高寒草地类型和海拔对濒危藏药独一味生长繁殖特征的影响
2021-09-13谢田朋崔治家张建旭张文广
谢田朋 崔治家 张建旭 张文广
摘 要:为探讨高寒草地类型和海拔对独一味(Lamiophlomis rotata)生长与繁殖特征的影响,在玛曲三种草地类型和三个海拔梯度下进行随机样方调查和样品收集,并对独一味的生长和繁殖特征指标进行测定。结果表明:(1)不同草地类型中,独一味地上部特征表现为高寒沼泽化草甸>高寒山坡草甸>高寒灌丛草甸,地下部特征表现为高寒沼泽化草甸>高寒灌丛草甸>高寒山坡草甸;(2)独一味生长繁殖特征随着海拔的升高而下降;(3)不同草地类型和海拔下,独一味有性繁殖结构的投入与植株大小间存在显著的正相关性,而无性繁殖投入与植株大小间不存在相关性;(4)不同草地类型和海拔下,独一味两种繁殖方式间不存在相关性。综上结果表明:独一味资源分配方式受到草地类型影响,是与环境长期适应后的结果;有性繁殖的发生需要植株一定量的营养生长积累,而无性繁殖投入可能是植株固有特性,与植株大小无关;根茎芽无性繁殖可能发生在地上部破坏之后而非主动行为,两种繁殖方式间的无相关性是否会受到除草地类型和海拔以外的其他因素影响还有待进一步研究。
关键词:药用植物,高寒草地类型,海拔,生长与繁殖特征,根茎芽
中图分类号:Q945.5
文献标识码:A
文章编号:1000-3142(2021)08-1346-08
Abstract: In order to explore the influence of alpine grassland types and altitudes on growth and reproduction traits of Lamiophlomis rotata, random quadrat investigation and sample collection were conducted under three grassland types and three altitude gradients in Maqu, and the growth and reproduction traits were measured. The results were as follows: (1) The traits of aboveground parts were alpine swamp meadow > alpine hillside meadow > alpine shrub meadow, and the traits of the underground parts were alpine swamp meadow > alpine shrub meadow > alpine hillside meadow; (2) The traits were decreased with the increase of altitude; (3) In different grassland types and altitudes, there was a significant positive correlation between the sexual reproduction structure and plant size, but there was no correlation between the asexual reproduction and plant size; (4) In different grassland types and altitudes, there was no correlation between the two reproduction modes. All the above results indicate that the resource allocation method is affected by grassland types, which is the result of long-term adaptation to the environment; The occurrence of sexual reproduction requires a certain amount of vegetative growth accumulation, while the asexual reproduction may be the inherent characteristics of plants, independent of plant size; The asexual propagation of rhizome bud may occur after the destruction of the aboveground rather than the active behavior. Whether the uncorrelation between the two propagation modes would be affected by other factors needs further study.
Key words: medicinal plant, alpine grassland types, altitude, growth and reproduction traits, rhizome bud
獨一味(Lamiophlomis rotata)为唇形科(Lamiaceae)多年生草本植物,原属糙苏属(Phlomis L.),现已从该属中分离成为独一味属(Lamiophlomis Kudo)的唯一物种。独一味花序高2.5~10 cm,直根系,叶片4~6片,辐状两两相对,贴地而生。在我国主要分布于西藏、青海玉树果洛地区、甘肃甘南、四川西部,零星分布于云南西北部;在国外的尼泊尔、不丹和锡金也有分布(祝聪等,2018)。独一味是一种重要的民族药资源,该药材先后记载于藏医著作《月王药诊》《四部医典》《晶珠本草》中,迄今已有1 200年的历史,独一味药材表面枯黄色或黄褐色,质坚硬、干枯、气腥臭,性甘、苦、平,归肝经;具活血止血、祛风止痛之功效,用于跌打损伤、外伤出血、风湿痹痛以及黄水病等(中华人民共和国药典,2015)。目前,全国仅甘肃独一味制药企业对独一味药材的年需求量就达1 000 t以上。市场对独一味巨大的药用需求和其有限的资源现状之间的矛盾,使其在2000 年就已被列为一级濒危藏药品种(李隆云等,2002)。
现有文献表明,独一味因为受到花序结构、花柱型态、花大小、种子休眠的限制,导致有性繁殖成功率很低(Jin et al., 2011;金兰等,2016),学者们通过打破种子休眠(Jin et al., 2011)、促进花粉萌发(金兰等,2012)、控制光照(张亚娟等,2007)等方式提高其有性繁殖成功率,或通过增加幼苗抗性(蔡子平等,2009)和与其他农作物间作(何淑玲等,2012)等方式提高后代成活率。但目前独一味药材主要来源依然是野生品种,在长期的自然选择下该物种并没有灭绝,这意味着独一味必然会通过其他途径来弥补有性繁殖的不足。
尽管有学者认为独一味不存在克隆繁殖现象(刘继梅,2006),但更多研究发现,独一味存在的根茎芽可供翌年发育(孙辉等,2012;金兰,2016),因此,独一味的繁殖体系中无性繁殖和有性繁殖是共同存在的。已有文献表明,独一味根茎芽的出现不是显而易见的现象,因此,根茎芽在什么环境条件下产生、与植株大小间的关联、与有性繁殖间的关系都成为了值得深究的问题。了解独一味对野外环境的适应性和不同繁殖方式与环境间的关系,对今后人工引种栽培具有指导意义。
本文以野生独一味为研究对象,通过野外随机样方调查和样品取样分析,具体探讨以下问题:(1)草地类型和海拔因素是否对独一味的生长与繁殖特征有影响;(2)草地类型和海拔因素下其繁殖特征与植株大小间的关系如何;(3)草地类型和海拔因素下其两种繁殖特征间的关系如何。
1 材料与方法
1.1 实验地概况
青藏高原高寒草甸分布区域广阔,区域气候与人类活动干扰差异较大。该实验地点位于甘肃省甘南藏族自治州玛曲县(102°4′12″ E、33°59′56″ N)附近,该地区地处青藏高原东北缘、黄河第一弯。其东南部与四川省比邻,西南部和西部与青海省接壤。地势西高东低,地形复杂,属典型的高寒湿润的高原气候,地表开阔,多风,每年八级以上的大风有50多天;年平均气温为1.2 ℃,年最冷月(一月)平均温度为-10 ℃,极端最低温度为-29.6 ℃,年最热月(七月)平均温度为11 ℃,极端最高温度为23.5 ℃;年均降水量为620 mm;属典型高寒草甸植被类型(吴征镒,1980)。
1.2 实验方法
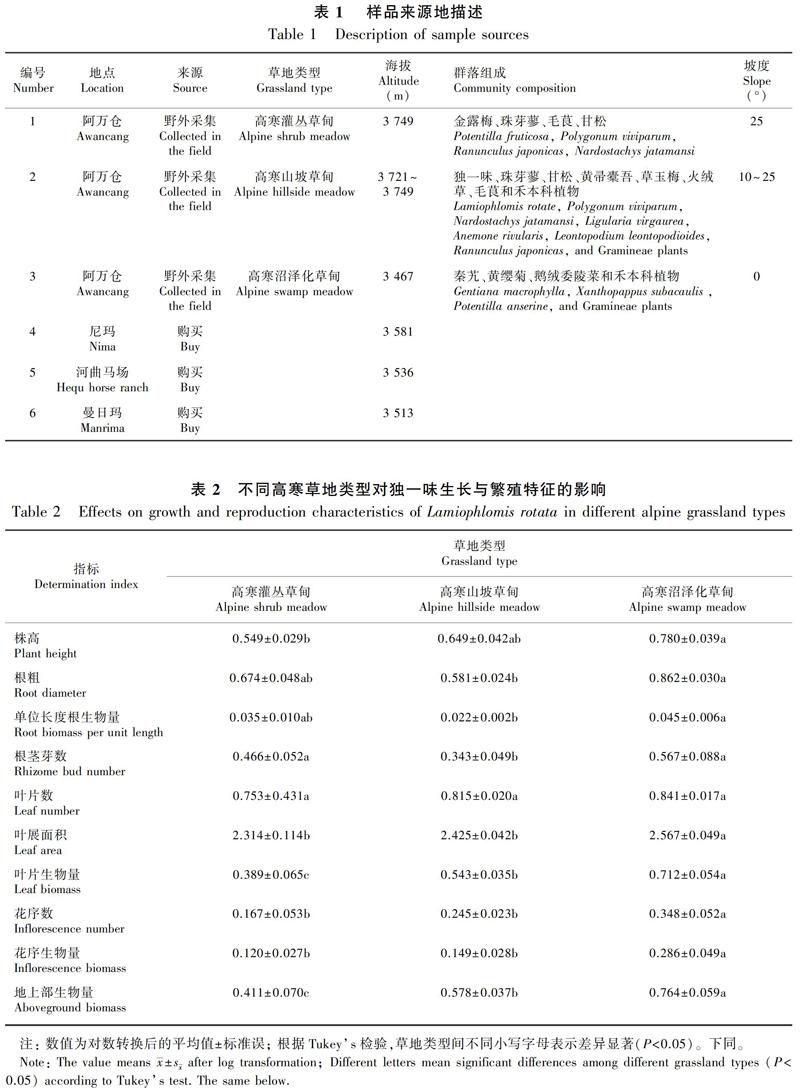
该实验于2019年7月在玛曲县附近对不同草地类型和海拔下生长的独一味进行5 m × 5 m的随机样方调查和样品采集,每个草地类型进行1~3次重复,海拔用手持GPS导航仪测定,另有编号4~6的地点样品购自当地药材收购商处(表1)。样品经兰州大学杜国祯教授鉴定为独一味。统计每个样方内独一味个体数及开花个体数,并随机挖取完整个体带回实验室,对采集样品和收购样品进行株高、根长、根粗、横叶展距、纵叶展距、叶片数、花序数、根茎芽数等指标的测定;样品经过烘箱80 ℃干燥24 h后用1/1 000电子天平测定地上部和地下部生物量。由于收购样品为牧民采挖,地下部结构不完整,不能用于评估根茎芽相关指标。在采挖过程中样品根部易断,因而用单位长度根生物量(g·cm-2)表示地下部生物量分配情况。叶展面积(cm2)=横叶展距×纵叶展距,用于估算叶面积情况。
1.3 统计方法
采用SPSS 20.0软件(SPSS Inc.,Chicago, IL)进行数据分析,为保证检测数据的方差齐性,在分析前对数据进行对数转换,转换后数据Levenes test 均为P>0.05。探讨高寒草地类型和海拔对独一味生长与繁殖特征的影响,采用一般线性模型(GLM)的one-way ANOVA,分析高寒草地类型和海拔对独一味的生长与繁殖相关指标的影响。不同因素间的平均值差异利用Tukeys 检测法。为检测高寒草地类型和海拔对个体大小与无性结构或有性结构之间关系的影响,采用协方差分析(ANCOVA),其中高寒草地類型和海拔作为固定因子,个体大小作为协变量。采用偏相关分析,通过基株大小的控制,检测有性繁殖与克隆繁殖之间的关系。
2 结果与分析
2.1 高寒草地类型对独一味生长与繁殖特征的影响
调查发现,独一味生长群落常由金露梅(Potentilla fruticosa)、珠芽蓼(Polygonum viviparum)、毛茛(Ranunculus japonicus)、甘松(Nardostachys jatamansi)、黄帚橐吾(Ligularia virgaurea)、草玉梅(Anemone rivularis)、火绒草(Leontopodium leontopodioides)、秦艽(Gentiana macrophylla)、黄缨菊(Xanthopappus subacaulis)、鹅绒委陵菜(Potentilla anserina)、禾本科(Gramineae)植物等组成(表1)。
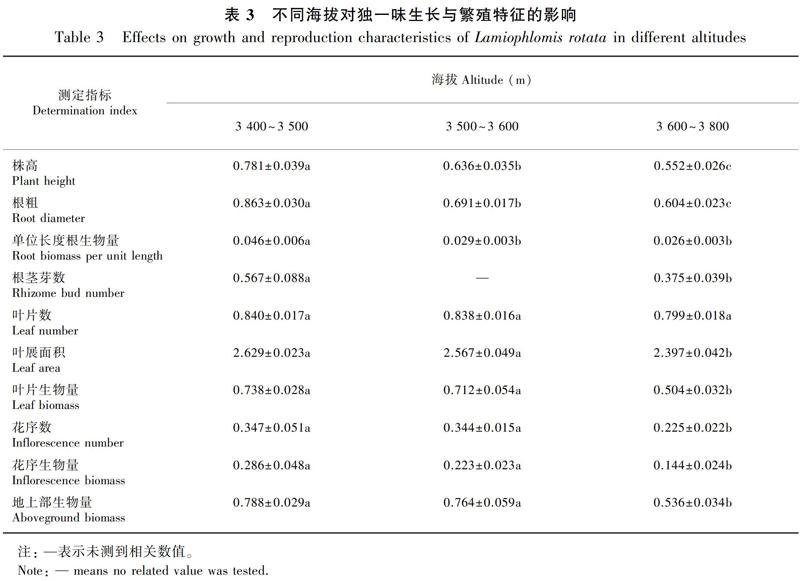
高寒草地类型对独一味的株高(F2,50=3.739,P=0.034)、根粗(F2,50=22.731,P<0.000 1)、单位长度根生物量(F2,50=6.194,P=0.004)、根茎芽数(F2,50=3.403,P=0.042)、叶展面积(F2,50=3.344,P=0.044)、叶片生物量(F2,50=8.417,P=0.001)、花序数(F2,50=4.252,P=0.020)、花序生物量(F2,50=4.295,P=0.023)、地上部生物量(F2,50=8.641,P=0.001)均有显著影响,仅对叶片数(F2,50=2.120,P>0.05)无显著影响。独一味的株高、叶展面积、叶生物量、花序数、花序生物量、地上部生物量均表现为高寒沼泽化草甸>高寒山坡草甸>高寒灌丛草甸;根粗、单位长度根生物量、根茎芽数表现为高寒沼泽化草甸>高寒灌丛草甸>高寒山坡草甸(表2)。
2.2 海拔对独一味生长与繁殖特征的影响
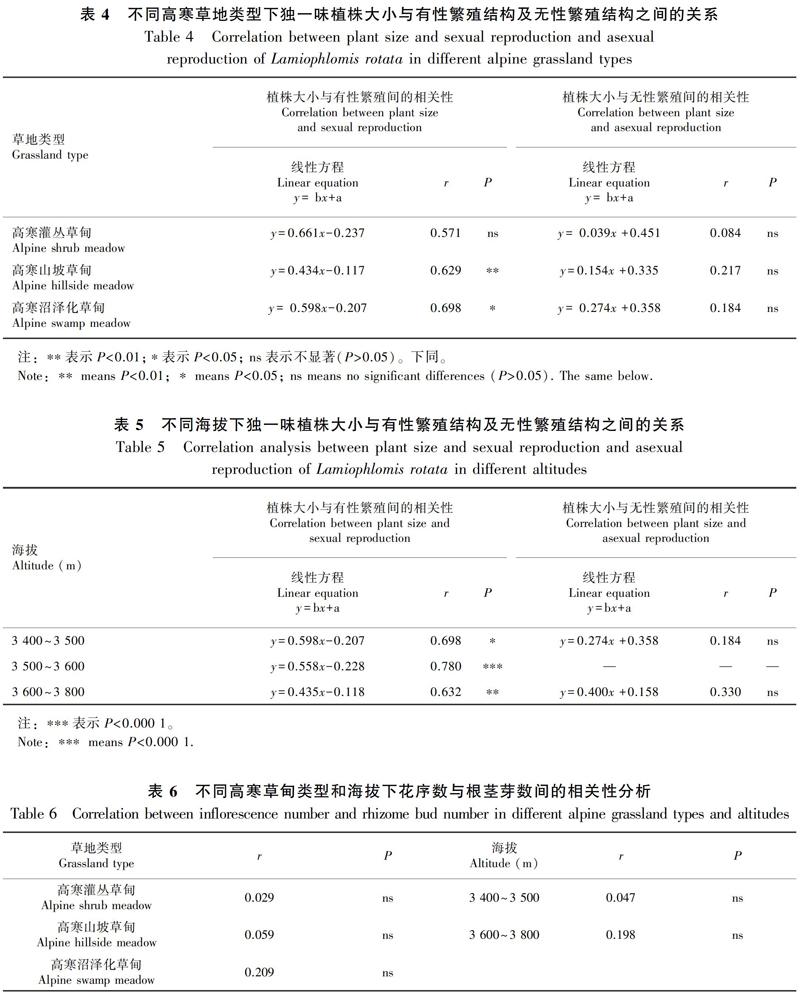
海拔对独一味的株高(F2,87=8.266,P=0.001)、根粗(F2,87=24.085,P<0.000 1)、单位长度根生物量(F2,87=5.333,P=0.008)、根茎芽数(F1,50=5.284,P=0.026)、叶展面积(F2,87=12.647,P<0.000 1)、叶片生物量(F2,87=15.053,P<0.000 1)、花序数(F2,87=28.914,P<0.000 1)、花序生物量(F2,87=5.037,P=0.010)、 地上部生物量(F2,87=15.500,P<0.000 1)均有显著影响,仅对叶片数(F2,87=1.583,P>0.05)无显著影响。独一味的株高、根粗、单位长度根生物量、根茎芽数、叶展面积、叶片生物量、花序数、花序生物量、地上部生物量均表现出随着海拔范围升高而下降的趋势(表3)。
2.3 植株大小与有性繁殖及无性繁殖间的关系
在高寒山坡草甸(P=0.005)和高寒沼泽化草甸(P=0.017)中,独一味植株大小与有性繁殖结构间存在显著的正相关性;在高寒灌丛草甸(P=0.429)中存在正相关趋势;而在任何草地类型中,植株大小与无性繁殖间均不存在相关性(P>0.05)(表4)。
在不同海拔下,独一味植株大小与有性繁殖结构间均存在显著的正相关性(3 400~3 500 m:P=0.017;3 500~3 600 m:P<0.000 1;3 600~3 800 m:P=0.002);而植株大小与无性繁殖间均不存在显著的相关性(P>0.05)(表5)。
2.4 有性繁殖和无性繁殖间的关系
不同高寒草地类型和海拔下,花序数与根茎芽数之间均不存在相关性(P>0.05),说明在独一味中两种繁殖方式间即不存在竞争,也不存在协同增长(表6)。
3 讨论与结论
3.1 生长繁殖特征对草地类型和海拔的适应性
独一味的生长繁殖特征在不同草地类型中表现不一致的现象说明其为了适应环境在资源分配上存在多种策略,独一味贴地生长的习性导致其在灌丛中无法竞争到足够的光源和地上空间,转而将更多资源投入到地下部组织中,增加了无性繁殖投入。而没有灌丛的情况下,独一味倾向于投入更多资源到地上部组织。这种在较为严苛的环境下偏向无性繁殖的策略与前人研究结果吻合(王洪义等,2005)。沼泽化草甸因群落中优势种少、水源充足等因素,更适合独一味种群生长,该研究结论与孙辉等(2012)对独一味野生资源状况实地调查结果基本吻合。
随着海拔高度的增加,空气密度、气压、温度和饱和蒸汽压等均有所降低,紫外线辐射增加,日变化大,这些不稳定的非生物因子对植物的胁迫增强(马文梅等,2019),因此限制了独一味的生长和繁殖,植物在获得的资源总量下降情况下,分配到各个器官的资源量也相应减少。这一结论与川西风毛菊(Saussurea dzeurensis)、羌塘雪兔子(S. wellbyi)的研究一致(王一峰等,2015;马文梅等,2019)。
3.2 植株大小与繁殖投入间的关系
独一味有性繁殖结构的投入与植株大小间显著的正相关性和无性繁殖结构的投入与植株大小间的无相关性在不同草地类型和海拔范围中均有所呈现。独一味的根茎芽作为有性繁殖的补偿,保证了该物种在恶劣的环境中即使没有种子繁殖也能使种群继续维持,这一机制很可能是植株本身的内在特性,与有性繁殖在发生前需要一定量的营养积累截然不同。类似结果在同样存在两种繁殖方式的高寒草甸杂草黄帚橐吾中也有报道(Xie et al., 2014)。Schmid et al.(1995)认为,对于多年生有无性繁殖习性的植物来说,无性繁殖投入相当于营养生长的一部分,因此,如果它们只是个体生长过程的一部分,就没有必要对植株大小有阈值要求。而有性繁殖投入是植株生活史周期最后一个可塑性结构,植株在积累了足够的资源之前是不可能错误地产生有性结构。
3.3 有性繁殖與无性繁殖间的关系
拥有两种繁殖方式的多年生植物,其繁殖方式间或存在此消彼长的竞争关系(Van Drunen & Dorken, 2012),或存在协同增长的正相关关系(Xie et al., 2014)。但该研究发现,独一味两种繁殖方式间在不同草地类型和海拔下均不存在相关性,这种现象在意大利海芋(Arum italicum)、黑麦草(Lolium perenne)中也有报道(Méndez,1999; Thiele et al., 2008),这可能与独一味根茎芽的特性有关,独一味根茎芽很可能在地上部全部枯萎或被采集破坏后,才能收到植株内的传导信号,随后被激活并开始补偿性生长,与植物被刈割后的反应类似(马银山等,2010),这与其他主动进行无性繁殖的植物有很大差别。这种无相关性的特征是否会受其他非生物因子(如温度、光照、水分、养分等)影响还有待进一步研究。
参考文献:
CAI ZP, CHEN Y, GUO FX, et al., 2009. Effects of exogenous Ca2+ on resistance to rapid cold and freeze of Tebet Lamiophlomis rotata(Benth.)Kudo seedlings [J]. J Desert Res, 29(5): 928-932. [蔡子平, 陈垣, 郭凤霞, 等, 2009. 外源Ca2+对藏药独一味幼苗速冷冻抗性的效应[J]. 中国沙漠, 29(5): 928-932.]
National Pharmacopoeia Commission, 2005. Pharmacopoeia of the Peoples Republic of China: Part one [M]. Beijing: China Medical Science Press, 1: 184. [国家药典委员会, 2005. 中华人民共和国药典: 一部[S]. 北京: 中国医药科技出版社,1: 184.]
HE SL, WANG GH, DONG LJ, et al., 2012. Effects of Lamio phlomis intercropping cultivation with Inula helenium I. on growth of Lamiophlomis [J]. Guangdong Agric Sci, 39(9): 20-21. [何淑玲, 王改花, 董利俊, 等, 2012. 甘南高原獨一味间作栽培藏木香对独一味生长的影响[J]. 广东农业科学, 39(9): 20-21.]
JIN L, LUO GH, DING L, 2011. Optimization of seeds germination conditions of Lamiophlomis rotata(Benth.) Kudo from Qinghai Province [J]. Med Plant, 2(3): 7-9.
JIN L, QIAO F, DING L, et al., 2016. The study of flowering characteristics and pollination efficiency for Lamiophlomis rotata of Qinghai Yushu [J]. Seeds, 35(10):41-43. [金兰, 乔枫, 丁莉, 等, 2016. 青海玉树独一味开花特性与传粉方式对传粉效率影响研究[J]. 种子, 35(10): 41-43. ]
JIN L, 2016. Preliminary study on breeding system of Qinghai Tibetan Plateau Lamiophlomis rorata [J]. N Hortic, (8): 145-149. [金兰, 2016. 青藏高原特有植物独一味繁育系统初步研究[J]. 北方园艺, (8): 145-149.]
JIN L, DU L, HAN HP, et al., 2012. Effects of GA3 on external pollen germination of transplanting Lamiophlomis rotata [J]. Seeds, 31(3):101-103. [金兰, 杜雷, 韩鸿萍, 等, 2012. 赤霉素对移栽独一味花粉体外萌发的影响研究[J]. 种子, 31(3): 101-103. ]
LI LY, ZHAN D, WEI YF, et al., 2002. Conservation of endangered species resources of Tibetan medicine in China [J]. Chin J Chin Mat Med, 27(8): 562-564. [李隆云, 占堆, 卫莹芳, 等, 2002. 濒危藏药资源的保护[J]. 中国中药杂志, 27(8): 562-564. ]
LIU JM, 2006. Genetic diversity andlipophilic composition in Lamiophlomis rotata, an endemic species of Qinghai-Tibetan plateau [D]. Shanghai: Fudan University: 93. [刘继梅, 2006. 青藏高原特有植物独一味的遗传多样性及其脂溶性化学成分分析[D]. 上海: 复旦大学: 93.]
MA YS, DU GZ, ZHANG ST, 2010. The impacts of fertilization and clipping on compensatory growth of Poa crymophilla [J]. Acta Ecol Sin, 30(2): 279-287. [马银山, 杜国祯, 张世挺, 2010. 施肥和刈割对冷地早熟禾补偿生长的影响[J]. 生态学报, 30(2): 279-287.]
MA WM, WANG YF, ZHAO XW, et al., 2019. Altitude differences in reproductive characteristics and resource allocation of Saussurea wellbyi [J]. Bull Bot Res, 39(5):707-715. [马文梅, 王一峰, 赵夏纬, 等, 2019. 羌塘雪兔子繁殖特征及资源分配的海拔差异[J]. 植物研究, 39(5): 707-715.]
MENDEZ M, 1999. Effects of sexual reproduction on growth and vegetative propagation in the perennial geophyte Arum italicum (Araceae) [J]. Plant Biol, 1(1): 115-120.
SCHMID B, BAZZAZ FA, WEINER J, 1995. Size dependency of sexual reproduction and of clonal growth in two perennial plants [J]. Can J Bot, 73(11): 1831-1837.
SUN H, JIANG SY, FENG CQ, et al., 2012. Status of wild resource of medicine plant Lamiophlomis rotata and its problems in sustainable use [J]. Chin J Chin Mat Med, 37(22): 3500-3505. [孙辉, 蒋舜媛, 冯成强, 等, 2012. 独一味 Lamiophlomis rotata野生资源现状与存在的问题[J]. 中国中药杂志, 37(22): 3500-3505.]
THIELE J, JORGENSEN RB, HAUSER TP, 2008. Flowering does not decrease vegetative competitiveness of Lolium perenne [J]. Basic Appl Ecol, 10(4): 340-348.
VAN DRUNEN WE, DORKEN ME, 2012. Trade-offs between clonal and sexual reproduction in Sagittaria latifolia (Alismataceae) scale up to affect the fitness of entire clones [J]. New Phytol, 196(2): 606-616.
WANG HY, WANG ZW, LI LH, et al., 2005. Reproductive tendency of clonal plants in various habitats [J]. Chin J Ecol, 24(6): 670-676. [王洪義, 王正文, 李凌浩, 等, 不同生境中克隆植物的繁殖倾向[J]. 生态学杂志, 24(6): 670-676.]
WANG YF, JIN J, HOU HH, et al., 2015. Changes in flowering resource allocation of Saussurea dzeurensis with elevations [J]. Chin J Plant Ecol, 39(9): 901-908. [王一峰, 靳洁, 侯宏红, 等, 2015. 川西风毛菊花期资源分配随海拔的变化[J]. 植物生态学报, 39(9): 901-908.]
WU ZY, 1980. Vegetation in China [M]. Beijing: Science Press: 624-649. [吴征镒, 1980. 中国植被[M]. 北京: 科学出版社: 624-649.]
XIE TP, ZHANG GF, ZHAO ZG, et al., 2014. Intraspecific competition and light effect on reproduction of Ligularia virgaurea, an invasive native alpine grassland clonal herb [J]. Ecol Evol, 4(6): 817-825.
ZHANG YJ, CHEN Y, GAO H, et al., 2007. Effects of different seed treatment and field mulching on seed germination of Lamiophlomis rotata [J]. J Gansu Agric Univ, 42(3): 60-63. [张亚娟, 陈垣, 高宏, 等, 2007. 种子处理及田间覆盖对独一味种子萌发的影响[J]. 甘肃农业大学学报, 42(3): 60-63. ]
ZHU C, LUO Y, DONG YB, et al., 2018. Study on suitable areas of Lamiophlomis rotata in Sichuan Province based on 3S technology [J]. Chin Trad Herb Drug, 49(6):1405-1412. [祝聪, 罗瑶, 董永波, 等, 2018. 基于3S 技术的四川省独一味生长适宜区研究[J]. 中草药, 49(6): 1405-1412.]
(责任编辑 周翠鸣)
