短尾杜鹃群落结构及多样性研究
2021-09-23龙海燕戴晓勇邓伦秀
郝 磊, 龙海燕, 戴晓勇, 邓伦秀
(1.贵州省林业科学研究院, 贵阳 550005; 2.贵州省生物研究所, 贵阳 550009)
群落是指在一定时间和空间范围内所有生物体的集合,各生物体间相互作用相互影响构成一个复杂有序的生态复合体[1]。植物群落中的物种占据着一定的生态位,具有一定的结构和功能,物种组成是群落最基本的特征,是形成群落的基础。群落结构是指群落物种在时间和空间上的配置状况,按不同的分类包括垂直结构、水平结构、种类结构、层片结构等。群落物种组成和结构特征是划分群落类型,衡量群落稳定性和预测群落动态(演替和波动)的重要依据[2-3]。多样性是指群落在物种组成、结构、功能和动态方面表现出的多样化和变异性,是物种丰富度和分布均匀性的综合反映。研究群落结构特征和多样性对了解群落现状、分析群落形成,预测群落动态变化方面具有重要意义,可为物种保护,生态恢复提供理论依据[4-5]。短尾杜鹃(RhododendronbrevicaudatumR. C. Fang et S. S. Chang)为贵州杜鹃花科(Ericaceae)杜鹃花属(Rhododendron)中的特有种类,仅在雷公山国家级自然保护区、麻江老蛇冲州级自然保护区和贞丰龙头大山保护区有分布,属狭域物种,数量较少,亟待保护。有关短尾杜鹃的研究鲜见报道,本研究以该种的群落为对象,旨在研究其群落结构特征和植物多样性。
1 研究区概况与方法
1.1 研究区概况
雷公山国家级自然保护区(以下简称雷公山保护区)位于贵州省黔东南州中部,地跨雷山、台江、剑河、榕江四县(东经108°5′~108°24′,北纬26°15′~26°32′),是我国中亚热带森林植被资源比较丰富的地区之一,主要保护对象为中亚热带森林生态系统和珍稀动植物资源,保存有最大的台湾杉天然群落;保护区总面积47 300 hm2,主峰海拔2 178.8 m,为苗岭之巅;保护区年均温14.3 ℃,年降水量1 400~1 600 mm;土壤类型包括黄壤、山地黄棕壤和山地灌丛草甸土;样地位置距离居民区约10 km,属于保护区的核心地段,原生性很强。老蛇冲州级自然保护区(以下简称老蛇冲保护区)位于贵州省黔东南州麻江县西南部(东经107°24′8″~107°17′34″、北纬26°25′23″~26°20′8″);保护区总面积8 678 hm2,平均海拔1 400 m,最高海拔1 862 m;保护区属中亚热带湿润季风气候,年均温13.9 ℃,降雨量1 400~1 530 mm;土壤母质以石灰岩、白云质砂页岩为主,土壤类型以黄壤为主,也有部分黄红壤和沙壤;主要保护对象为中亚热带常绿阔叶林生态系统;样地位置距离居民区约2 km,属于保护区的边缘地段,由于在20年前受到严重破坏,属于次生植被类型。上述两个保护区相距约140 km。
1.2 研究方法
1.2.1样地设置与调查
野外调查于2020年3月末至4月初进行,分别在雷公山保护区的雷公坪(样地 Ⅰ)和老蛇保护区的大开田(样地 Ⅱ)进行短尾杜鹃群落野外调查;调查选择有代表性的地段设置20 m×30 m的样地,并将样地划分为24个5 m×5 m的小样方;生境调查:拍照并记录样地位置、海拔、坡向、坡度、坡位、土壤类型、土层厚度;群落调查:按照小样方顺序调查记录样地内所有胸径DBH≥1 cm的个体种名、胸径、高度、冠幅;灌木层和草本层记录种名、数量、平均高度、盖度。对不能确定种名的采集标本,查阅《贵州植物志》《中国植物志》鉴定(表1)。

表1 短尾杜鹃群落样地概况
1.2.2多样性指数及重要值计算
Margalef丰富度指数(R)=S-1/lnN;
Pielou均匀度指数(Jsw)=H′/lnS;
重要值(IV)=(相对多度(DR)+相对频度(FR)+相对显著度(PR))/3;
式中:S为样地内总物种数;N为样地内总个体数;Pi为第i个物种的个体数在样地内所占的比例;a为群落A的物种数;b为群落B的物种数;c为群落A和群落B共有物种数。
1.3 数据处理
采用Excel软件进行数据统计分析,Originlab 9.1软件作图。
2 结果与分析
2.1 群落物种组成与重要值
2.1.1物种组成
2个短尾杜鹃群落共有维管束植物139种,隶属于57科95属(表2);其中群落Ⅰ有39科59属72种,物种数最多的是蔷薇科(9种),其次为杜鹃花科(8种)和百合科(4种),分别占群落总数的12.50%、11.11%、5.56%,含2~3种的科有13科(共28种),占38.89%,单种科有23科(共23种),占31.94%。群落Ⅱ有45科71属95种,物种数最多的科是杜鹃花科(9种),其次为蔷薇科(8种)、樟科(5种)和百合科(5种),分别占该群落总数的9.47%、8.42%、5.26%、5.26%,含2~4种的科有16科(共43种),占45.26%,单种科有25科(共25种),占26.32%。物种组成是群落的基本特征之一,是构成群落的基础和区分不同群落类型的重要因素。群落Ⅱ的物种丰富度比群落Ⅰ高,主要原因可能是群落Ⅱ受人为干扰频繁,次生性较强,干扰形成的生态位为更多的物种提供了生存空间;也符合中度干扰理论,即中等程度的干扰可以维持较高的物种多样性。

表2 短尾杜鹃群落维管束植物科属种组成
2.1.2重要值
2个短尾杜鹃群落各层次优势种明显(表3);群落Ⅰ乔木层重要值排名前三的依次为木荷(27.19)、亮叶桦(19.07)、短尾杜鹃(13.84),三者之和占乔木层重要值的60.10%;群落Ⅱ乔木层重要值排名前三的依次为贵州桤叶树(13.19)、美脉花楸(9.68)、木荷(8.79),三者之和占乔木层重要值的31.66%。群落Ⅰ灌木层重要值排名前三的依次为狭叶方竹(64.87)、短尾杜鹃(9.26)、木荷(3.07),三者之和占灌木层重要值的77.20%;群落Ⅱ灌木层重要值排名前三的依次为狭叶方竹(35.96)、杜鹃(6.17)、半齿柃(4.91),三者之和占灌木层重要值的47.04%。群落Ⅰ草本层重要值排名前三的依次为黑足鳞毛蕨(15.57)、十字薹草(14.47)、橐吾(6.83),三者之和占草本层重要值的36.87%;群落Ⅱ草本层重要值排名前三的依次为姬蕨(9.80)、五节芒(8.93)、浆果薹草(8.29),三者之和占草本层重要值的27.02%。2个群落草本层均不发达,对群落结构形成和生态功能发挥贡献有限。重要值是衡量某一物种在群落中的地位和作用的量化指标[6],2个群落乔木层优势现象明显但优势种种类存在差异,排名前十的物种中有4个共有种,分别是木荷、亮叶桦、短尾杜鹃和珍珠花。灌木层狭叶方竹的重要值远高于其他物种,主要原因是狭叶方竹数量多,盖度大,重要值相对也大。2个群落草本层不发达,主要原因是群落郁闭度较高,透光率低,草本植物难以生存[7]。

表3 短尾杜鹃群落主要物种重要值Table 3 Importance values of main species in R. brevicaudatum communities
2.2 区系特征(属的分布区类型)
蕨类植物分布区类型参考臧得奎[8]的划分方法,种子植物分布区类型划分参考吴征镒[9-10]的划分方法(表4)。群落Ⅰ的59个属可划分为11个分布区类型,热带亚洲分布、北温带分布、东亚分布属数量并列第一,占总属数比例均为19.23%;其次为泛热带分布13.46%和旧世界热带分布9.62%;群落Ⅱ的71个属可划分为11个分布类型,北温带分布、东亚分布属数量并列最多,占总属数比例为22.22%,其次为热带亚洲分布15.87%和泛热带分布12.70%。群落Ⅰ热带性质的属(2~7项)有26属,占总属数的50.00%,群落Ⅱ热带性质的属(2~7项)有26属,占总属数的41.27%。2个群落地理成分较为复杂,热带性质的属占优势,构成了各自群落的主体,群落物种区系特征与其所处中亚热带季风气候及地理环境相一致。
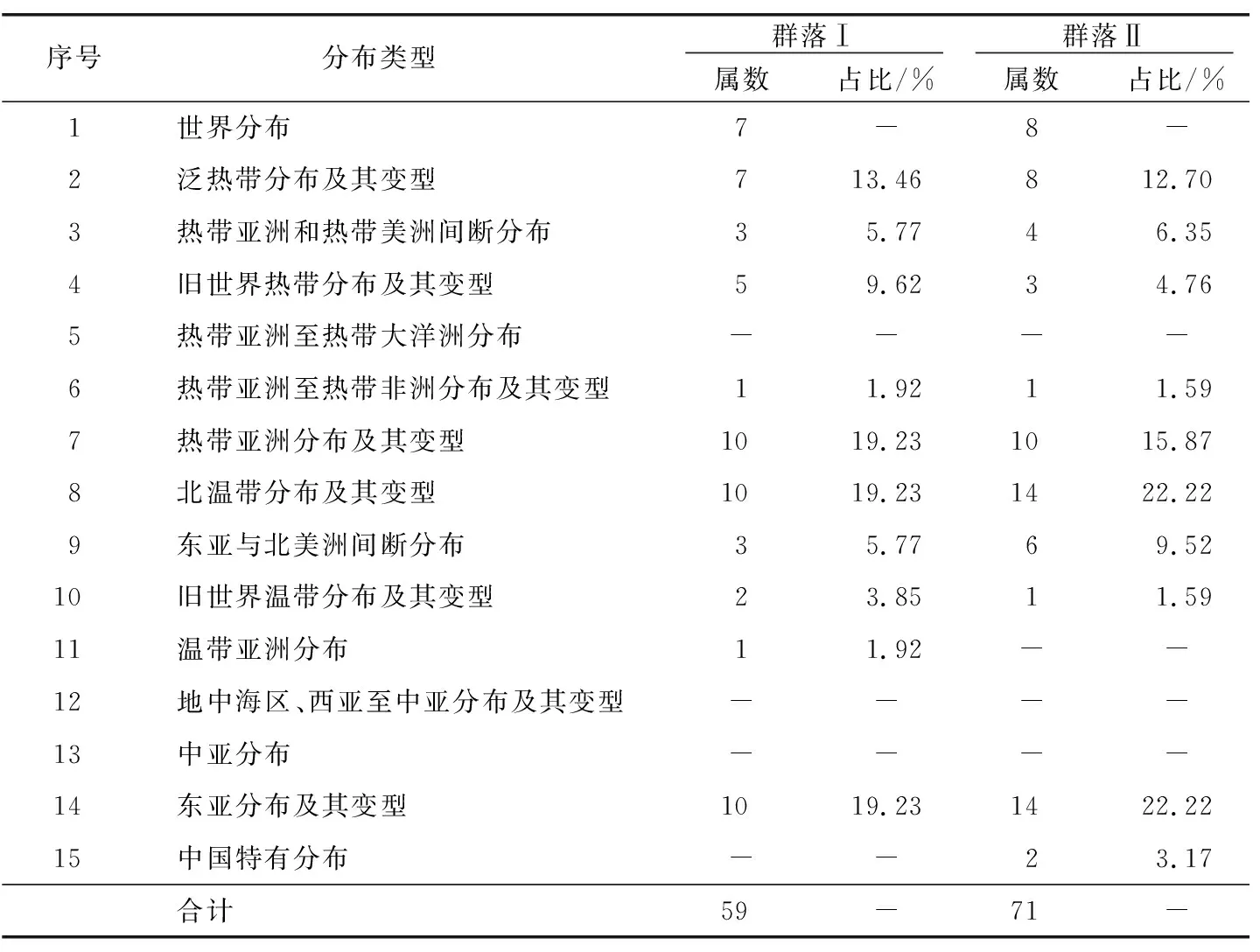
表4 短尾杜鹃群落维管束植物属的分布区类型
2.3 植物生活型谱
根据Raunkiaer生活型分类系统(图1),群落Ⅰ高位芽植物数量最多,占比66.67%,其他依次为地面芽植物(18.05%)、地上芽植物(8.33%)、地下芽植物(5.56%),一年生植物占比最低(1.39%);群落Ⅱ高位芽植物占比最多,为70.53%,其他依次为地面芽植物(11.58%)、地上芽植物(9.47%)、地下芽植物(7.37%),一年生植物最少(1.05%)。植物的生活型是植物与环境长期适应的一种形态表现,是植物对环境及其节律变化的综合反映,不同地区、不同群落类型植物生活型各不相同[11-12]。生活型谱是某一地区或某一群落内不同生活型物种构成及比例关系,可作为群落结构和外貌的表征。尽管不同群落植物生活型谱存在差异,但2个群落植物生活型均以高位芽植物为主,说明该地区气候温暖湿润,植物不需要特殊的保护便可安全越冬[13-14]。
2.4 群落结构分析
2.4.1垂直结构
垂直结构指群落在垂直方向上具有分层现象,即成层性,群落垂直结构(层次)的划分标准参照杨阳等[15]的研究。调查统计显示,2个短尾杜鹃群落成层性明显,垂直方向上可分为3个层次,即乔木层、灌木层和草本层。群落Ⅰ乔木层优势种有木荷、亮叶桦、短尾杜鹃等;灌木层优势种为狭叶方竹,以及乔木层树种的幼苗和幼树,如短尾杜鹃、木荷、珍珠花等;草本层种类和数量均较少,主要有黑足鳞毛蕨、十字薹草等;群落Ⅱ乔木层主要由贵州桤叶树、美脉花楸、木荷等组成;灌木层以狭叶方竹、杜鹃、半齿柃、短尾杜鹃为优势种;草本层不发达,以姬蕨、五节芒、浆果薹草为主。
2.4.2径级结构
1) 群落径级(DBH)结构
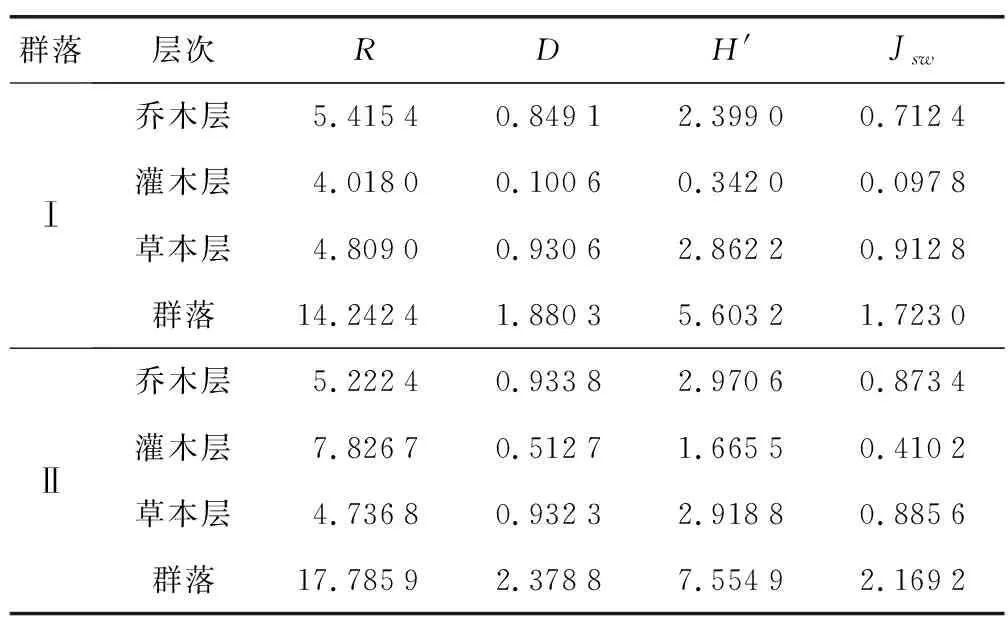
以2cm为径级单位统计径级分布(图2),群落Ⅰ DBH≥1 cm的个体共有237个,径级范围为1~43.2 cm,平均胸径6.1 cm,径级结构呈“倒J型”,个体数随径级增大而减少,中小径级个体数量多,主要集中在2 cm 2) 主要优势种径级结构 主要优势种径级结构(图3),群落Ⅰ木荷DBH≥1 cm的个体数有38个,径级范围1~43.2 cm,平均胸径10.6cm;径级结构呈“倒J型”,不连续,主要集中在1 cm 2.4.3高度级结构 1) 群落高度级(H)结构 以2 m为高度级单位统计群落高度级分布(图4)。群落Ⅰ高度级范围为1.5~20 m,平均高度5.8 m,高度级结构呈“倒J型”,个体数随高度增加而减少,主要集中在2≤H<4 m,占总数的41.35%。群落Ⅱ高度级范围为1.5~20 m,平均高4.1 m,高度级结构呈“单峰型”,主要集中在2≤H<6 m,占总数的76.32%。群落高度级结构一定程度上可以反映群落的现状、外貌以及受外部环境的影响程度。群落Ⅰ的平均高度高于群落Ⅱ,说明群落Ⅰ受外界环境干扰较少,群落相对稳定,演替逐步向顶级群落发展,群落Ⅱ则因为频繁的干扰演替进程缓慢。 2) 主要优势种高度级结构主要优势种高度级结构(图5),群落Ⅰ木荷高度级范围为1.5~20 m,平均高度8.1 m;高度级结构呈“倒J型”,主要集中在2≤H<4 m,占总数的28.95%;亮叶桦高度级范围2.5~20 m,平均高度9.7 m,高度级结构呈“单峰型”,个体主要集中在6≤H<14 m,占总数的72.50%%;短尾杜鹃高度级范围为1.5~5 m,平均高度3.1 m,高度级结构呈“单峰型”,主要集中在2≤H<4 m,占总数的71.64%。群落Ⅱ贵州桤叶树高度级范围为2~7.5 m,平均高度4.7 m;高度级结构呈“单峰型”,主要集中在4≤H<6 m,占总数的50.00%;美脉花楸高度级范围为5~8 m,平均高度6.3 m,高度级结构呈“单峰型”,个体主要集中在6≤H<8 m,占总数的62.50%。木荷高度级范围为4~10 m,平均高度6.1 m,主要集中在6≤H<8 m,占总数的56.52%。群落高度级结构与群落物种的生长型、群落发展阶段和群落环境密切相关[16]。2个群落主要优势种高度级结构存在差异,群落Ⅰ优势种高度级结构总体大于群落Ⅱ;共有树种木荷平均高度群落Ⅰ大于群落Ⅱ,说明2个群落演替阶段不同,受环境影响程度也不一样。 2.5.1群落相似性 根据Jaccard相似性指数,2个短尾杜鹃群落有共有属35个,共有种28个,两者共有属和共有种相似系数分别为0.368 4、0.201 4。2个群落处于相同的气候区内,气候、地形、海拔等环境因素相似,因此,群落物种具有一定程度的相似性。 2.5.2多样性指数 各多样性指数在不同群落和同一群落不同层次间的变化趋势存在差异(表5)。群落Ⅱ的Margalef丰富度指数、Simpson优势度指数、Shannon-Wiener多样性指数和Pielou均匀度指数均高于群落Ⅰ,说明群落Ⅱ整体物种种类更多,各物种多度差异不如群落Ⅰ大,分布相对更均匀。除群落ⅡMargalef丰富度指数外,2个群落灌木层各多样性指数均低于乔木层和草本层。主要原因是灌木层狭叶方竹数量占绝对优势,群落Ⅰ和群落Ⅱ分别有狭叶方竹2 727株和1 011株,占各自灌木层总个体数的94.82%和69.48%,分布极不均匀。群落Ⅱ灌木层有维管束植物58种,远高于乔木层30种和草本层27种,Margalef丰富度指数较高。 表5 短尾杜鹃群落各层次多样性指数 3.1.1物种组成及重要值 植物群落存在地域分异,不同群落物种组成千差万别,物种组成是群落最基本的特征之一,是形成群落结构的基础和区分不同群落类型的因素[16-17]。群落Ⅱ的物种种类高于群落Ⅰ,一方面是因为2个群落所处的演替阶段不同,另一方面群落Ⅱ受人为干扰频繁,干扰提高了空间异质性,使更多物种定居成为可能,也符合中度干扰维持较高的物种多样性理论[18]。2个群落乔木层重要值较高的优势种不尽相同,排名前十的物种中有共有种4个,分别是木荷、亮叶桦、短尾杜鹃、珍珠花;灌木层狭叶方竹重要值最高,其次为短尾杜鹃、杜鹃、半齿柃;草本层不发达,种类和数量较少,重要值最高的为黑足鳞毛蕨,其次为十字薹草、姬蕨、浆果薹草等。重要值是衡量某一物种在群落中地位和作用的量化指标。乔木层优势种种类和重要值存在差异主要是因为2个群落处于不同的演替阶段,所受干扰程度不一,导致构成群落的物种及其作用大小不同;灌木层狭叶方竹在2个群落中的重要值远高于其他物种,主要原因是狭叶方竹数量占绝对优势,相对多度和相对盖度大;2个群落草本层种类和数量较少,主要原因是乔木层和灌木层发达,群落郁闭度较高,草本植物生长受到抑制[19]。 3.1.2区系特征 群落Ⅰ热带性质的属(2~7项)有26属,占总属数的50.00%,群落Ⅱ热带性质的属(2~7项)有26属,占总属数的41.27%。2个群落热带性质的属均占优势,与其所处中亚热带季风性气候及地理环境相一致[21],本研究与谭维政等[22]研究结果相似。 3.1.3生活型谱 群落Ⅰ高位芽植物数量最多(66.67%),群落Ⅱ高位芽植物占绝对优势(70.53%),2个群落植物生活型均以高位芽为主,说明该地区水热条件较好,植物生长周期长。生活型是生物体长期处于特定的环境条件下,通过适应性进化表现出的相应的结构和外貌,不同物种间可通过趋同适应形成相同的生活型。因此,相同或相似的生境条件下生物体具有相同或相似的生活型构成,群落结构和外貌也表现出相似性[23-24]。 3.1.4群落结构 群落结构是构成群落的物种在空间和时间上的配置状况[23]。2个群落垂直方向上可分为乔木层、灌木层和草本层,径级和高度级结构呈“倒J型”或“单峰型”,中幼个体数量多,成熟个体数量少,群落总体更新状况良好,本研究与孙越等[26]、温韩东等[27]的研究结果相似。2个群落优势现象明显,主要优势种径级结构和高度级结构呈“倒J型”或“单峰型”,种群为增长型或稳定型,无衰退型,研究结果与匡旭等相似[26]。植物径级及高度与其所在群落现状和环境密切相关,如演替阶段,人为干扰等。本研究结果表明,2个群落处于不同的演替阶段,群落Ⅰ处于演替后期阶段,向顶级群落发展,群落Ⅱ处于演替中期阶段。随着演替的进行,群落中一些先锋树种如亮叶桦可能会被逐步淘汰,而木荷、贵州青冈、巴东栎等将成为顶级群落树种。 3.1.5植物多样性 物种多样性是生物多样性的重要组成部分,是评价生态系统结构复杂性和功能多样性的重要指标之一[27],多样性是物种丰富度和分布均匀性的综合反映[28]。Margalef丰富度指数(R)可以直接反映群落物种种类的多少,群落物种种类越多,R值越大;Shannon-wiener多样性指数(H′)是群落物种种类多样性和分布均匀性的综合体现,H′值越大,表明物种数量越多,分布越均匀;Pielou均匀度指数(Jsw)是用以表征物种分布情况的,Jsw值越大,表明各物种多度差异越小,分布越均匀;Simpson优势度指数(D)用以衡量群落物种优势地位,D值越高,说明群落优势现象越不明显,多样性越高。本研究结果显示,植物多样性指数在不同群落和同一群落不同层次间的变化趋势存在差异。群落Ⅱ的R、D、H′和Jsw高于群落Ⅰ。除群落ⅡR外,灌木层各多样性指数均低于乔木层和草本层。群落Ⅱ各多样性指数高于群落Ⅰ,说明群落Ⅱ物种种类更丰富,各物种多度差异相对较小,分布更均匀。除群落ⅡR外,2个群落灌木层多样性指数均低于乔木层和草本层。主要原因是灌木层狭叶方竹数量占绝对优势,其他物种占比较低,分布不均匀所致。R可以直观的反映群落或某一层次物种数目的多寡,群落Ⅱ灌木层有维管束植物58种,远高于乔木层30种和草本层27种。 1) 2个群落共记录到维管束植物57科95属139种,其中群落Ⅰ有39科59属72种,群落Ⅱ有45科71属95种。2个群落物种重要值最大的是灌木层的狭叶方竹,群落Ⅰ和群落Ⅱ乔木层重要值排名前三的种有木荷、亮叶桦、短尾杜鹃、贵州桤叶树、美脉花楸。 2) 2个群落在属的水平上可划分为11个分布区类型,地理成分比较复杂,均以热带成分占优势,群落Ⅰ和群落Ⅱ分别占总属数的50.00%和41.27%;生活型谱均以高位芽植物为主,群落Ⅰ和群落Ⅱ分别占66.67%和70.53%。 3) 群落成层性明显,垂直方向上可分为乔木层、灌木层和草本层。群落总体径级结构和高度级结构呈“倒J型”或“单峰型”,更新良好。主要优势种径级结构和高度级结构呈“倒J型”或“单峰型”,种群为增长型或稳定型。 4) 2个群落属和种的相似性系数分别为0.368 4、0.201 4,群落Ⅱ的R、D、H′和Jsw高于群落Ⅰ。灌木层狭叶方竹占绝对优势,分布极不均匀,除群落Ⅱ的R外,灌木层各多样性指数均低于乔木层和草本层。 5) 群落Ⅰ人为活动较少,演替时间更长,处于演替后期阶段,向顶级群落发展,群落Ⅱ受人为干扰频繁,处于演替中期阶段。2.5 多样性分析
3 讨论与结论
3.1 讨 论
3.2 结 论
