不结球白菜BrABF1基因的克隆与功能分析
2021-09-10徐园园李竹帛周贺芳王建军侯喜林刘同坤
徐园园 李竹帛 周贺芳 王建军 王 镇 侯喜林 刘同坤
(1南京农业大学作物遗传与种质创新国家重点实验室/农业农村部华东地区园艺作物生物学与种质创制重点实验室/园艺学院,江苏 南京 210095;2淮南市农业科学研究所,安徽 淮南 232001;3 苏州农业职业技术学院,江苏 苏州 215008)
植物开花是由营养生长到生殖生长的标志,是植物生命活动中的重要过程[1-3]。同时,植物的开花时间是农业生产上的一个重要农艺性状,适宜的开花时间能保证农产品的产量和品质,具有重要的经济学意义。但开花时间的调控本身极为复杂,是植物学领域的研究热点之一。前人通过研究十字花科模式植物拟南芥(Arabidopsis thaliana)的开花机制,总结出六大开花调控路径:光周期途径[4]、春化途径[5]、赤霉素途径[6]、自主途径[7]、温敏途径和年龄途径[8-9]。由此可知,高等植物的开花过程不仅受内源因素(植物体内的激素水平、植物衰老等)的影响,同时也受到外界环境因素(光照长短、低温春化等)的影响。
脱落酸(abscisic acid,ABA)属于半萜类物质,作为内源信使通常被认为是响应各种环境胁迫的“应激激素”,在植物对抗非生物胁迫和生物胁迫中起着重要作用[9],如有研究表明喷施ABA 可提高油菜植株的抗寒性、越冬率和产量[10]。除此之外,ABA 贯穿几乎所有植物的整个生长过程,如参与种子的萌发和休眠、抑制幼苗的生长、促进气孔关闭、促进叶片和果实脱落、促进植物的衰老进程等[11-13]。目前也有研究证明ABA 还广泛参与了植物的成花诱导、花芽分化及开花调控等过程[14],如外源ABA 处理可显著诱导荔枝叶片中ABA 的生成,同时提高荔枝的成花率[15]。但ABA 如何调控植物开花,还值得进一步深入探究。
不结球白菜(Brassica rapassp.chinensis)属于十字花科(Cruciferae)芸薹属(Brassica)植物,因其营养丰富、适应性广、生长周期短等特点在我国农业生产中占据着重要的地位[22]。不结球白菜的开花抽薹时间在品种间存在差异,品质、产量也受开花时间影响[23]。实际生产表明,先期抽薹的不结球白菜产量和品质均存在显著下降现象[24],但是其深层调控机理尚不明确。因此研究不结球白菜抽薹开花的调控机制对于提高其产量以及种质创新具有重要的指导意义。
前期研究已初步发现BrABF3 参与调控不结球的开花时间,在此基础上,继续研究其同源基因BrABF1 对不结球开花时间调控的作用。本研究采用同源克隆的方法获得BrABF1 基因序列,利用生物信息学方法对BrABF1 基因进行序列分析,采用亚细胞定位技术研究不结球白菜BrABF1 的空间表达,利用GUS 染色的方法探究其在拟南芥中的表达部位,利用转基因拟南芥研究BrABF1 对开花时间的影响,以期为选育新品种、明确不结球白菜开花的分子机理提供理论基础。
1 材料与方法
1.1 植物材料
采用南京农业大学园艺学院白菜系统生物学实验室提供的不结球白菜品种苏州青为试验材料,用蒸馏水将种子清洗数次后,置于室内洁净的培养皿中催芽2~3 d,之后将种子播种在灭菌的基质中,将其置于相对湿度为85%±5%,光/暗时间为16 h/8 h,光/暗温度为22℃/18℃的人工气候室中培养[25]。
1.2 DNA、RNA 的提取及cDNA 合成
待苏州青植株长至四叶期时,取叶片0.2 g,在液氮中充分研磨,用植物基因组提取试剂盒(江苏康为世纪生物科技有限公司)提取叶片基因组DNA,作为克隆BrABF1 启动子的模板。用RNA Simple Total RNA Kit (TaKaRa,大连)提取样品总RNA,用PrimeScriptTMⅡ1st Strand cDNA Synthesis Kit(TaKaRa,大连)将RNA 反转录为cDNA 作为克隆基因所需要的模板。用Nano-300 微量分光光度计(杭州奥盛仪器有限公司)检测提取的DNA、总RNA 浓度和纯度。
1.3 BrABF1 基因及其启动子的克隆
从白菜数据库中查找BrABF1 的序列并设计引物BrABF1-F/R,引物序列见表1。以苏州青的cDNA 为模板进行PCR 扩增。PCR 反应体系:模板1.0 μL,正、反引物各1.0 μL,PrimeSTAR Max Premix(2×)10.0 μL,ddH2O 7.0 μL。反应程序:98℃预变性3 min;98℃变性10 s,55℃退火15 s,72℃延伸3 min,共35 个循环;72℃终延伸10 min,4℃保存。用10 g·L-1的琼脂糖凝胶电泳检测PCR 产物并回收条带正确的目的片段。将回收产物连接到 pMD18-T 载体(TaKaRa,大连)上,转化至大肠杆菌DH-5α 感受态(北京全式金生物技术有限公司)中,挑取阳性克隆进行PCR 并测序。
克隆BrABF1 基因的启动子时用不结球白菜苏州青品种的DNA 作为模板,所用引物pBrABF1-F/R 利用NCBI 在线软件Primer-BLAST 设计,引物序列见表1。后续试验过程参考上文。
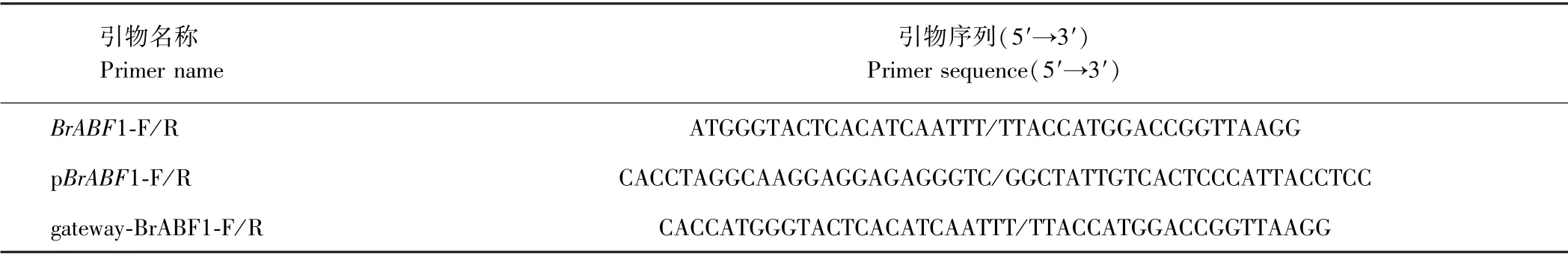
表1 所用的引物Table 1 Primers used
1.4 BrABF1 基因的生物信息学分析
用Bioxm 2.7 软件分析BrABF1 基因序列的开放阅读框(open reading frame,ORF)。为分析BrABF1在进化过程中与其他物种中的亲缘关系,从NCBI 数据库中下载不同物种ABF1 蛋白的氨基酸序列,用DNAMAN 6.0 软件对氨基酸序列进行比对,用MEGA 7.0 软件对氨基酸序列进行系统进化树分析。
丝木棉源于异木棉。三十多年前,异木棉由一颗种子从异国他乡漂洋过海,来到广州。它原产南美洲的阿根廷,树干肥大,有突刺,像刺猬一样,而树高通常在十米到十五米之间,树姿优美,花艳密集。在南方许多城市,特别是在广州,市民直呼它的学名——异木棉,因其繁花挂枝,粉色拂面,格外美丽,也有地方称其为“美人树”或“美丽异木棉”。在日本,异木棉叫白杜,别称丝木棉、桃叶卫矛。在华软学院,丝木棉被誉为校花。
应用在线软件PlantCARE(http:/ /bioinformatics.psb.ugent. be/webtools/plantcare/html/) 分析BrABF1启动子序列中的顺式作用元件。
1.5 Gateway 技术载体的构建
1.5.1BrABF1 基因片段的PCR 扩增BrABF1 载体的构建采用Gateway 技术,所用引物gateway-BrABF1-F/R 序列见表1,用高保真酶进行PCR 扩增,反应程序:98℃预变性30 s;98℃变性30 s,57℃退火30 s,72℃延伸30 s,35 个循环;72℃终延伸10 min,4℃保存。用10 g·L-1琼脂糖凝胶电泳对PCR 产物进行检测,最后用琼脂糖凝胶DNA 回收试剂盒对目的片段进行胶回收。
1.5.2 BP 重组反应 将上一步回收的PCR 产物与入门载体pENTRTM/D-TOPO® 进行重组反应。总体系6.0 μL:PCR 产物2.0 μL、pENTRTM/D-TOPO® 载体1.0 μL、盐溶液1.0 μL、ddH2O 2.0 μL,25℃反应30 min。将反应后的产物转化至DH-5α 感受态细胞中,均匀涂布于含有50 mg·L-1卡那霉素的固体LB 培养基上,置于37℃培养箱中倒置培养过夜。次日早晨挑取阳性单克隆摇菌测序,测序结果正确的即为重组质粒(入门载体)pENTRTM/D-TOPO® -BrABF1。
1.5.3 构建表达载体 反应总体系6.0 μL:表达载体 pEarlyGate101 1.0 μL,pENTRTM/D-TOPO ® -BrABF1 2.0 μL,LR 酶混合物1.0 μL,TE Buffer(pH 值8.0)2.0 μL,25℃反应30 min。反应后的产物转化至DH-5α 感受态细胞中,均匀涂布于含有50 mg·L-1卡那霉素的固体LB 培养基上,置于37℃培养箱中倒置培养过夜,挑取阳性单克隆菌落进行摇菌测序,结果正确说明获得表达载体pEarlyGate101-BrABF1-YFP。
1.5.4 构建GUS 表达载体 构建方法参考1.5.3,总体系6.0 μL:表达载体pFast-G04 1.0 μL,pENTRTM/D-TOPO® -BrABF1 2.0 μL,LR 酶混合物1.0 μL,TE Buffer(pH 值8.0)2.0 μL,25℃反应4 h。转化过程同1.5.3。
1.6 BrABF1 的亚细胞定位
将构建好的融合表达载体pEarlyGate101-BrABF1-YFP 采用液氮冻融法转化至农杆菌GV3101,均匀涂布于含有50 mg·L-1卡那霉素和100 mg·L-1利福平的LB 平板上,将平板倒置于28℃培养箱中培养48 h,挑取阳性单克隆菌落进行摇菌测序,检测结果正确的即为pEarlyGate101-BrABF1-YFP 转化成功的GV3101菌株。
将阳性单克隆菌液在含有卡那霉素和利福平的LB液体培养基中28℃、180 r·min-1培养20 h,接着取1%体积分数的菌液进行扩大培养,当菌液的OD600值达到1.0左右时,收集菌株,4 000 r·min-1离心5 min。用终浓度为10 mmol·L-1MgCl2、10 mmol·L-12-N-吗啉乙磺酸(2-Morpholinoethanesulfonic Acid,MES,pH 值5.7)、150 μmol·L-1乙酰丁香酮重悬菌体,使其OD600值为0.8,室温放置5 h,将含有pEarlyGate101-BrABF1-YFP 的农杆菌菌液用1 mL 的无针头注射器注射于一月龄的烟草叶片背面,其中H2B-RFP 用于细胞核标记。注射后的烟草植株置于人工气候室培养3 d 后,在激光共聚焦显微镜下观察荧光情况并拍照记录。
1.7 转基因拟南芥的获得
将GUS 表达载体pBrABF1:GUS 和过表达载体pEarlyGate101-BrABF1-YFP 采用液氮转化法分别导入农杆菌GV3101(详细转化步骤见1.6)。将含有pBrABF1:GUS 和pEarlyGate101-BrABF1-YFP 的农杆菌分别接种于5 mL 含利福平和卡那霉素的液体LB(Luria-Bertani)培养基中,28℃、180 r·min-1振荡培养18 h 后将菌液转移至200 mL 含有相同抗生素的LB培养基中,28℃、180 r·min-1振荡培养至OD600值为1.0,5 000 r·min-1离心5 min 收集菌株,用渗透缓冲液重悬,使OD600为0.8。用蘸花法将拟南芥的花序浸入重悬液,侵染1 min,每周侵染1 次,共侵染2~3 次。刚侵染完植株所收获的种子记为T0代。之后将消毒后的T0代种子播种于具有潮霉素(Hygromycin B)和特美汀(Termetin)的MS 板上,筛选出pBrABF1:GUS和35S:BrABF1-YFP阳性植株。
1.8 GUS 组织染色
利用潮霉素抗性在MS 固体培养基上筛选pBrABF1:GUS 转基因拟南芥至T3代,用浓度为1 mg·mL-1的X-Gluc(5-溴-4-氯-3-吲哚-β-D-葡萄糖苷酸)染色液对板上15 d 大的拟南芥进行GUS 染色:将植株置于90%的丙酮溶液中预冷10~15 min;染色液清洗1~2 次;将植株置于染色液中,抽真空2~3次,每次10 min;37℃保温12 h;30%酒精脱色30 min;用含有50%酒精、37%甲醛、5%乙酸的脱色液脱色1次;70%酒精清洗1 次。染色结束后将植株置于体式显微镜下观察并拍照记录。
1.9 过表达转基因植株开花时间统计分析
以野生型和BrABF1 过表达转基因拟南芥植株T2代为材料,统计其在短日照条件下的开花时间。将两种基因型的拟南芥种植于相对湿度为85%±5%,光/暗时间为8 h/16 h,光/暗温度为22℃/18℃的人工气候室中。将播种的日期计为0 d,统计其开花时所用的时间,统计开花时的基叶数,并用Excel 软件进行t 检验显著性分析。
2 结果与分析
2.1 BrABF1 基因的克隆
以不结球白菜苏州青的cDNA 为模板克隆出来的BrABF1 条带如图1-A 所示。序列分析表明,不结球白菜BrABF1 基因含有1 个1 104 bp 的ORF,共编码367个氨基酸(图1-B),BrABF1 基因的GeneBank 登录号为MT643821。
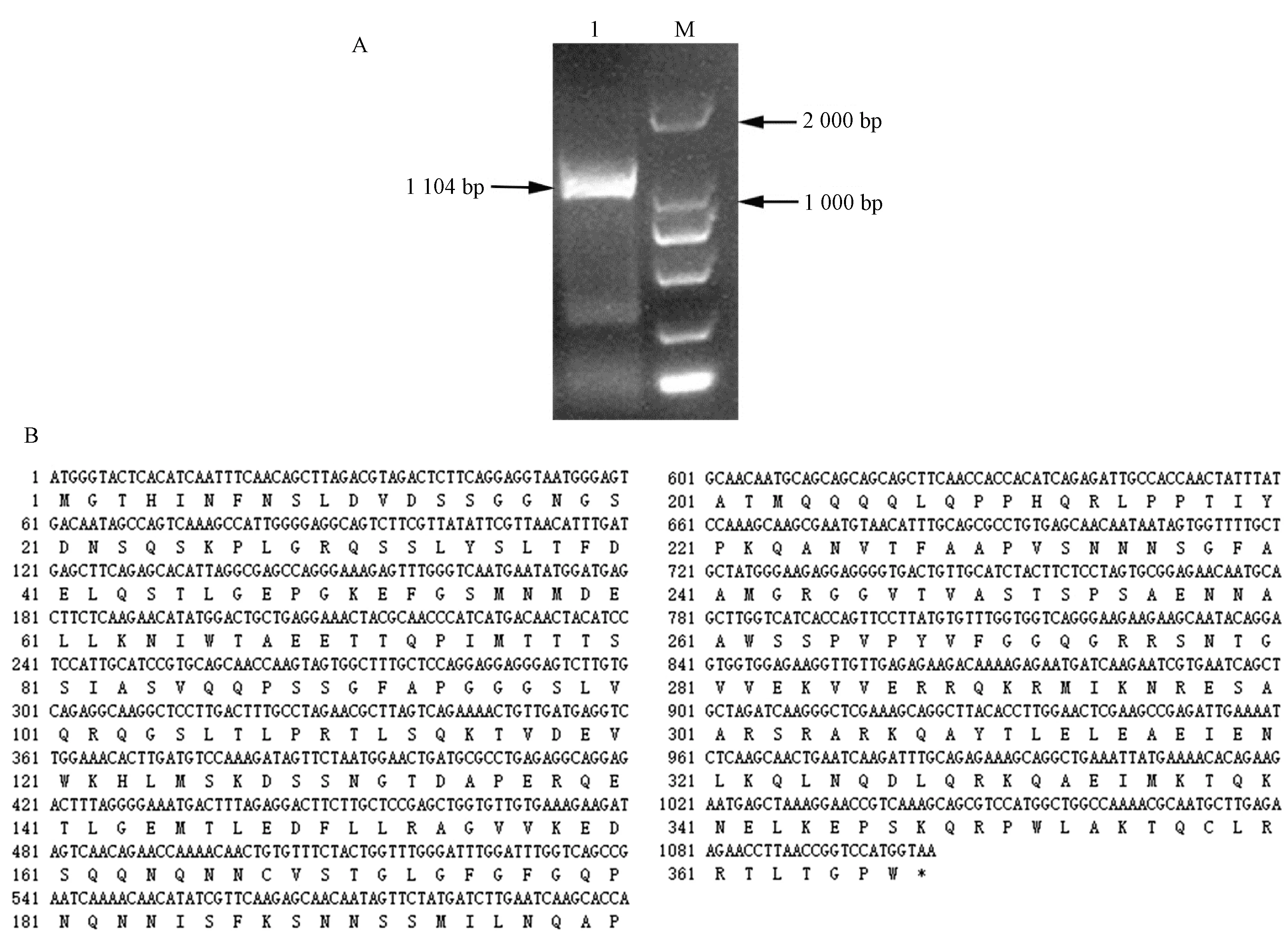
图1 BrABF1 基因的PCR 扩增产物(A)及核苷酸、编码氨基酸序列(B)Fig.1 PCR amplified product of BrABF1(A)and nucleotide sequence,its encoded amino acid sequence (B)
2.2 生物信息学分析
2.2.1BrABF1 的氨基酸序列比对和进化分析 用DNAMAN 6.0 软件对ABF1 氨基酸序列进行比对(图2-A),用MEGA 7.0 软件对氨基酸序列进行系统进化树分析,如图2-B 所示。不结球白菜与甘蓝型油菜(Brassica napus)、高山离子芥(Chorispora bungeana)、萝卜(Raphanus sativus)、拟南芥(Arabidopsis thaliana)、荠菜(Capsella rubella)、山萮菜(Eutrema salsugineum)、亚麻芥(Camelina sativa)、野甘蓝(原变种)(Brassica oleraceavar.oleracea) 的同源性分别为89.30%、65.25%、83.46%、70.47%、72.15%、65.05%、71.22%、89.30%;与不结球白菜、甘蓝型油菜和野甘蓝(原变种)的同源性最高,亲缘性关系最近。
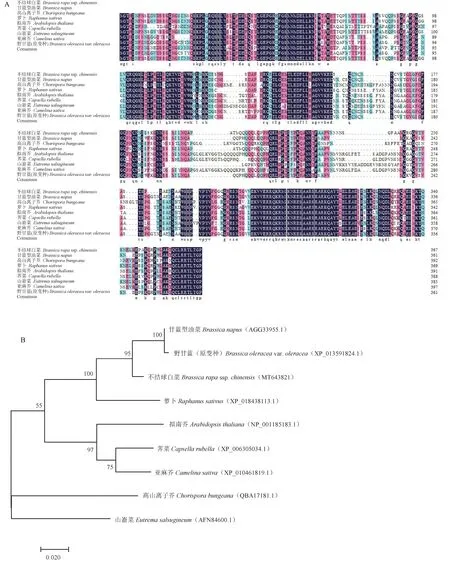
图2 不结球白菜与其他物种ABF1 同源氨基酸序列比对(A)及进化树分析(B)Fig.2 The alignment of amino acid of ABF1 from non-heading Chinese cabbage and other plants(A) and phylogentic tree(B)
2.2.2BrABF1 启动子序列分析 通过用PlantCARE在线软件分析BrABF1 启动子序列(表2),结果发现,BrABF1 启动子序列中除了有大量的TATA-box 和CAAT-box 启动子基本核心元件之外,还有很多光响应元件、参与激素反应的顺式作用元件,以及少量参与低温响应的顺式作用元件、参与防御和应激反应的顺式作用元件等。这些结果表明光照条件、植物激素以及逆境胁迫可能参与BrABF1 基因的表达调控。
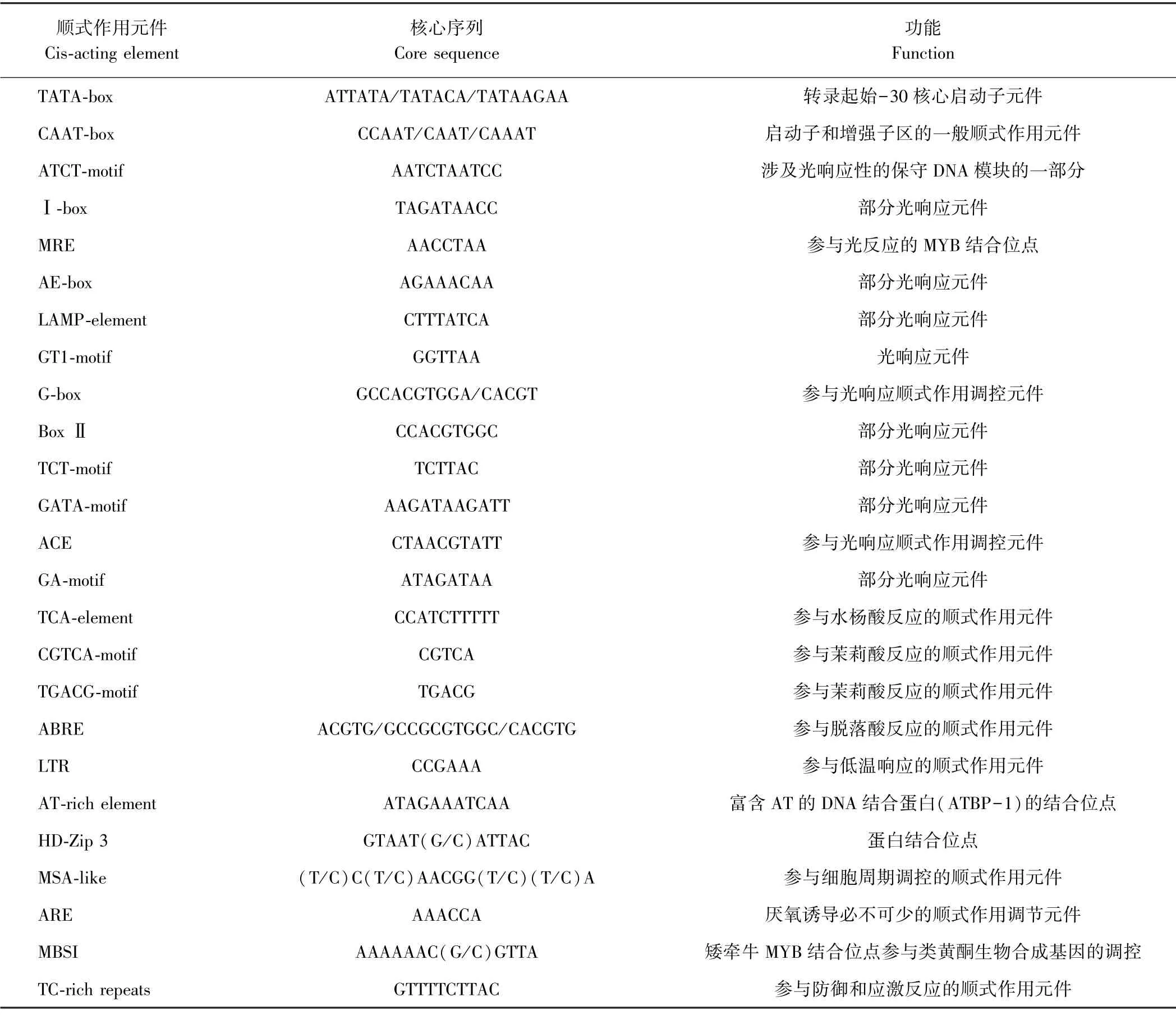
表2 BrABF1 启动子区包含的顺式作用元件Table 2 Cis-acting elements in BrABF1 promoter
2.3 BrABF1 的亚细胞定位分析
利用共聚焦显微镜观察注射后的本氏烟草叶片中的荧光,结果表明(图3):pEarleyGate101-BrABF1-YFP 在黄色荧光蛋白(yellow fluorescent protein,YFP)激发光下可以看到明显的黄色荧光,核标签在红色荧光蛋白(red fluorescent protein,RFP)激发光下显示红色荧光,且黄色荧光和红色荧光重合,表明BrABF1 蛋白定位在细胞核上。说明BrABF1 基因可能具有编码蛋白的功能。
2.4 GUS 组织染色
GUS(β-D-glucuronidase,β-D-葡糖醛酸酶)染色实验中,在体视显微镜下观察到pBrABF1:GUS 转基因拟南芥的叶脉部分几乎均被染色,而野生型拟南芥的叶脉处则没有被染色(图4)。说明能在pBrABF1:GUS 转基因拟南芥的叶脉处检测到GUS 蛋白的表达,即:由BrABF1 驱动的GUS 蛋白主要在叶脉中特异性表达。
2.5 转基因植株开花时间统计分析
通过对野生型和35S:BrABF1-YFP过表达转基因植株在短日照条件下的开花时间统计分析可知,转基因植株开花时的基叶数明显多于野生型植株(图5-A);转基因植株的开花时间明显晚于野生型植株(图5-B,C)。表明在短日照条件下,BrABF1 基因在植物的开花过程中起抑制作用。
3 讨论
植物在适宜的外部环境中,生长到一定阶段的时候,其形态结构和生理结构均会发生改变,其中由营养生长到生殖生长的转变过程最为典型[26-27],这个过程即高等植物的开花过程。有研究表明,不结球白菜抽薹开花性状均受2 对主基因控制[28]。此外,该过程受多种激素调控,其中植物激素起到了至关重要的作用[29]。ABA 是植物的五大内源激素之一,众多研究表明,ABA 对植物开花具有重要影响,如刘扬等[30]认为适量ABA 处理可以诱导青葙花芽形成;喻雄等[31]研究表明ABA 有利于油茶花芽的生理分化和生长,高含量的ABA 则会促进花的凋落。而外源ABA 的施用可能会推迟拟南芥的开花时间[32]。
开花是影响不结球白菜品质和产量的一个重要因素,ABF1 是植物组织中ABA 介导的渗透压信号传导的主要下游转录因子[33],其基因表达水平低于其他ABF基因,但同样在干旱胁迫中起着重要作用[20]。酵母单杂试验表明,ABF1 能调节AtTPPI的表达应对干旱胁迫[34],本研究以不结球白菜苏州青的cDNA 为模板克隆出BrABF1 基因,通过对其启动子序列分析可知,BrABF1 基因的表达可能与光照、植物激素以及胁迫环境等因素有关;亚细胞定位试验结果表明,BrABF1 蛋白定位在细胞核上,说明BrABF1 基因可能具有编码蛋白的功能;GUS 染色结果表明BrABF1 启动子驱动的GUS 蛋白主要在拟南芥的叶脉中特异性表达;通过统计短日照条件下野生型和转基因植株的开花时间推测BrABF1 基因可能抑制植物的开花。有研究表明,包括ABI5 和其他的ABF 转录因子(如ABF1、ABF3 和ABF4)的bZIP 转录因子能直接促进FLC的表达,在ABA 介导的开花时间中起抑制作用[35]。
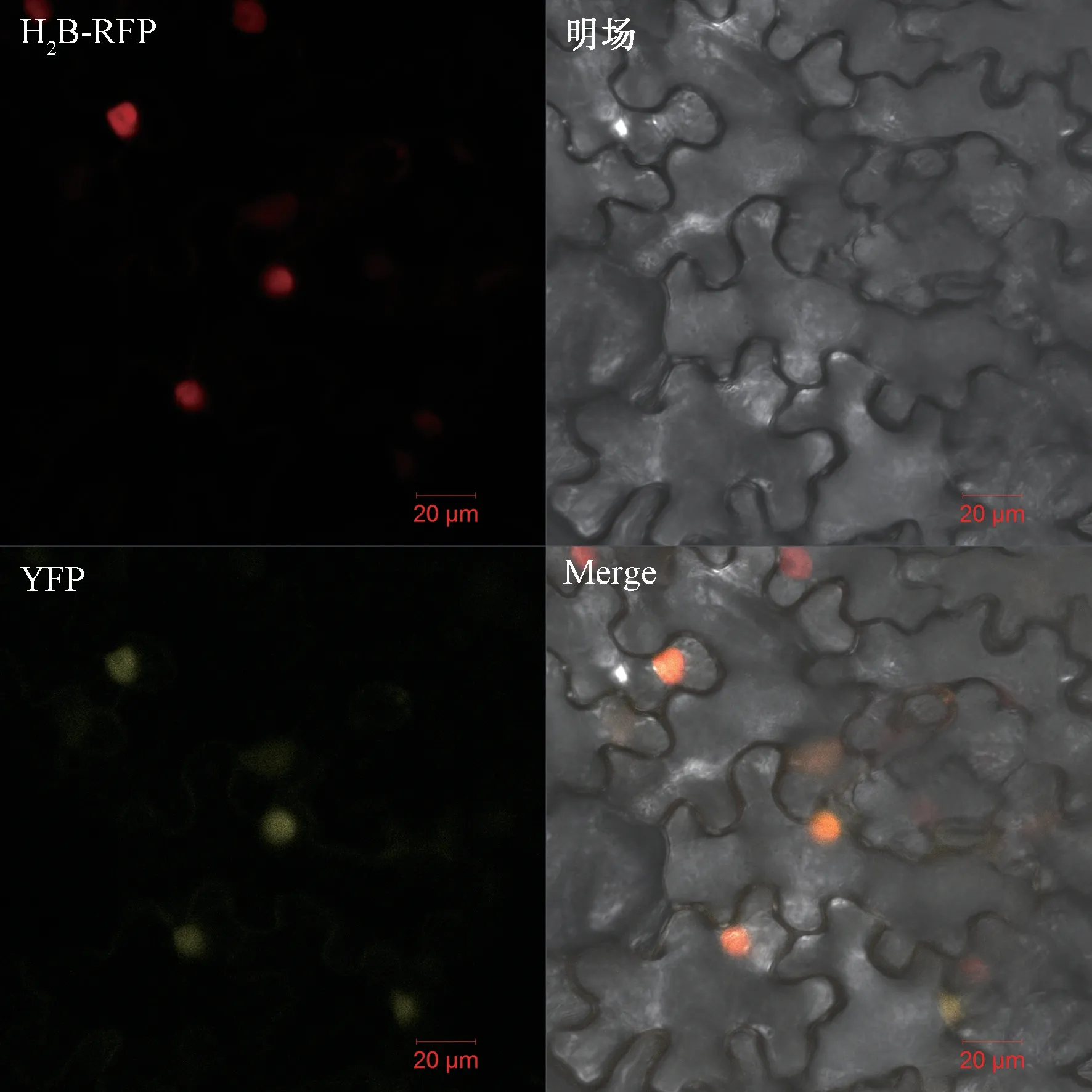
图3 BrABF1 在本氏烟草叶片中的亚细胞定位Fig.3 Subcellular localization of BrABF1 in leaves Nicotiana benthamiana

图4 拟南芥幼苗GUS 染色Fig.4 GUS staining of Arabidopsis seedlings

图5 短日照条件下转基因拟南芥开花时间统计分析(A 和B)及与野生型植株表型对比(C)Fig.5 Statistical analysis of flower time in transgenic Arabidopsis thaliana (A and B) and phenotypic comparison with wide type plants (C) under short-day conditions
基于上述研究结果,推测BrABF1 不仅参与植物ABA 信号途径的抗逆反应,而且能调控植物开花。这对今后进一步深入研究ABA 介导的不结球白菜开花具有很大的借鉴意义,可为提高不结球白菜的商品价值奠定基础。
4 结论
本研究从不结球白菜中克隆出BrABF1 基因,BrABF1 基因ORF 序列长1 104 bp,编码367 个氨基酸,其表达可能与光照、植物激素以及胁迫环境等因素有关。BrABF1 蛋白定位在细胞核上,主要在植物的叶脉中特异性表达。BrABF1 过表达植株在短日照条件下开花时间延迟,推测BrABF1 基因能抑制不结球白菜的开花。
