物量的增加主要由中潮和低潮区生物增加导致的,高潮区反而是减少的(表20)。潮间带生物数量变化趋势可能与秦山核电站建成后邻近海域潮间带中人为采捕活动的减弱有关。1985年一期工程动工,在此之前,一些较大个体潮间带生物如牡蛎、蟹等可能会被当地渔民采捕;而核电站建成后,当地渔村搬迁,周围几无渔捕活动,因此在2010—2012年采到大量大个体的近江牡蛎等,提高了潮间带生物密度和生物量。同时秦山核电产生的温排水也可能有助于潮间带生物的附着及生长,且该区域沉积物有机质含量在升高,共同促进潮间带生物密度和生物量增长[40]。

表20 潮间带生物密度和生物量历史变化[21,29]
2010—2012年的种类数较1985 年略有减少。1985年在硬相底质断面采得潮间带生物14种,软相底质断面14~38种;2010—2012年在硬相底质断面仅采得10种(含4种藻类),软相底质断面7~16种[21,29]。2个年代生物类群均以甲壳类动物和软体动物为主[21,29],多毛类较少,无明显变化。
3 结语
综上所述,由于秦山核电厂邻近海域的观测资料较少,本研究无法描述其邻近海域生态变化的具体过程,基于现有数据,参考其他电厂生态效应的研究结果,笔者分析认为,秦山核电邻近海域生态环境的主要变化是由营养盐浓度升高和沉积物有机质增加导致的。营养盐浓度升高是藻类丰度增加的最主要原因;温排水的潜在影响主要是促进暖水性藻类增殖和浮游生物小型化;沉积物有机质增加和人类采捕活动减少可能是底栖生物和潮间带生物减少的主要原因。因此,即使在水交换条件良好的海域,电厂温排水的生态影响也应长期关注。
