猪精子发生候选基因PYGO2的分离、表达和亚细胞定位研究
2021-09-27霍金龙曾日彬刘丽仙霍海龙
赵 筱,张 霞,范 俐,杨 忠,霍金龙,曾日彬,刘丽仙,霍海龙*
(1.云南农业职业技术学院,昆明 650212;2.吕梁学院生命科学系,吕梁 033001;3.云南省农业科学院粮食作物研究所,昆明 650205;4.云南生物制药有限公司,昆明 650503;5.云南农业大学动物科学技术学院,昆明 650201)
PYGO2(pygopus family PHD finger 2),也叫PYGOPUS2或PYGOPUSHomolog2,是PYGOPUS家族成员,在果蝇中被首次鉴定为Wnt信号通路中β-连环蛋白/TCF复合物的必需转录共激活因子,它作为一种DNA结合蛋白以Wnt依赖性和非依赖性途径发挥功能[1]。Wnt信号通路是一个复杂的蛋白质相互作用网络,该通路参与细胞(包括干细胞)的增殖分化过程,其功能涉及胚胎发育、组织稳态维持和癌症发生等方面,也参与骨骼发育、骨代谢、细胞分化、极性和凋亡等生理过程[2]。该通路与细胞间的旁分泌或细胞自身的自分泌通信相关,在动物中高度保守,共序列在果蝇、小鼠、大鼠、牛、羊、狗、人类中都非常类似[3]。PYGO2能与甲基化的组蛋白结合,招募RNA聚合酶Ⅱ,进而激活靶基因转录[4-5],促进组氨酸乙酰化,并调控染色质浓缩[6]。PYGO2能影响细胞的侵染和转移,能通过Wnt/β-catenin信号通路促进血管的渗透[7],在脑、眼睛、毛囊、皮肤、肺、肾、乳腺、胰腺、卵巢、食道以及生殖腺等组织中广泛表达[8]。PYGO2调节错误或靶向缺失会导致机体严重的发育障碍,甚至会引发多种疾病,包括乳腺癌[9]、神经胶质瘤[10]、卵巢癌[11]、肝癌[12]、食管癌[13]、结肠癌[14]、肺癌[15]、肥胖以及糖尿病[16]等。PYGO2能通过Wnt依赖性机制调节基因转录、细胞分化及精子发生[6, 12]。睾丸组织中的PYGO2主要在染色质重塑的长形精子细胞中表达,是促进精子成熟的正调控因子,也因此被称为染色质重塑因子,它可以通过调节组蛋白甲基化和乙酰化而参与染色质重塑过程并引发主动转录[17]。另外,PYGO2可担当CTNNB1、HNMT、TMPRSS11D(HAT)与染色质之间的支架蛋白,并在CTNNB1的核保留中起重要作用[4]。PYGO2在睾丸中以PHD依赖性方式与染色质结合,其具有染色质重塑的睾丸特异性,错误调节会导致精子在生成过程中发生障碍[17]。敲除PYGO2基因的雄性小鼠,其精蛋白的表达明显减弱,导致精子缺陷、数量减少或无精子,进而引起小鼠发育异常和不育[4]。在人上,PYGO2编码区突变会导致蛋白空间结构破坏并诱发特发性少精子症和无精子症[18],PYGO2还参与生殖细胞中不依赖于HAT的调节机制,不仅与染色质结合,而且在睾丸中参与了长形精子细胞的染色质重塑[19]。最近发现,PYGO2在雄激素依赖性和非雄激素依赖性PrCa细胞系的过度表达可能导致前列腺癌加重甚至转移[20],故PYGO2还可作为前列腺切除术后PSA进展的潜在诊断标记[21]。
近几十年来,Y染色体微缺失、囊性纤维化、精索静脉曲张、克莱氏综合征等导致精子数量减少,男性不育症成比例增加[22]。但导致不育的原因较为复杂,特发性少精子症和无精子症是多病因引发的,其具体发病机制尚未被完全阐明,因此,研究人员一直致力于寻找引发这些疾病的根本原因[23]。基于PYGO2基因在精子形成过程中的重要作用,本研究以版纳微型猪近交系(BMI)为研究材料,克隆猪PYGO2基因,并进行了一系列蛋白质生物信息学功能分析,利用qPCR和亚细胞定位确定其表达,为PYGO2在猪精子生成方面的功能研究奠定基础。
1 材料与方法
1.1 试验动物和试剂
版纳微型猪近交系10月龄公猪3头,体况良好,来自昆明原种猪场,屠宰取睾丸和其他组织样品,试验所需试剂均购自大连TaKaRa公司。
1.2 基因扩增及序列测定
根据GenBank猪PYGO2 mRNA序列(NM_001185175)设计PYGO2-clone引物(表1),以BMI睾丸cDNA为模板扩增PYGO2基因全长编码区。反应体系25 μL:2×GC Buffer 12.5 μL,2.5 mmol·L-1dNTP mix 2.5 μL,10 μmol·L-1PYGO2-clone上、下游引物各1 μL,20 ng·μL-1cDNA 1.5 μL,Ex Taq酶0.5 μL,H2O 6 μL。扩增程序:95 ℃ 5 min;95 ℃ 30 s,59 ℃ 30 s,72 ℃ 80 s,35个循环;72 ℃ 10 min。扩增产物送昆明硕擎生物公司测序。

表1 PYGO2基因引物信息
1.3 mRNA表达分析
以睾丸组织cDNA为模板制作标准曲线,利用引物PYGO2-qPCR(表1)检测PYGO2在BMI猪15种组织的mRNA表达,同时以GAPDH(表1)为内标进行校正,采用2-ΔΔCt方法进行数据分析。
1.4 亚细胞定位检测
根据上述已获得的PYGO2基因CDS序列及pEGFP-C1绿色荧光蛋白表达载体上酶切位点的分布情况,设计PYGO2-ex引物(表1)再次扩增猪PYGO2基因。将产物与pMD18T克隆载体连接,然后将重组载体pMD18T-PYGO2与pEGFP-C1载体同时双酶切(EcoRⅠ、BamHⅠ),获得真核表达重组质粒pEGFP-C1-PYGO2,转染猪睾丸ST细胞系。用红色的Mito Tracker染色ST细胞线粒体,用蓝色的Hoechst 33342染色ST细胞细胞核,显微镜下检测PYGO2蛋白在ST细胞中的表达定位情况。
1.5 生物信息学分析
利用Lasergene7.1校对测序的PYGO2序列,用ProtParam程序预测PYGO2蛋白质的分子量、分子式、等电点、正负电荷残基数;PYGO2蛋白质的二级结构、疏水结构和功能位点分别使用SOPMA、ProtScale和Prosite预测;用MEGA7.0分析7个哺乳动物PYGO2氨基酸序列同源性并构建进化树;用String进行蛋白互作网络分析。
2 结 果
2.1 PYGO2基因及氨基酸结构信息
利用PYGO2-clone引物扩增PYGO2基因的完整编码区(CDS)及部分非编码区(UTR),获得1 229 bp 长的产物(图1A)。PYGO2基因CDS序列长1 221 bp,编码406个氨基酸(图1B),含有PHD_PYGO2保守结构域(图1B,1C,1D)。PYGO2定位于猪(Sscrofa11.1)4号染色体,全长4 268 bp,Ensemble网站分析发现有4个外显子和3个内含子(图1D)。
2.2 PYGO2蛋白质序列及结构分析
猪PYGO2蛋白质分子量41.19 ku,分子式C1826H2775N507O545S20,等电点6.89,负电荷残基和正电荷残基均为26。PYGO2蛋白质406个氨基酸的二级结构中无规则卷曲占比最大,包含307个氨基酸;α螺旋次之,有57个氨基酸;延伸链28个氨基酸;β转角最少,有14个氨基酸。蛋白质的第379位氨基酸具有最大疏水值1.778,第43位氨基酸处具有最小疏水值-3.389,N端和C端均疏水。含有酶磷酸化、酰胺化等活性位点。
2.3 PYGO2多物种氨基酸序列同源性分析
7个哺乳动物PYGO2氨基酸序列比较结果发现,猪PYGO2(AVB77243.1)与牛Cattle(NP_001095712)、绵羊Sheep(XP_014962310)、小鼠Mouse(NP_001280697)、大鼠Rat(NP_001099917)、人Human(NP_612157)和黑猩猩Chimpanzee(XP_008968530)相似性分别为98.3%、97.8%、97.5%、97.3%、97.3%、97.3%,7个 物种间仅有24个不同氨基酸位点(图2A),构建的NJ树表明,牛与绵羊聚为一小支,黑猩猩与人聚为一小支,然后两者与猪聚为一大支;大鼠与小鼠聚为一支(图2B)。
2.4 PYGO2蛋白互作网络分析
蛋白互作网络分析显示,猪PYGO2与9个蛋白可能存在相互作用,包括BCL9、CTNNB1、UCK1、JUP、LEF1、CBY1、DKK1、WDR5、ENSSSCG00000015829,其中与BCL9、WDR5、CTNNB1、UCK1、JUP、LEF1相互作用较为紧密,与BCL9蛋白相互作用最为紧密(图3)。
2.5 PYGO2的多组织荧光定量表达
PYGO2 mRNA在被检测的15个组织中均有不同程度表达,在尿道球腺、前列腺、睾丸、精囊腺、附睾等生殖腺中表达相对较高,在十二指肠、结肠和肺中也有一定表达,在其余组织表达相对较低(图4)。

柱上不同大写字母表示差异显著
2.6 PYGO2亚细胞定位分析
亚细胞定位结果表明,PYGO2蛋白主要定位在猪睾丸ST细胞系的细胞核内,也有少量分布于细胞质,与PSORT网站预测的94.1%定位在细胞核一致(图5)。
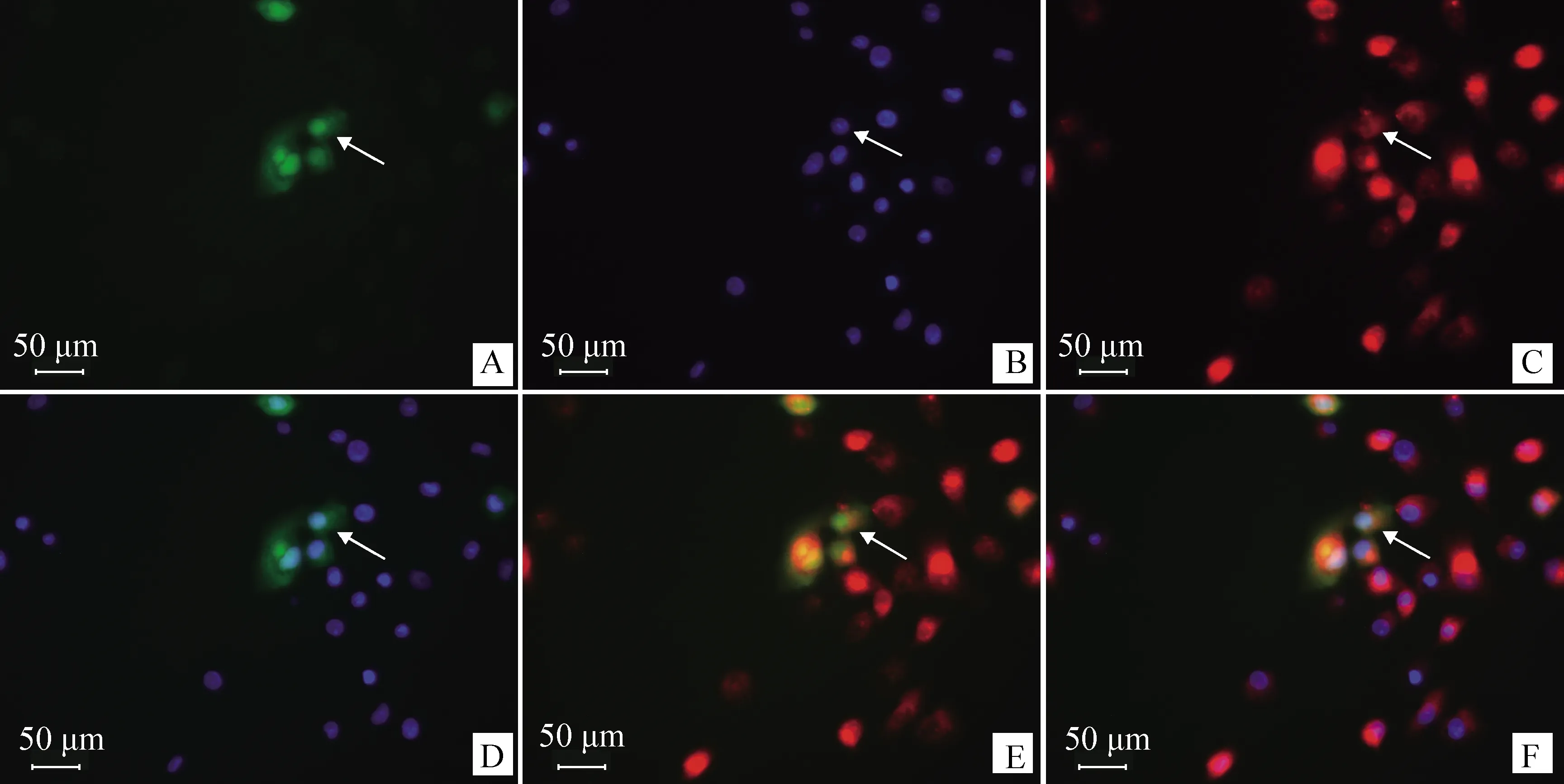
箭头所示:A.绿色荧光蛋白;B.蓝色细胞核;C.红色线粒体;D.融合的绿色荧光蛋白与细胞核;E.融合的绿色荧光蛋白与线粒体;F.融合的绿色荧光蛋白、细胞核和线粒体
3 讨 论
本研究通过BMI睾丸cDNA扩增获得了PYGO2基因序列1 229 bp(图1A),包括编码序列1 221 bp,编码406个氨基酸(图1B、1C),序列已提交GenBank,获得基因和蛋白质登录号分别为KY644518和AVB77243.1。猪PYGO蛋白C端包含PHD锌指结构域(图1B、1C、1D),在329~382位共53个氨基酸。PHD的长度一般多为50~80个氨基酸,其手指基序类似于金属结合RING域和FYVE域,以单个手指的形式出现,在物种进化中高度保守,常以两个或三个为簇,也可以和其他结构域共同参与基因表达调控[3, 24]。携带PHD结构域的蛋白质具有“读码器”功能,能将染色质重塑与基因转录的变化联系起来[25],但PYGO2在睾丸中体现的是与染色质结合的结构蛋白的功能,并非经典的转录因子功能,PYGO2-A342E突变会导致小鼠睾丸特异性缺陷,但组织形态没有明显异常,表明PYGO2在睾丸中结合的是染色质[26]。Liang等[8]在哺乳动物中发现,PHD可与组蛋白H3上被甲基化的赖氨酸结合,参与组蛋白修饰因子的募集及组蛋白甲基化,并通过Wnt通路的靶基因招募β-catenin来激活转录,去除PHD结构域后,PYGO2的转录活性降低了80%。但果蝇的PHD结构域携带了一个氨基酸突变,使其无法结合H3K4me,这可能是PHD结构域在物种进化上的一个例外[26]。

A.PYGO2基因RT-PCR产物:M.DNA相对分子质量标准;1.PCR产物。B.PYGO2基因编码序列及氨基酸序列:双下划线表示起始密码子,单下划线为保守结构域PHD_PYGO2(329~382位氨基酸),星号表示终止密码子;上一行字母为核酸序列,对应的下一行为其编码的氨基酸序列。C.PYGO2蛋白质保守结构域。D.PYGO2所处染色体位置、cDNA序列和蛋白质保守结构域
PYGO2主要在Wnt/β-连环蛋白信号传导通路中发挥重要功能,也是Wnt和Akt两个信号通路下游的共同节点,可通过翻译后修饰调节正常组织和肿瘤组织中细胞的生长和分化[27]。在猪PYGO2蛋白质也发现了一些磷酸化和酰胺化位点,这为进一步研究该基因翻译后修饰的功能奠定了基础。氨基酸序列比对发现,猪PYGO2与牛、绵羊、小鼠、大鼠、人和黑猩猩等哺乳动物序列相似度均大于97%,7个 物种间仅有24个差异氨基酸位点(图2A),表明PYGO2在物种进化上高度保守。

图2 猪和其他6个物种PYGO2氨基酸序列差异位点(A)及分子系统进化树(B)
蛋白互作网络分析发现,PYGO2与BCL9、CTNNB1、UCK1、JUP、LEF1、CBY1、DKK1、WDR5、ENSSSCG00000015829等9个蛋白存在相互作用(图3)。其中,ENSSSCG00000015829还未注释基因名字;BCL9和PYGO2的相互作用最为明显,能对Wnt/β-Catenin信号通路起正调节作用,是基因转录的关键衔接蛋白[28],PYGO2-BCL9复合物可作为结合染色质的连接因子,增强Wnt通路的特异性[29]。CTNNB1(也称为β-catenin)在Wnt-β-catenin信号传导途径中起关键作用,不仅与多种肿瘤的发生相关,也在动物生殖生理中发挥重要功能,如在山羊中,它的可变剪接与转录和生理过程相关,26 bp片段的缺失和初产仔数相关[30]。UCK1是基于KLHL2的E3连接酶的直接生理底物,能催化尿苷磷酸化为尿苷5′-单磷酸,和KLHL2形成二元复合物,处于嘧啶-核苷酸生物合成的挽救途径,在细胞凋亡中发挥重要作用,是肿瘤识别的分子标记[31]。JUP能与SOX4一起调控Wnt信号,与睾丸生殖细胞肿瘤相关[32]。LEF1是转导Wnt3a信号的主要转录因子,是癌症预后的生物标志物[33],特别在前列腺癌方面,LEF1与PYGO2一样,是引发和维持癌变的信号通路和机制中心,可以调节雄激素活性和其受体的表达,是理想的治疗靶标[34]。CBY1参与β-catenin的组成型激活,该蛋白下调能促进β-catenin在细胞核区域的滞留和转录激活,能影响干细胞未折叠蛋白的应激反应和引发细胞自噬[35]。DKK1分泌蛋白可通过诱导细胞凋亡和阻止细胞增殖来抑制Wnt/β-catenin通路,DKK1失调与阿尔茨海默病、骨形成和癌症的发病有关,也可以作为这些病症的生物标志物,DKK1同样也是癌症免疫疗法的理想靶标[36]。WDR5能靶定组蛋白H3,在H3K4三甲基化和MLL复合物的形成过程中发挥重要功能,WDR5也是胚胎干细胞多种复合物的组成部分[37]。以上8个与PYGO2互作蛋白的发现为进一步深入研究PYGO2的功能提供了方向。
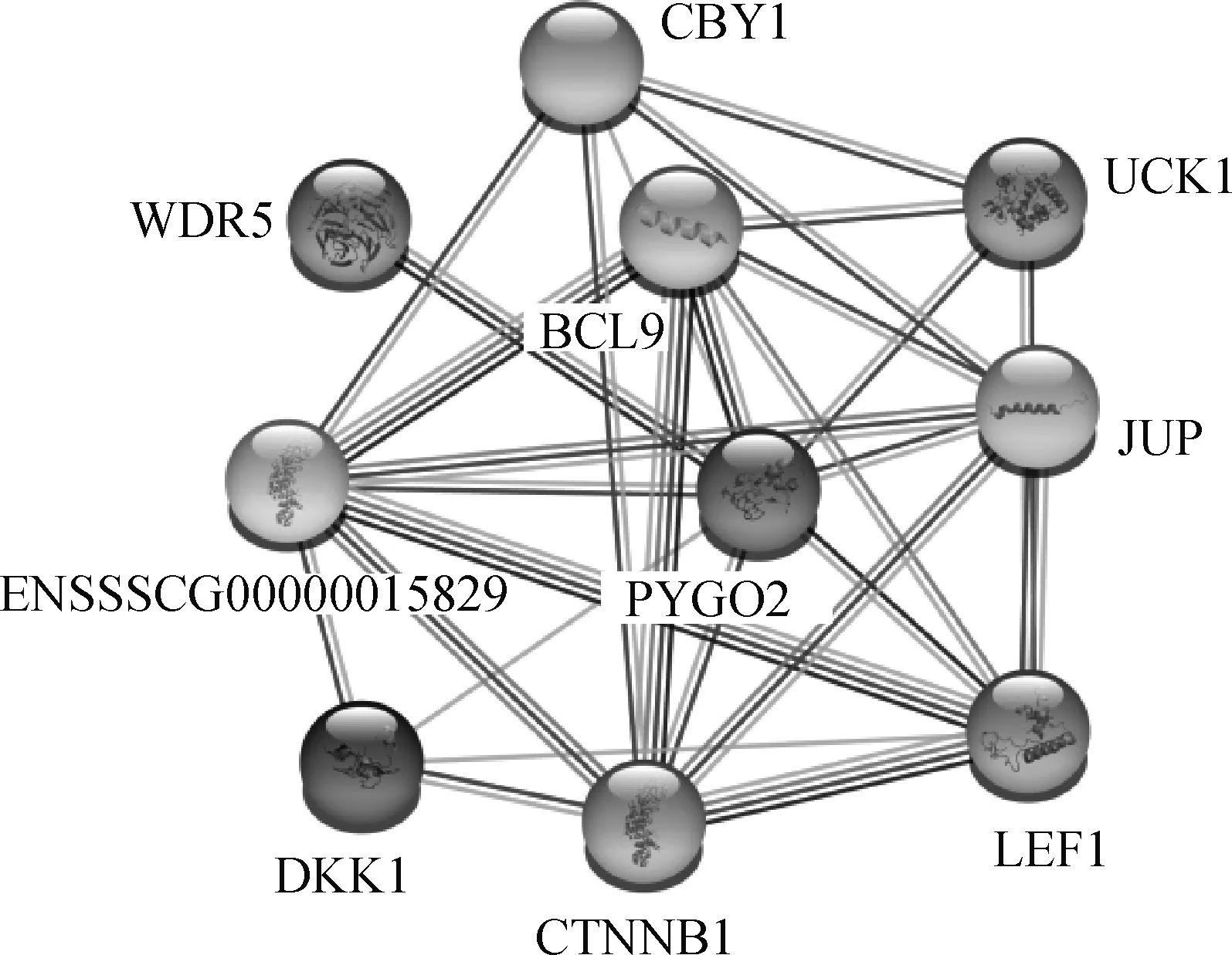
线表示蛋白质之间作用的置信度,线越多置信度越高
猪PYGO2 mRNA在被检测的15个组织中均有表达,但主要在性腺和副性腺中相对高表达,这说明PYGO2在生殖腺中发挥重要功能;在副性腺特别是尿道球腺中表达最高(图4),说明该基因在猪中也可能参与交配的性兴奋过程,并促进了一些蛋白酶、唾液酸和氨基糖类物质的分泌。小鼠中的研究表明,PYGO2蛋白可以在多种组织、多个系统中广泛表达,由于该基因参与Wnt信号,故只有在Wnt信号激活后才能维持这些组织的发育和形态发生[38]。PYGO2在睾丸中的表达主要限制在生精过程中染色质重塑发生阶段的长形精子细胞中,降低PYGO2的表达会阻滞精子发生,小鼠可以存活到成年,但是雄性生殖细胞会发生核致密化,最终导致雄性不育[4]。进一步,本研究的亚细胞定位结果显示,PYGO2蛋白主要在细胞核内表达,这与在人[38]和小鼠[26]中的研究结果一致。
4 结 论
本研究使用RT-PCR方法获得了BMIPYGO2基因的CDS序列;利用生物信息学分析方法研究了该基因的结构、蛋白质的功能和特性,构建了系统进化树及蛋白质互作网络;通过qPCR获得了PYGO2多组织mRNA表达谱;通过绿色荧光表达载体pEGFP-C1获得了亚细胞定位。本研究可为PYGO2基因在版纳微型猪近交系中的功能研究奠定基础,也可为其他哺乳动物特别是人类男性不育病理原因的深入挖掘提供基础资料。
